
Abstract
One of the major problems in bone regeneration is that seeded cells do not effectively infiltrate into the central scaffolds. In this study, we generated chitosan/gelatin scaffolds (C group), and investigated whether exogenous addition of stromal cell-derived factor 1 (SDF-1) in the scaffolds (S group) and additional hypoxic preconditioning (S/P group) could enhance bone regeneration by promoting cell migration of rat bone marrow stromal cells (BMSCs). SDF-1 effectively promoted cell migration in vitro and this effect was further enhanced by hypoxic preconditioning. The S/P group had a better cell viability than the other two groups (all p < 0.05). Moreover, hypoxia-inducible factor 1 α and CXC chemokine receptor 4 (CXCR4) in BMSCs were significantly higher in the S/P group. For in vivo studies, bone regeneration ability was evaluated at 7, 14, and 21 days by microcomputed tomography. The S/P group showed a statistically greater amount of bone formation compared to the C group and S group at each time point (all p < 0.05). While the C group was still poorly healed for calvarial defect after 3 weeks, the S/P group demonstrated the most effective bone healing with only a smaller translucent area in the fracture area. Our results confirm that SDF-1/CXCR4 signaling promotes cell migration to the central scaffold after hypoxic preconditioning and enhances bone regeneration.
Impact Statement
Current tissue engineering strategies have progressed recently from a historical approach that is based primarily on biomaterials to a cell- and tissue-based approach. Essentially all orthopedic tissue engineering strategies can be distilled to a strategy or combination of strategies that seek to increase the number or relative performance of bone-forming cells. In our study, we explored a “cell guidance” approach using a chemokine, SDF-1α, to promote cell migration to the central scaffold, to enhance bone regeneration. In our opinion, SDF-1α is a promising candidate for in situ recruitment of bone-forming cells in regenerative therapies under hypoxia precondition.
Introduction
Currently, autogenous bone grafting remains the gold standard for repair of large bone defects. This approach has several problems such as the limited availability of autogenous bone and the risk of donor site morbidity.1,2 Scaffold-based tissue engineering is an alternative approach for treating bone defects, but it is limited by the fact that the seeded cells have difficulty in infiltrating the inner scaffolds. 3
When a fracture happens, the rupture of blood vessels, activation of platelets, and secretion of tissue factors by endothelial cells can result in hematoma with hypoxia around the local tissue. 4 The CXC chemokine receptor 4 (CXCR4) is a specific receptor for the stromal cell-derived factor 1 (SDF-1). Both SDF-1 and CXCR4 are expressed in bone marrow stromal cells (BMSCs). 5 Mice with disruption to SDF-1 or CXCR4 genes die in late gestation or within an hour of birth. 6 In addition, SDF-1/CXCR4 signaling is thought to be a master regulator of stem cell migration. Several studies have demonstrated that transplanted CXCR4+ stem cells migrate to ischemic tissues. 7 CXCR4+ mesenchymal stem cells (MSCs) engrafted intravenously were able to home to the site of fracture in a rat model. 8 Increased SDF-1 expression is hence considered a key signal to promote the migration of stem and progenitor cells to these tissues to participate in bone repair and regeneration. 9 We have also demonstrated that the SDF-1/CXCR4 axis plays a crucial role in accelerating fracture healing under the condition of traumatic brain injury in rats. 10 Taken together, available data indicate that SDF-1/CXCR4 signaling has a central role in bone healing by regulating the recruitment of stem and progenitor cells. Considering the broad hypoxic microenvironments involved in bone healing, it is necessary to evaluate the contribution of hypoxia at the fracture site to the behavior of stromal cells. Furthermore, whether and how hypoxia interacts with the SDF-1/CXCR4 signaling during bone regeneration remain largely unknown.
