
Abstract
Tissue-engineered skeletal muscle constructs have attracted significant attention in the field of regenerative medicine. However, the contractile force produced by tissue-engineered skeletal muscle constructs is not comparable with that of natural skeletal muscle tissues. To improve contractile force generation of tissue-engineered skeletal muscle constructs, we investigated the effects of co-culturing C2C12 myoblasts with PC12 neural cells in the fabrication of skeletal muscle tissue constructs. Immunocytochemical analysis revealed differentiation of C2C12 myotubes and outgrowth of PC12 neurites toward the aligned myotubes with improved βIII-tubulin-positive neuronal area, suggesting enhanced neural differentiation of PC12 cells. Furthermore, co-culturing with PC12 cells improved the formation of sarcomeres and contractile activities of C2C12 myotubes, while the addition of nerve growth factor enhanced PC12 neurite outgrowth and further enhanced C2C12 myotube contractile activities. Formation of neuromuscular junctions in the C2C12/PC12 co-culture system was evidenced by the formation of acetylcholine receptor clusters, chemical inhibition of contractile myotubes, and expression of agrin. We then applied this co-culture system to the fabrication of neuron-incorporated skeletal muscle constructs using the magnetic force-based tissue engineering technique. In response to electrical pulses, the co-cultured tissue constructs generated maximum twitch (65.8 μN/mm2) and tetanic (135.1 μN/mm2) forces that were twofold higher than those of C2C12 monocultured constructs (twitch force, 36.3 μN/mm2; tetanic force, 65.9 μN/mm2). These results indicate that the nerve cell-incorporated skeletal muscle tissues constructed in this study could be used in drug testing and biological research for neuromuscular diseases.
Impact Statement
In this study, we fabricated innervated skeletal muscle tissue constructs comprising C2C12 myoblasts and PC12 neural cells using a magnetic force-based tissue engineering technique. We found that the C2C12/PC12 co-culture enhanced neural differentiation of PC12 cells and sarcomere formation of C2C12 myotubes, accompanying with neuromuscular junction formation. The innervated skeletal muscle tissue constructs generated significantly higher contractile forces compared with aneural (C2C12 monoculture) skeletal muscle tissue constructs. These innervated skeletal muscle tissue constructs can be a useful tool for drug testing and biological research for neuromuscular diseases.
Introduction
T
To date, many techniques have been applied for the fabrication of skeletal muscle tissue constructs, including myoblast alignment by topographical constraints, 7 electrical field, 8 mechanical stretching, 9 chemical patterning, 10 and three-dimensional (3D) bioprinting. 11 One promising approach for the fabrication of functional muscle tissue constructs is magnetic force-based tissue engineering (Mag-TE), whereby cells labeled with functionalized magnetic nanoparticles have been used to fabricate tissue constructs by applying a magnetic force.12,13 In our previous study, a Mag-TE technique enabled the formation of a ring-shaped tissue construct possessing high cell density and an oriented multilayered structure. 14 Furthermore, the skeletal muscle constructs fabricated by the Mag-TE technique generated contractile forces in response to electrical stimulation. 15 However, the contractile force of these artificial skeletal muscle tissue constructs was considerably low compared with that of native skeletal muscle tissues.
Heterotypic cellular interactions are vital for biological functions, and co-culturing with different cell types is a promising strategy for establishing cellular communication in vitro. 16 It has been reported that co-culturing of C2C12 cells with fibroblasts can improve myogenic cell differentiation, 17 and that co-culturing of C2C12 cells with endothelial cells can result in the development of prevascularized tissues with the formation of a capillary network. 18 The major hallmark of skeletal muscle tissue maturation and functionality is the force generation capacity. Because the force generation capacity depends on electrical pulse stimulation (EPS) from the central nervous system via the motor neurons, innervation of neurons into the skeletal muscle tissue constructs is a feasible approach. Moreover, neuronal input is intrinsic for optimal muscle development and function, and its absence results in the degeneration of myotubes. 19 Innervation of neurons into a muscle fiber forms a chemical synapse between neuron and muscle cells, called the neuromuscular junction (NMJ). In NMJs, synaptic transmission is achieved through the action of neuronally released acetylcholine (Ach) and acetylcholine receptors (AchR) located in the postsynaptic membrane of muscle fibers. 20 In vertebrates, clustering of AchRs is the main feature of NMJ formation. 21 During the interaction between neuron and muscle cells, the neuron secretes a 200 kDa proteoglycan, agrin, that causes AchR clustering on the surface of cultured myotubes. 22
Recently, Ostrovidov et al. reported that 3D co-culture of C2C12/PC12 cell is effective for the maturation of C2C12 myotubes and upregulation of neuromuscular markers. 23 However, the crosstalk between two different types of cells and the impact on enhancement of contractile force generation has yet to be elucidated. In the present study, we aimed to investigate whether co-culturing of C2C12 cells with PC12 neural cells leads to the formation of NMJs and improves the contractile force generation of skeletal muscle tissues constructed by Mag-TE.
Materials and Methods
Experimental design
The objective of the study was to investigate whether co-cultured muscle–neuron constructs improve contractile force generation. For this purpose, this study included the following experiments: (1) two-dimensional (2D) muscle–neuron co-culture with and without nerve growth factor (NGF) was conducted to investigate the effects of the co-culture on contractile activity of myotubes and formation of NMJ (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea); (2) further, 2D muscle monoculture with and without conditioned medium was conducted to explore whether the effects were caused by paracrine factors secreted from neural cells or attributed by cell–cell interaction between myotubes and neural cells; (3) finally, 3D muscle–neuron co-culture was conducted by fabricating muscle–neuron constructs to investigate whether contractile force generation was enhanced (Supplementary Fig. S2).