The general approach for SDF-1 delivery in tissue engineering is direct protein incorporation into the scaffolds, which results in improved regeneration of tissue or organ defects. 11 Inspired by the critical roles of cell chemotaxis mechanism of SDF-1/CXCR4 in the fracture healing process, we hypothesized that the addition of SDF-1 along with hypoxic preconditioning may promote bone repair by facilitating stromal cell migration. To test this hypothesis, we generated and characterized the chitosan/gelatin (C/G) scaffolds, and in vitro tested the biocompatibility of the scaffolds and rat BMSC migration efficiency after SDF-1 addition and hypoxic preconditioning. Using the model of critical-sized calvarial defects in rats, we also in vivo evaluated the bone regeneration ability of the scaffolds with SDF-1 addition and hypoxic preconditioning of stromal cells.
Materials and Methods
Animals
Male Sprague-Dawley (SD) rats with body weight of ∼300 g were purchased from the animal center of Harbin Medical University, Harbin, China. All animals received humane care in accordance with “The Code of Ethics of the World Medical Association” for animal experiments (Revision of Directive 86/609/EEC). All experiments involving the use of animals were conducted with strict adherence to the protocols approved by the ethics committee of Harbin Medical University.
Isolation and culture of BMSCs
BMSCs were isolated and cultured as previously reported. 12 Briefly, SD rats were euthanized using an overdose injection of pentobarbital, and the bone marrow was flushed out from the femurs with Dulbecco's modified Eagle's medium (DMEM; HyClone). After centrifugation, the pellet was resuspended and cultured with DMEM supplemented with 10% fetal bovine serum (HyClone) in culture bottles, in a humidified atmosphere with 5% CO2 at 37°C. Two days later, the nonadherent cells were removed, while the adherent cells were further cultured and then subsequently passaged. After three passages, the cells were collected and verified to be BMSCs as reported previously. 12
Scaffold construction
Two scaffolds, the C/G scaffold and the chitosan/gelatin/SDF-1 (C/G/S) scaffold, were developed in this study. The generation of the C/G scaffold was based on our previous published method. 12 Briefly, chitosan (Biosharp, Anhui, China) was dissolved in 1% v/v acetic acid and gelatin (Biosharp) was dissolved in double-distilled water (ddw) in a 50°C water bath. The two solutions were then mixed to form 100 mL of a homogeneous solution containing 2.5% w/v chitosan and 2.5% w/v gelatin. The solution was left for cross-linking at room temperature overnight. The mixed solution was frozen at −80°C for 8 h and then lyophilized for 48 h to form the scaffold. To remove the extra acetic acid and glutaraldehyde, the scaffold was immersed in 1% w/v NaOH for 30 min, rinsed with ddw, and then immersed in 2% w/v sodium borohydride for 30 min. The C/G scaffold was then obtained after rinsing with ddw and lyophilizing again. The C/G/S scaffold was made by adding 2.5 μg of SDF-1 to 100 mL of mixed solution when preparing the C/G scaffold.
Evaluation of cell adhesion to the scaffolds
The scaffolds were cut into cylinders with a height of 0.5 cm and immersed in culture medium for 2 h. A total of 1 × 10 5 rat BMSCs in 3 mL of medium were then mixed with scaffolds of different groups and agitated overnight on a shaking table. At 3 days after culturing, cell adhesion of BMSCs was assessed. In the C/G scaffold group (C group), BMSCs were cultured under a normal oxygen condition with the C/G scaffold. In the C/G/S scaffold group (S group), BMSCs were cultured under a normal oxygen condition with the C/G/S scaffold. In the C/G/S scaffold plus hypoxic preconditioning group (S/P group), BMSCs were cultured under a 3% oxygen condition with the C/G/S scaffold. The cell adhesion ratio was calculated using the following formula: (1 × 10 5 − n1)/(1 × 10 5 ) × 100%, where n1 was the number of adherent cells on the specified scaffold.
Cell seeding and culturing on the scaffolds
Rat BMSCs at passage 5 were inoculated into different scaffolds. For cell seeding, 20 μL of suspended BMSCs at a concentration of 2 × 10 5 cells/mL was slowly dropped to one side of the scaffold. After 10 min, the scaffold was inverted and another 20 μL was dropped to the other side of the scaffold. Cell-seeded scaffolds of the C and S groups were cultured under a normal oxygen condition for 3 h and an appropriate amount of cell culture medium was added. The medium was changed every 2 days during a total of 7 days of culturing. Cell-seeded scaffolds of the S/P group were cultured under the condition with 3% oxygen for 24 h, and after medium change, the culturing was continued under the hypoxic condition for an additional 48 h. Thereafter, the scaffolds in the S/P group were transferred to a normal oxygen condition and cultured for 4 days. The medium was changed every 2 days. To examine the survival of BMSCs, live/dead staining of the cells seeded on scaffolds by LIVE/DEAD® Viability/Cytotoxicity Kit (Molecular Probes, Invitrogen) was performed as per the manufacturer's instructions.
Structural property by scanning electron microscopy
At 3 days after rat BMSC seeding, the surface and internal structures of the scaffolds were examined by scanning electron microscopy (SEM). Briefly, the scaffolds were cut and coated with gold-palladium. Morphology was visualized by a scanning electron microscope (Hitachi, S-3400n, Japan).
Hematoxylin and eosin staining
At 3, 5, and 7 days after rat BMSC seeding, hematoxylin and eosin (H&E) staining was performed following the standard procedures. Briefly, the scaffolds were fixed using polyoxymethylene (4%) for 6 h, immersed in ddw overnight, and rinsed with running water for 4 h. Dehydration was conducted through immersing the samples in a gradient alcohol solution from low to high concentrations (70% for 2 h, 80% overnight, 90% for 2 h, 95% for 1 h, 100% for 1 h, and 100% again for 30 min), and in n-butyl alcohol for 12 h. The scaffolds were subsequently immersed in xylene and liquid paraffin. The paraffin blocks were sectioned to 5 μm and stained with H&E.
Cell growth assay
At 3, 5, and 7 days after the seeding of rat BMSCs, cell growth was analyzed using the CCK-8 (Cell Counting Kit-8) assay kit (Jian Cheng, Nanjing, China) following the manufacturer's instructions. Cell growth was quantified by optical density (OD) values at 450 nm. All assays were performed in triplicate.
Western blot analysis
To determine the expression of hypoxia-inducible factor 1 α (HIF-1α) and CXCR4 in BMSCs, Western blot analysis was performed following the standard procedures. Total proteins were extracted from the cells detached from the scaffold at the indicated time points (at 1–3 days after seeding of rat BMSCs), using the M-PER Mammalian Protein Extraction Reagent (Pierce, Rockford, IL). Protein concentration was determined using the Nanodrop 1000 spectrophotometer (Thermo, Wilmington, DE). The proteins were separated on 12% SDS-PAGE (sodium dodecyl sulfate –polyacrylamide gel electrophoresis) and transferred to polyvinylidene difluoride membranes. After blocking, the membranes were incubated with anti-HIF-1α antibody (1:500 dilution; Wanleibio, Inc., China), anti-SDF-1 antibody (1:500 dilution; Wanleibio, Inc.), or anti-β-actin antibody (1:1000 dilution; Wanleibio, Inc.) at 4°C overnight. After three washes, the membranes were incubated with goat anti-rabbit horseradish peroxidase-conjugated IgG (1:5000 dilution; Wanleibio, Inc.) at 37°C for 45 min, and detected using enhanced chemiluminescence detection reagents (Thermo). Each band on the films was quantified using the ImageJ software (National Institutes of Health, Bethesda, MD) and protein levels of HIF-1α and CXCR4 were normalized to that of β-actin.
Animal model and scaffold transplantation
A total of 36 healthy SD rats were used in this study. Critical-sized calvarial defects were established as previously described. 13 After anesthesia, a sagittal incision of 1.5–2 cm was performed. The calvarium was revealed and one full-thickness defect of 5 mm was created on one side of the middle ridge using a trephine at 1500 rpm. The defects were treated with cell-seeded scaffolds immediately. Subsequently, the periosteum, muscles, and skin were sutured separately for closure.