Cell culture
Mouse myoblast C2C12 cells (American Type Culture Collection, Manassas, VA) were grown in low-glucose Dulbecco's modified Eagle's medium (LG-DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin G potassium, and 0.1 mg/mL streptomycin sulfate. To induce myogenic differentiation, the medium was changed to LG-DMEM supplemented with 2% calf serum (CS) for 2D cell culture or 0.4% Ultroser G (Pall, East Hills, NY) for 3D tissue culture, with antibiotics. Cells were cultured at 37°C in a 5% (v/v) CO2 incubator.
Rat pheochromocytoma PC12 neural cells (Riken Bioresource Center, Ibaraki, Japan) were grown in DMEM (high glucose) supplemented with 10% FBS, 10% horse serum (HS), and antibiotics. Cultures were maintained for 3 days at 37°C in a 5% (v/v) CO2 incubator until confluence and were then harvested for co-culture. For neural differentiation, the medium was changed to high-glucose DMEM supplemented with 1 ng/mL NGF (R&D Systems, Minneapolis, MN), N2 supplement (Invitrogen, Carlsbad, CA), and antibiotics.
2D muscle–neuron co-culture with and without NGF
For 2D muscle–neuron co-culture, C2C12 cells (2 × 104 cells) were seeded into wells of a 24-well tissue culture plate (Thermo Fisher Scientific, Darmstadt, Germany) in LG-DMEM with 10% FBS. On day 2 of culturing, PC12 cells (2 × 104 cells) were seeded onto the C2C12 cells (day 0). The co-culture condition was maintained for 7 days by replacing the differentiation medium (LG-DMEM with 2% CS with or without the addition of 1 ng/mL NGF and N2 supplement) at 24-h intervals (Supplementary Fig. S1).
Immunofluorescent staining was performed to show PC12 neurites and C2C12 myotubes. Samples of 2D monocultures and co-cultures on day 7 were stained with anti-MHC (sc20641; Santa Cruz Biotechnology, Santa Cruz, CA) and/or mouse anti-β III tubulin (TuJ) antibodies (R&D Systems). For determining the TuJ-positive area, co-cultured samples from days 1, 4, and 7 were stained with anti-TuJ antibody (R&D Systems), and the TuJ-positive area in each field was measured using BZ-Analyzer software and a BZ-9000 microscope (Keyence, Tokyo, Japan). Micrographs of five fields of view (275 × 363 μm) in three separate wells were randomly captured, and myogenic differentiation rate and myotube width were determined using previously described methods. 23 Briefly, cell samples from day 7 were double-stained with anti-α-actinin antibody (EA-53; Sigma-Aldrich, St. Louis, MO) and 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA). Micrographs were then randomly captured and α-actinin-positive myotube widths and numbers of DAPI-stained nuclei were measured using BZ-Analyzer software (Keyence). The differentiation rate was calculated using the following equation: differentiation rate = (number of DAPI-stained nuclei in α-actinin-positive myotubes in a field of view/number of DAPI-stained nuclei in a field of view). To determine myotube width of samples, the five largest myotubes within the five fields of view for each of the three separate wells per sample were measured using BZ-Analyzer software (Keyence).
Assembly of sarcomeres is an important prerequisite for contractile maturity of myotubes. To investigate the striation patterns of myotubes, C2C12 cells (2.7 × 103 cells) and/or PC12 cells (2.7 × 103 cells) were cultured in 35-mm glass-bottom dishes (Code 3970-035; Asahi Techno Glass, Tokyo, Japan). On day 7, the differentiated myotubes of both C2C12 monoculture and C2C12/PC12 co-culture samples in the 35-mm glass-bottom dishes were placed in a chamber for EPS (C-dish; IonOptix, Milton, MA). A previous study reported sarcomere assembly by EPS in C2C12 cells. 24 Also, the sarcomere formation in myotubes decreased rapidly after termination of continuous EPS with changes in atrophy-related gene expression. 25 In the present study, electrical pulses were applied for 30 min at 0.3 V/mm for 4 ms at 2 Hz. Then, immunostaining with anti-α-actinin antibody (Sigma-Aldrich) was performed. Microscopic images of five fields of view in each of three separate dishes were captured under a BZ-9000 fluorescence microscope (Keyence) and processed using BZ-Analyzer software (Keyence). Finally, numbers of myotubes consisting of sarcomeric α-actinin were counted.
Contractile activity of myotubes of both monoculture and co-culture samples in 35-mm cell culture dishes (Greiner Bio-One, Frickenhausen, Germany) was measured before and after incubation with 12.5 μM of neuromuscular blocking agent, (+)-tubocurarine chloride pentahydrate (curare; Tokyo Chemical Industries, Tokyo, Japan) to determine the functional synaptic contacts between neurons and myotubes. On day 7, the dishes were placed in a chamber for electrical stimulation (C-dish). Electrical pulses were applied at 0.3 V/mm for 4 ms at 2 Hz, and myotube movement was recorded at a rate of 15 frames per second for 20 s at three positions on the bottom surface in each of three separate dishes using a BZ-9000 microscope (Keyence). The dishes were then incubated with curare for 15 min at 37°C. The electric pulses were applied again and myotube movement was recorded using a BZ-9000 microscope (Keyence). Because the difference in pixel intensity between the first (displacement = 0) and successive images is mainly due to changes in the movement of objects, the subtracted images indicate moving parts. The moving parts were analyzed by the motion analyzer software (Keyence) and displacement was measured. To estimate the range of displacement, five myotubes displaying the highest contractile activity in each of three fields of view in three separate dishes were measured using the motion analyzer software (Keyence).26,27
For qualitative assessment of the formation of AchR clusters, day 7 samples of 2D monocultures and co-cultures were incubated with Alexa Fluor 488-conjugated α-bungarotoxin (α-BTx; Thermo Fisher Scientific) dissolved in differentiation medium with or without NGF (1:1000) for 1 h at 37°C, before fixation. The samples were then stained with mouse anti-TuJ antibody (R&D Systems). Finally, AchR clusters were observed under a FV10i confocal laser-scanning microscope (Olympus, Tokyo, Japan). For quantitative analysis, after incubation with Alexa Flour 488-conjugated α-BTx (Thermo Fisher Scientific), samples were stained with anti-α-actinin antibody (EA-53; Sigma-Aldrich). Then, the α-BTx-positive area (μm 2 ) within the α-actinin-positive myotubes in each field of view was measured using BZ-Analyzer software (Keyence). A standard cluster was classified as a α-BTx-positive area above 3 μm 2 . Then, the total number of standard clusters in each field was counted. Micrographs of five fields of view in each of three separate wells were randomly captured.
2D muscle monoculture with and without conditioned medium
PC12 culture medium was harvested every 24 h and filtered. C2C12 cells were cultured in culture medium or conditioned medium containing a 1:1 ratio of PC12 culture medium to fresh culture medium. Cells were cultured to day 7 with medium replacement at 24 h intervals. On day 7, myotube contractile activity was determined. 26
Agrin expression level was determined using an enzyme-linked immunosorbent assay (ELISA) kit (Ray Biotech, Norcross, GA). On day 7, cells were washed with phosphate buffered saline and treated with radioimmunoprecipitation assay (RIPA) buffer (Thermo Fisher Scientific). Cells were then collected from the dish by gently scraping with a cell scraper before being transferred to microtubes. The collected co-cultured cell samples were centrifuged at 600 g for 10 min at 4°C, and agrin-ELISA was performed in accordance with the manufacturer's protocol. The limit concentration level of agrin detection using the agrin-ELISA kit was 0.06 ng/mL.
3D muscle–neuron co-culture
Magnetite cationic liposomes (MCLs) were prepared from colloidal magnetite and a lipid mixture consisting of N-(trimethylammonioacetyl)-didodecyl-
For magnetic labeling, C2C12 cells (3 × 106 cells) were seeded in 100-mm tissue culture dishes (Thermo Scientific) containing 10 mL of culture medium in the presence of MCLs (net magnetite concentration, 100 pg/cell), and incubated for 6 h to allow for MCL uptake by the cells. 26 A collagen solution was prepared by mixing type I collagen solution (Nitta Gelatin, Osaka, Japan), DMEM, and neutralization buffer (0.05 M NaOH) at a volume ratio of 8:1:1. A PC12 cell suspension (1 × 106 cells) was mixed with an extracellular matrix (ECM) precursor solution composed of collagen solution (70 μL; final concentration, 0.5 mg/mL), FBS (15 μL), and matrigel basement matrix (15 μL; BD Biosciences). The PC12 cells in the ECM solution (final concentration, 100 μL) were mixed with the MCL-labeled C2C12 cell suspension (1 × 106 cells in 50 μL). The C2C12/PC12 mixture was then added to wells of a 24-well ultra-low attachment plate (150 μL/well; Corning, New York, NY) with a polycarbonate cylinder (diameter, 12 mm; height, 5 mm) fixed at the center of each well, and a neodymium magnet (diameter, 30 mm; magnetic induction, 0.4 T) was placed under the wells. Thereafter, growth medium was added to each well. One day after cell seeding, the cell layer shrank around the cylinder, resulting in the formation of a ring-shaped cellular construct. The cellular ring was removed from the cylinder and hooked around two stainless steel pins (Shiga, Tokyo, Japan) that were positioned 10 mm apart from one another. For myogenic differentiation of tissue constructs, the cellular rings were cultured in differentiation medium consisting of LG-DMEM supplemented with 0.4% Ultroser G (Pall) with or without N2 supplement (Invitrogen) and 1 ng/mL NGF (R&D Systems) in wells of six-well culture plate for 7 days (Supplementary Fig. S2).
Contractile force generation of skeletal muscle tissue constructs was measured as described previously. 15 Briefly, carbon electrodes were placed 10 mm apart at the opposite sides of a tissue culture plate. A muscle bundle was hooked around two stainless steel minute pins (Shiga). One pin was attached to a force transducer (AE-801; SonorOne, Sausalito, CA), and the other was fixed to a silicon rubber sheet placed at the bottom of the culture plate. Electrical pulses were computer-controlled with specially designed LabView software (National Instruments, Austin, TX). For measuring twitch contractions, the tissue sample was stimulated with an electrical pulse of 20 V with a width of 10 ms. For measuring tetanic contractions, the tissue sample was stimulated with electrical pulses with the following properties: voltage, 20 V; width, 10 ms; frequency, 50 Hz; and duration, 2 s. Specific force generation was evaluated using normalized muscle contractility units: force/cross-sectional area of the constructs.
Statistical analysis
Statistical comparisons were evaluated by the Mann–Whitney rank-sum test, and values of p < 0.05 were considered significantly different.