Microcomputed tomography analysis
Animals were euthanized with an overdose of anesthetic on days 7, 14, and 21 after scaffold transplantation. The calvarium bone was harvested for evaluation. No visible inflammation and infection were observed on the harvested samples. A micro-CT (computed tomography) scanner (Scanco Medical, Bassersdorf, Switzerland) was used to examine the samples at a spatial resolution of 15 μm (500 projections/180°, 1-mm aluminum filter, 100 kV, and 100 mA). Image layers were used to reconstruct three-dimensional geometries of the samples using the VGS Studio Max software (Volume Graphics, Heidelberg, Germany).
An examiner who was blind to the grouping performed the quantitative analyses. Quantitative evaluation of bone regeneration was assessed by the percentage of bone within the surgical defect area. Measurements were carried out directly using the digital images. The commercially available software (Adobe Photoshop CS3) was used to count the number of pixels within the total defect area. The borders of the newly formed bone were manually marked on the computer screen using a digital pen. Subsequently, the pixels within the marked area were counted using the software. Based on the work from Jung et al.,
14
the area fraction of bone was calculated as follows:
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6.0 (CA). The normal distribution of the data (for Figs. 1, 2, and 3) was tested by the Shapiro–Wilk test. All data were presented as mean ± standard deviation. One-way ANOVA (analysis of variance) was used for analyzing the data in Figure 1. Two-way ANOVA was used for analyzing the data in Figures 2 and 3. The Tukey's multiple comparisons test was used for the post hoc test. Two-sided p < 0.05 was considered statistically significant.

Preexperimental observation of rat BMSC cell adhesion to the scaffolds. A total of 1 × 10 5 rat BMSCs in 3 mL of culture medium were mixed with scaffolds of different groups by mechanical force. At 3 days after culturing, cell adhesion was assessed. In the C/G scaffold group (C group), BMSCs were cultured under normal oxygen condition with the C/G scaffold. In the C/G/S scaffold group (S group), BMSCs were cultured under normal oxygen condition with the C/G/S scaffold, in which SDF-1 was added into the C/G mixed solution during cross-linking. In the C/G/S scaffold plus hypoxic preconditioning group (S/P group), BMSCs were cultured under the 3% oxygen condition with the C/G/S scaffold. n = 3 for each group. BMSCs, bone marrow stromal cells; C/G/S, chitosan/gelatin/SDF-1; SDF-1, stromal cell-derived factor 1.