Results
Effects of C2C12 cells on neuronal differentiation of PC12 cells in 2D co-culture
Immunocytochemical analysis of C2C12/PC12 co-cultured cells using anti-MHC (myogenic differentiation marker) and anti-TuJ (neuronal differentiation marker) antibodies revealed that both C2C12 and PC12 cells differentiated into myotubes and neuronal cells, respectively, in co-culture conditions (Supplementary Fig. S3). Furthermore, addition of NGF resulted in extensive neuronal protrusions along the myotubes. To investigate the effects of co-culture on the differentiation of PC12 cells, TuJ-positive neuronal area at different co-culture time intervals was evaluated (Fig. 1). PC12 cells started to differentiate on day 1 of co-culturing, and the TuJ-positive neuronal area increased over the 7-day culture period (Fig. 1A, C). NGF increased the TuJ-positive area of monocultured PC12 cells (Fig. 1B, D). Furthermore, co-cultured C2C12 cells increased the TuJ-positive area of PC12 cells, and the addition of NGF to the co-culture further increased this (Fig. 1B, D).
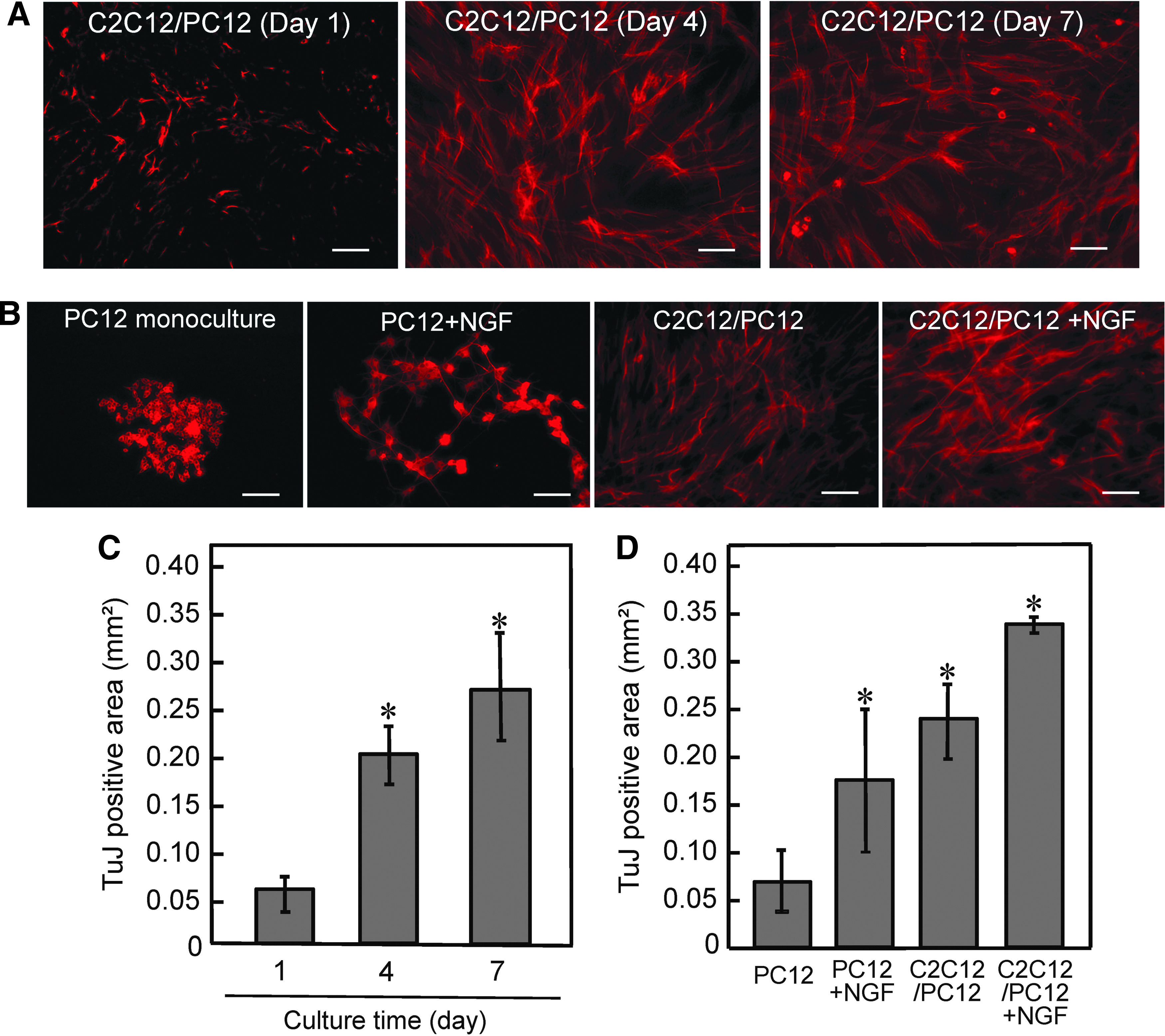
Effects of co-culture on PC12 differentiation.
Effects of PC12 cells on myogenic differentiation of C2C12 cells in 2D co-culture
To investigate the impact of co-culture on myogenic differentiation of C2C12 cells, myotubes on day 7 were immunostained with anti-α-actinin antibody (myogenic differentiation marker) (Fig. 2A), and differentiation rate and myotube width were determined as an indicator of hypertrophy. Both differentiation rate and myogenic width did not differ between co-culture and monoculture conditions (Fig. 2B, C). In contrast, larger numbers of sarcomeric α-actinin-positive striated myotubes were observed in co-cultured myotubes, with NGF addition further increasing the number of myotubes with striation (Fig. 3). To explore the effects of co-culture on myotube function, 2D contractile activity was analyzed by examining myotube movement in response to electrical pulses. Myotubes on day 7 displayed contractile responses to EPS of 0.3 V/mm for 4 ms at 2 Hz. As shown in Figure 4A, C, E, and G, contractile activity of co-cultured myotubes was higher than that of monocultured myotubes, and NGF addition slightly, but not significantly, enhanced myotube displacement.

Effects of co-culture on C2C12 differentiation.
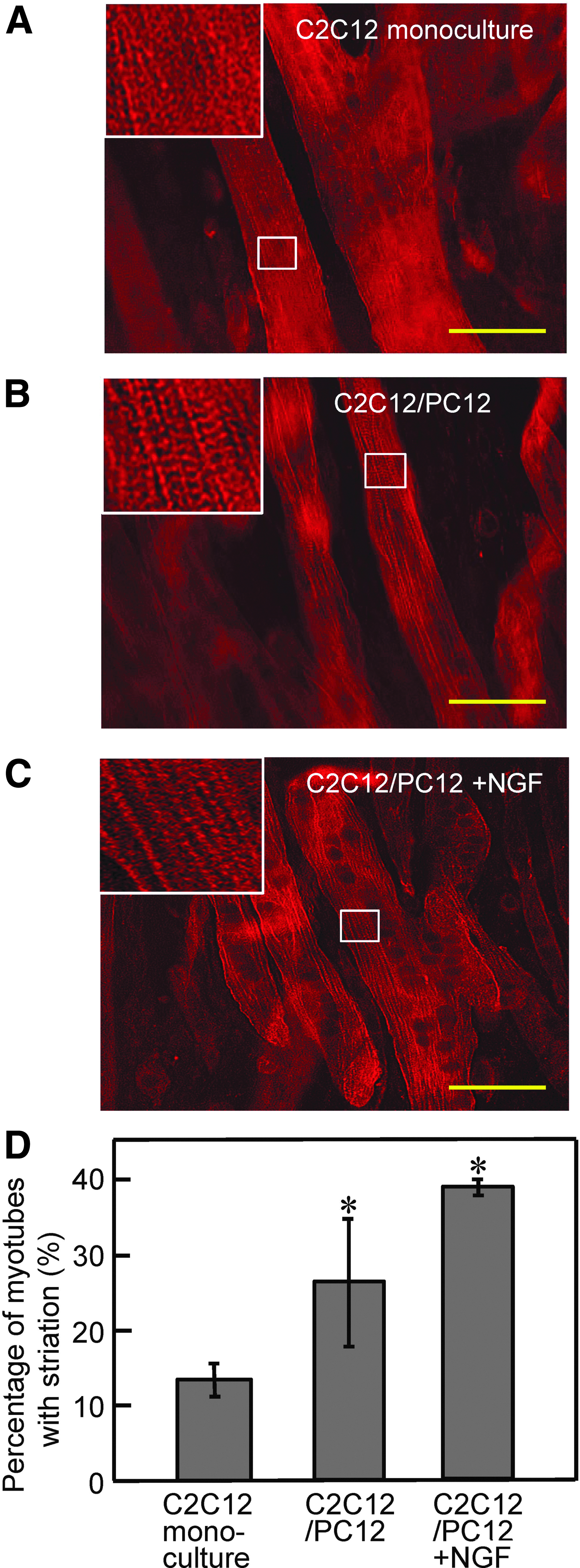
Effect of co-culture on striation formation in myotubes.
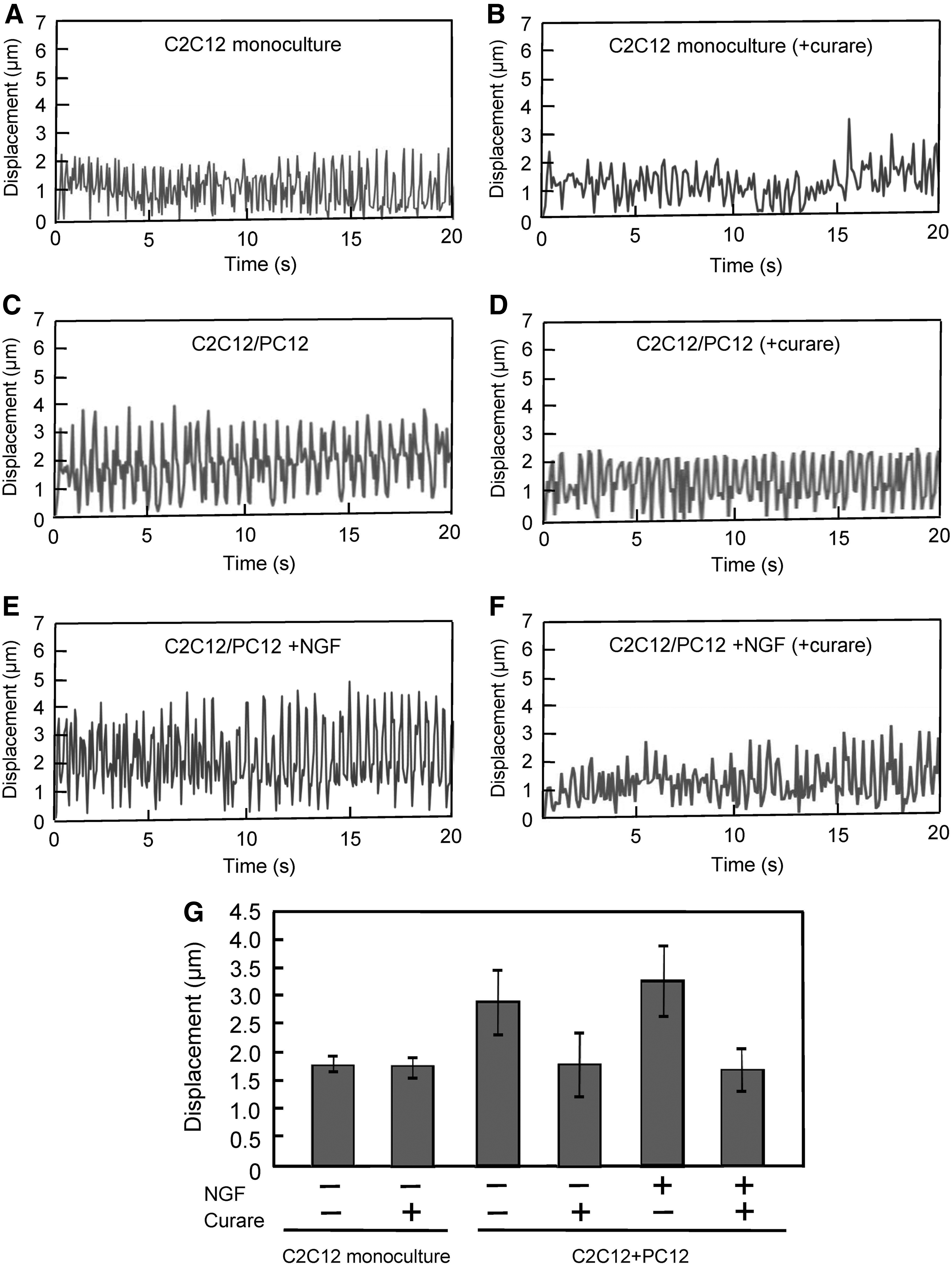
Effects of co-culture on the contractile activity of myotubes.