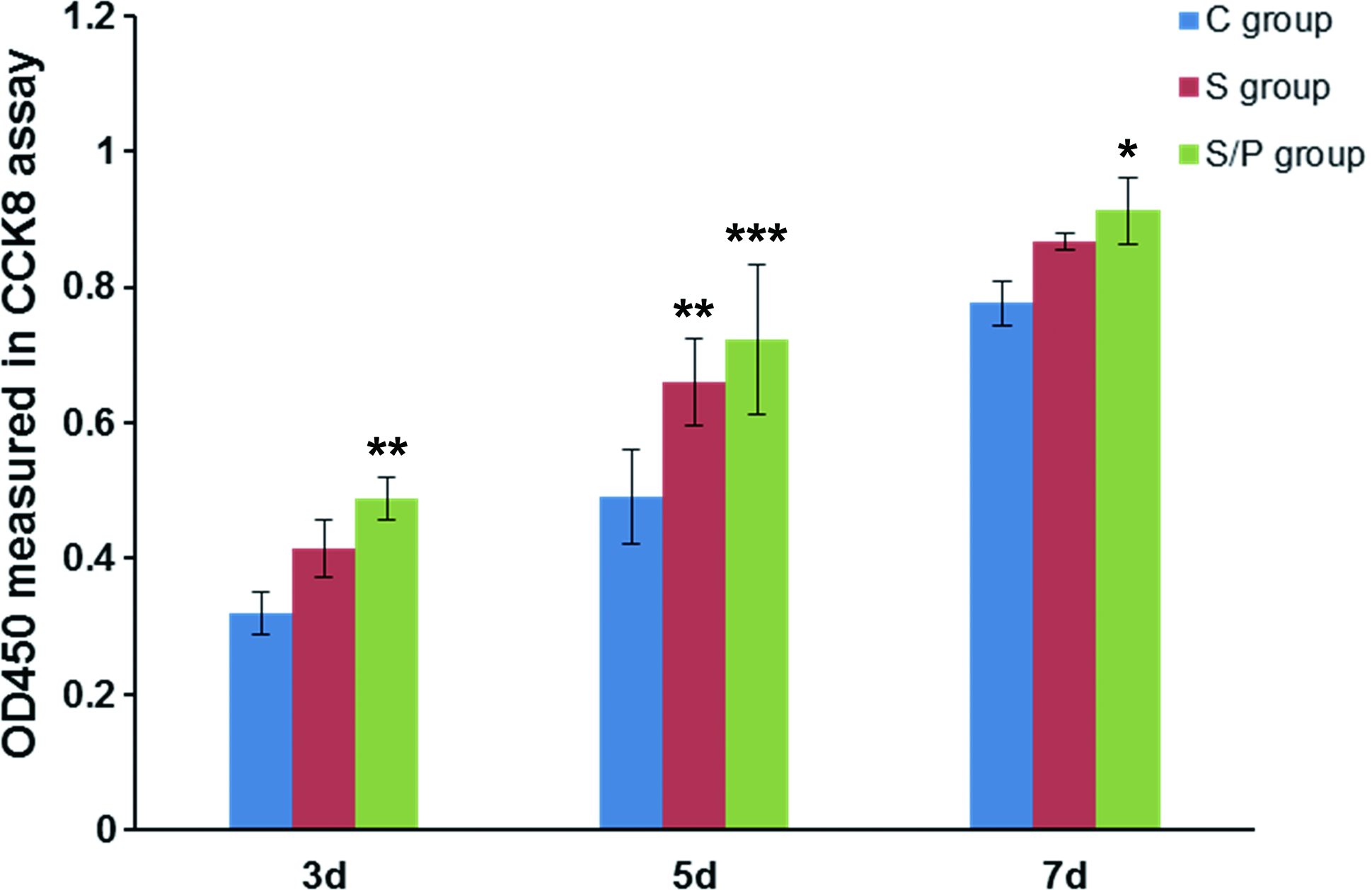
SDF-1 and hypoxic preconditioning promoted the cell proliferation of rat BMSCs seeded on the scaffolds. Experimental procedures were the same as described in Figure 5. The cell growth for each group at 3, 5, and 7 days after seeding was measured by the CCK-8 kit assay. n = 3 for each group. *p < 0.05, **p < 0.01, ***p < 0.001 versus the C group. CCK-8, Cell Counting Kit-8. Color images are available online.

Transplantation of BMSC-seeded scaffolds of the S/P group significantly enhanced the in vivo bone repair of calvarial defects in rats. Critical-sized calvarial defects were established in rats, and rats were treated with scaffolds of the C group, S group, and S/P group. The mean area fraction of the newly formed bone within the defect areas was calculated at 7, 14, and 21 days after therapy. n = 4 for each group. ***p < 0.001, versus the C group; ##p < 0.01, ###p < 0.001, versus the S group. Color images are available online.
Results
SEM analysis revealed the basic structural property of the scaffolds, which allowed effective cell adhesion
White, porous scaffolds were produced by lyophilization and cross-linking of chitosan and gelatin. First, we performed SEM to determine the surface and inner structure of the C/G scaffolds. The surface of the scaffold exhibited open pores and thin walls. Pore size was 75–90 μm (Fig. 4a). The central section of the scaffold showed well interconnected pores with size >150 μm (Fig. 4b). The scaffold was soft and elastic to some extent. Therefore, its surface was prone to be compressed during incision, which usually led to the formation of smaller pores, while the inner part of the scaffold retained larger pores. Next, we evaluated the functional properties of the scaffold in terms of supporting cell adhesion. At 3 days after seeding, rat BMSCs in the C group (23.7% ± 18.3%), S group (31.7% ± 13.5%), and S/P group (39.6% ± 9.1%) displayed comparable cell adhesion ratios (Fig. 1). These results suggested that the supplementation of SDF-1 during the cross-linking and additional hypoxic preconditioning of BMSCs did not have significant effects on cell adhesion in the scaffolds we used in this study.