Formation of NMJs
Curare attenuated the contractile activities of co-cultured myotubes (Fig. 4B, D, F, G), indicating that the enhanced activity of myotubes was attributed to the functional synaptic interaction based on NMJs. AchR clustering between neurons and myotubes is another important feature to determine the formation of NMJs. Immunocytochemical analysis using α-BTx neurotoxin, which is known to bind to the AchR found at the NMJ, revealed that AchR clusters were found with α-actinin-positive myotubes (Fig. 5A) and with TuJ-positive neurites (Supplementary Fig. S4) in C2C12/PC12 co-cultured cells. As shown in Figure 5B, co-cultured cells significantly increased the number of AchR clusters with a >3 μm 2 size compared with monocultured cells.

Effects of co-culture on the formation of AchR clusters on myotubes.
When considering the mechanism of action, PC12-derived neurons may enhance the contractile activity of myotubes by secreting paracrine factors. To elucidate this, C2C12 cells were cultured in conditioned medium and the contractile activities of the myotubes were evaluated under EPS (Fig. 6). No significant differences in both contractile activity profiles (Fig. 6A) and myotube displacement (Fig. 6B) were observed for the cells cultured in conditioned medium, suggesting that the enhanced contractile activity of myotubes in co-culture was attributable to cell–cell attachment between C2C12 and PC12 cells. To further explore the mechanism of how PC12 cells enhanced myotube activity via NMJs, the expression of agrin protein—known to be secreted by neurons and involved in the aggregation of AchR 22 —was investigated. A substantial level of agrin was detected in C2C12/PC12 co-cultured cells and also in monocultured PC12 cells (Fig. 6C), while no agrin was detected in the conditioned medium, suggesting that the level of agrin released from PC12 cells is extremely low and the close contact between C2C12 and PC12 cells is important for mutual interaction. Taken together, these results, that is, attenuation of contractile activity by curare (Fig. 4), formation of AchR cluster (Fig. 5), and agrin expression (Fig. 6C), indicated the formation of NMJs in C2C12/PC12 co-culture.
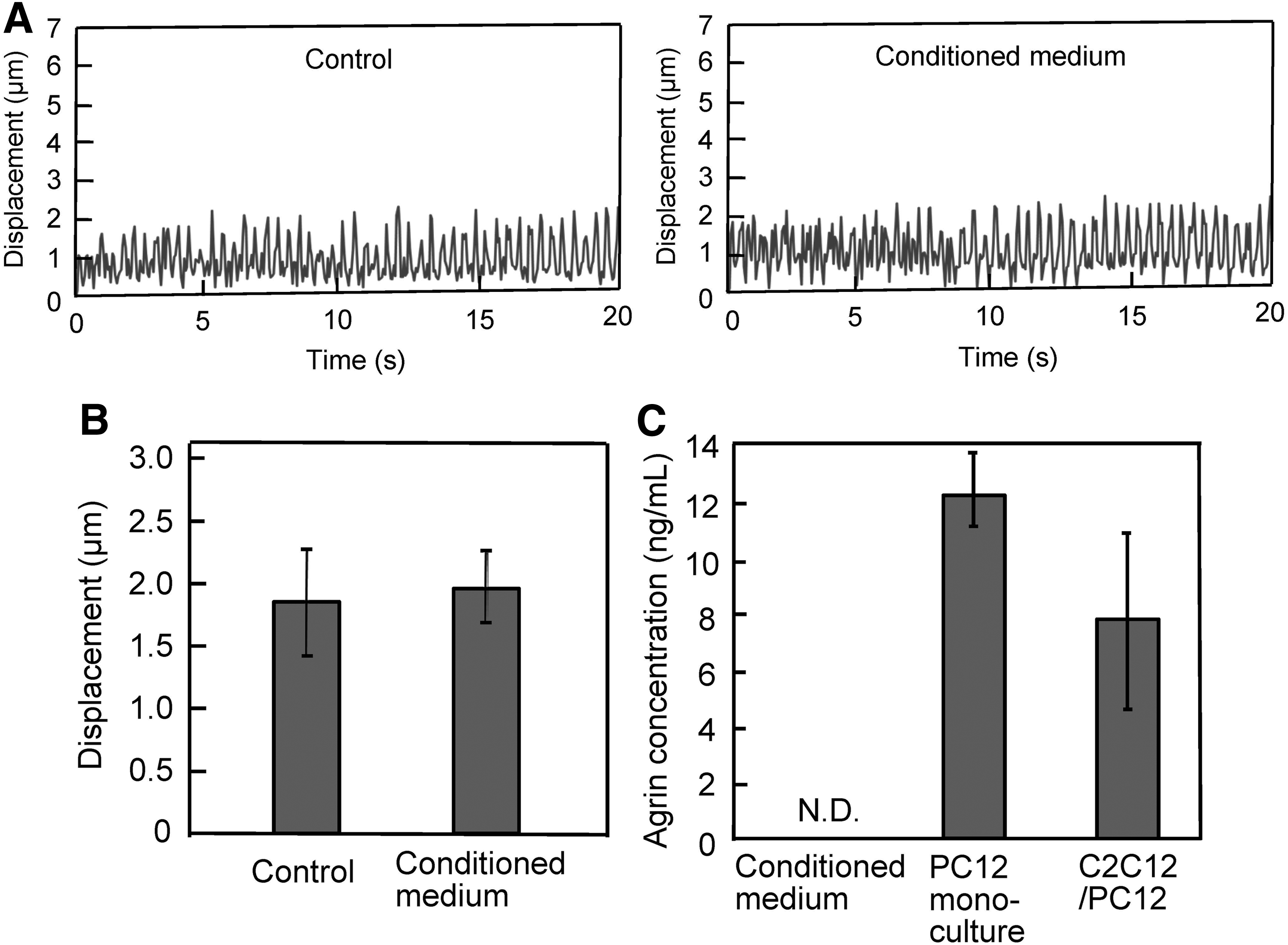
Effects of PC12 cell condition medium on contractile activity. C2C12 cells were cultured with PC12 cell condition medium. EPS (voltage, 0.3 V/mm; width, 4 ms; frequency, 2 Hz) was applied, and the profiles of myotube displacement
Contractile force generation by artificial muscle–neuron constructs
The co-culture system was applied to the fabrication of artificial muscle–neuron constructs using a Mag-TE technique. MCL-labeled C2C12 cells mixed with PC12 cells were uniformly accumulated at the bottom of the well by applying a magnetic force, and a sheet-like structure contracted to form a ring-like tissue construct surrounding the cylinder. The tissue ring coated with ECMs formed fascicle-like tissues on day 7 in the differentiation medium. The average diameter of the tissue-engineered skeletal