Structural properties of the C/G scaffold as revealed by SEM.
Qualitative observation of cell attachment and migration behavior of BMSCs on the scaffolds
To investigate the effects of SDF-1 expression and hypoxic preconditioning on cellular behavior of BMSCs on the surface and inner sections of the scaffolds, SEM and live/death staining were performed at 3 days after cell seeding. SEM showed that cells were well spread on each scaffold (Fig. 5). However, cells grown on the scaffold of the S/P group had the best spreading (Fig. 5c). In the fluorescence live/dead staining, only live cells showed positive green fluorescence. As shown in Figure 6, more green fluorescence-positive cells were observed in the S/P group, either on the surface or in the inner part of the scaffold. We also performed H&E staining to evaluate the overall migration of the cells toward the inner sections of the scaffold. In the C group and S group, BMSCs were mainly distributed on the surface of the scaffolds, while in the S/P group, more cells were identified in the central part of the scaffold (Fig. 7). Taking together, exogenous SDF-1 and hypoxic preconditioning facilitated the migration of BMSCs in the scaffolds.
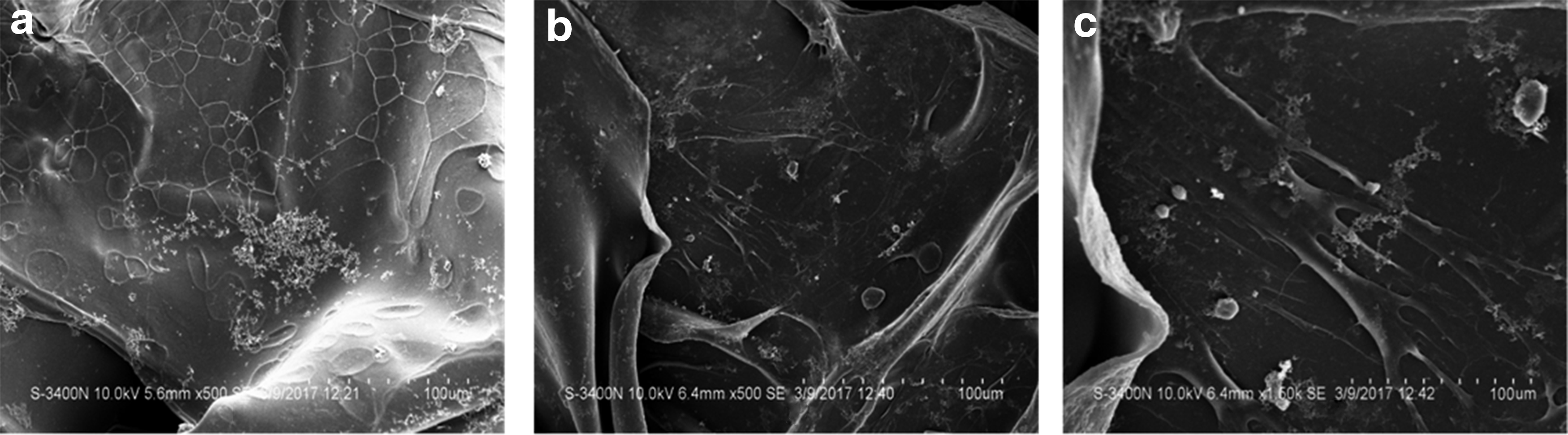
SEM analysis of the distribution and migration of rat BMSCs seeded on the scaffolds.
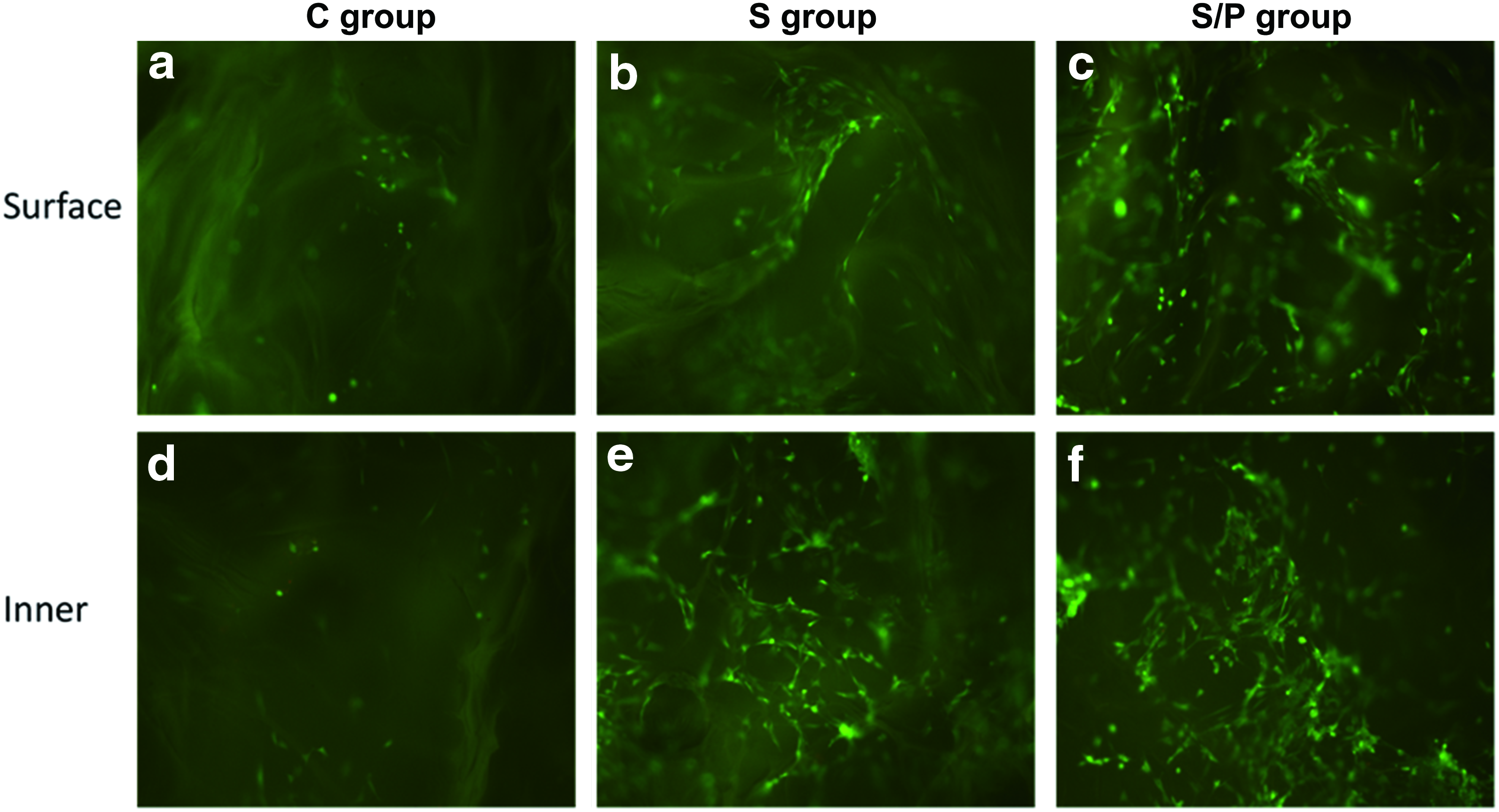
Live/dead staining of rat BMSCs grown on the surface and inner surface of the scaffolds.