muscle constructs was ∼500 μm. Histological examination of the tissue constructs showed that C2C12 cells differentiated into multinucleated myotubes and the PC12 cells existed alongside the myotubes (Supplementary Fig. S5). To evaluate contractile force generation, both innervated (C2C12/PC12 co-cultured tissue) and aneural (C2C12 cell tissue) constructs were stimulated with electrical pulses. The force generation profiles in response to electrical pulses are shown in Figure 7A and B. Twitch contractions were observed after low-frequency stimulation (Fig. 7A), while tetanus was observed by repeated electrical stimulation at higher frequency (Fig. 7B). The innervated muscle tissue constructs generated higher physical forces by twitch (Fig. 7C) and tetanus contractions (Fig. 7D) compared with those of aneural muscle bundles, and NGF addition further enhanced force generation. These results clearly indicate that co-culture with PC12 neuronal cells had a positive effect on the contractile force generation of C2C12 skeletal muscle constructs.

Contractile properties of C2C12/PC12 co-cultured tissue constructs.
Discussion
In vitro 3D models of muscle–neuron constructs with NMJs between co-cultured myotubes and neurons would be a considerable benefit for biological study, drug testing, and regenerative medicine. Muscle–neuron co-culture systems using primary neuron and muscle cells have been reported.29,30 Mars et al. utilized an in vitro model in which human myotubes derived from primary muscle satellite cells were innervated by neurons extending from spinal cord explants of fetal rat, and proposed that the model was suitable for functional studies. 31 Larkin et al. engineered nerve–muscle constructs by using rat muscle cells and fetal nerve explants, and evaluated the functional characteristics, including enhanced contractile force generation induced by neural stimulation. 32 Although primary cells, including autologous and patient-derived cells, are considered to be a first choice for regenerative medicine, muscle–neuron co-culture systems based on primary cells have drawbacks with biological studies and for drug testing, including batch-to-batch variation of cell type ratio. Moreover, primary co-culture systems often require a cocktail of multiple myogenic and neurotrophic factors that have not been thoroughly studied. 33 In the present study, we established a C2C12/PC12 co-culture system, analyzed its efficacy on the morphology and contractility of myotubes in 2D culture, determined the formation of NMJs, and finally applied this system to the fabrication of 3D muscle–neuron constructs to improve contractile force generation. Cell lines, including C2C12 and PC12 cells, have the capability to proliferate indefinitely. In addition, the use of cell lines overcomes problems of cell type heterogeneity associated with primary cell cultures. Thus, we used C2C12 and PC12 cell lines to allow for the study of experimental parameters under a controlled and reproducible environment, which is more amenable to analysis.
In the present study, we showed that both C2C12 and PC12 cells underwent clear differentiation in the co-culture condition. It has been previously reported that neurotrophins, which are involved in neurite outgrowth 34 and include NGF, brain-derived neurotrophic factor (BDNF), and neurotrophin-3 (NT-3), are secreted from C2C12 cells. 35 Thus, the neurotrophins secreted from C2C12 cells may induce PC12 neural differentiation within the co-culture system, and addition of external NGF further augmented this function (Fig. 1). Interestingly, the C2C12/PC12 co-culture system had any impact on neither myogenic differentiation rate nor myotube hypertrophy. Similarly, Martin et al. reported that the presence of motor neurons had no significant effects on myogenic cell fusion and myotube hypertrophy. 36 In contrast, Ostrovidov et al. reported that C2C12/PC12 co-culture improved the myogenic differentiation rate. 23 The differentiation rate in the present study (around 40%; Fig. 2) was much higher than that reported by Ostrovidov et al. (10%–20%). In addition, Martin et al. used primary muscle-derived cells and constructed highly matured 3D skeletal muscle tissues with striated myotubes. 36 The level of muscle striation is recognized as a surrogate marker of muscle maturity. In the present study, the number of myotubes displaying striation was increased by co-culture (Fig. 3D). We therefore speculate that the degree of myogenic differentiation (differentiation and maturation level) could explain differences between studies.
We previously demonstrated that morphological analysis using myogenic differentiation markers, including MHC and α-actinin, is insufficient for evaluating the function of skeletal muscle tissues. This is because the level of myogenic differentiation rate is not always consistent with that of contractile force generation of 3D tissue-engineered skeletal muscle constructs. 26 Contractile force generation capability should be the most important feature of tissue-engineered skeletal muscle constructs. This is because it reflects not only the level of differentiation—including the expression of muscle regulatory factors, but also the niche, including the ECM, as well as the architecture, including sarcomere formation. In the present study, the number of myotubes with striation of sarcomeric α-actinin was increased in co-culture conditions (Fig. 3D), along with enhanced 2D contractile activity (Fig. 4), and these parameters were highly correlated with contractile force generation of 3D tissue-engineered skeletal muscle constructs (Fig. 7). These results suggest that as well as evaluating contractile force generation of 3D tissue-engineered skeletal muscle constructs, the evaluation of sarcomere formation and/or contractile activity of myotubes in 2D culture is applicable as an indicator for drug testing of neuromuscular diseases.
The goal of this study was to fabricate innervated 3D skeletal muscle tissue constructs with contractile force generation capability using the C2C12/PC12 co-culture system. The Mag-TE technique has been demonstrated as a powerful tool for fabricating contractile force-producing skeletal muscle tissue constructs.15,26,27 In this study, co-cultured muscle tissues were successfully fabricated using the Mag-TE technique, and the C2C12/PC12 co-cultured tissues showed significant improvement of physical force generation (Fig. 7). EPS from motor neurons is the most important cue for skeletal muscle development and maturation 37 ; therefore we hypothesized that co-culturing with PC12 neural cells can mimic the in vivo niche to engineer in vitro functional C2C12 skeletal muscle tissues. Neuronal innervation leads to phenotypical changes of both AchR and sodium channel subtype, as well as MHC expression, 38 and it is believed that these types of changes in molecular expression result in the enhancement of contractile activity. In the present study, we showed NMJ formation as evidenced by AchR clustering (Fig. 5) with agrin expression (Fig. 6C), and inhibition of myotube contraction by curare (Fig. 4). A possible mechanism for the enhanced contractile activity in this study may be the release of Ach from PC12 cells, because Ach can activate myotubes at the NMJ. To elucidate this mechanism, we added the AchR antagonist curare to both monocultured and co-cultured cells. Curare significantly attenuated the contractile activity of co-cultured myotubes, but had no effect on the monocultured myotubes. These results suggest that the enhanced contractile activity of co-cultured myotubes was caused by Ach-mediated neural induction. The AchR clustering is triggered by agrin. 22 However, no detectable agrin level in the medium was produced by PC12 cells (Fig. 6C), and the conditioned medium did not increase the contractile activity of C2C12 myotubes (Fig. 6A, B). Similarly in our previous reports, 39 although mouse embryonic fibroblasts (MEFs) as a feeder produce many growth factors that are important for self-renewal of embryonic stem (ES) cells, conditioned medium derived from MEF culture cannot maintain pluripotency of ES cells, indicating that the amount of growth factors secreted from MEF is not enough for inducing the performance. In contrast, magnetic accumulation of magnetically labeled feeder cells onto ES cells improved undifferentiated growth of ES cells, 39 suggesting that physical contact (specifically tight and close) between target cells and feeder cells is essential for efficient provision of growth factor signals. In the case of agrin, because motor neurons secrete agrin into the synapse, close interaction between neuron and muscle is important. These results suggest that co-culture system is effective to form cell–cell interaction, and C2C12/PC12 system successfully formed NMJ, resulting in augmentation of contractile force generation.
In the present study, we developed innervated 3D skeletal muscle constructs using C2C12/PC12 cell lines. Although force generation capability of C2C12 cell tissues was enhanced by co-culturing with PC12 neural cells, the maximum contractile force (135.1 μN/mm 2 ) was still limited (∼0.1% of that of mammalian adult skeletal muscle tissues40,41). Currently, effects of the co-culture of primary myoblasts with primary motor neuron are being investigated.
Conclusion
We successfully fabricated innervated skeletal muscle tissue constructs using a C2C12/PC12 co-culture system and Mag-TE. These innervated skeletal muscle tissue constructs generated significantly higher contractile forces compared with aneural (monoculture) skeletal muscle tissue constructs. These innervated skeletal muscle tissue constructs may be a useful tool for drug testing and biological research for neuromuscular diseases.
Footnotes
Acknowledgments
This work was financially supported in part by Grants-in-Aid for Scientific Research (Nos. 23686121 and 17H03469) of the Japan Society for the Promotion of Science (JSPS). One of the authors (M.A.) is a scholar of the Ministry of Education, Culture, Sports, Science, and Technology (MEXT, Japan). We thank Elizabeth Finnie, PhD, from Edanz Group (![]() ) for editing a draft of this article.
) for editing a draft of this article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.