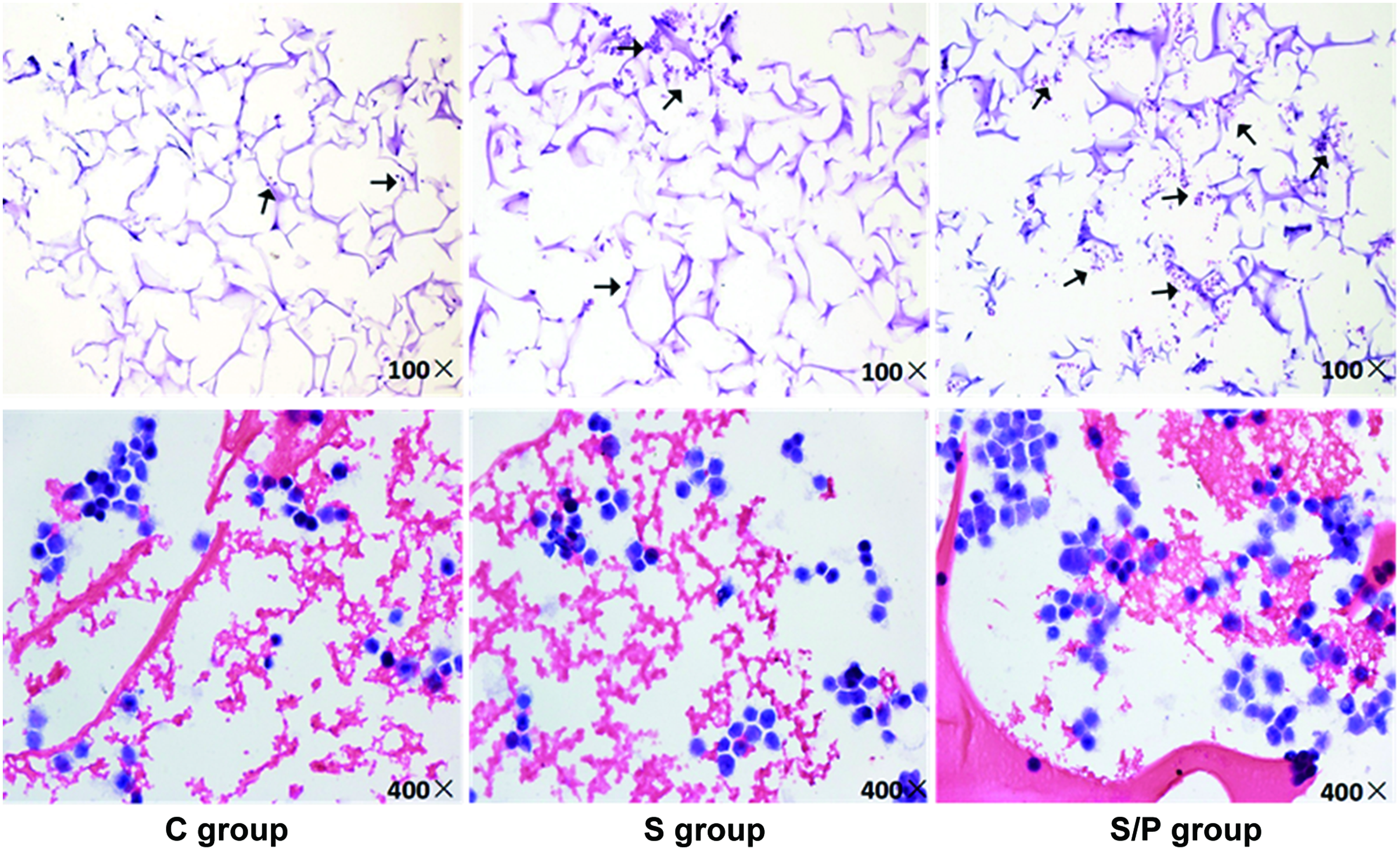
H&E staining of rat BMSCs seeded on the scaffolds. Experimental procedures were the same as described in Figure 5. At 3 days after seeding, rat BMSCs were subjected to H&E staining to examine their distribution and migration. Top: original magnification 100 × . Bottom: original magnification 400 × . Black arrows indicate cells adhered to the scaffolds. H&E, hematoxylin and eosin. Color images are available online.
SDF-1 and hypoxic preconditioning promoted the cell proliferation of rat BMSCs seeded on the scaffolds
To further substantiate the promoting role of SDF-1 and hypoxic preconditioning on cell viability, we measured cell growth by CCK-8 kit assay at 3, 5, and 7 days after BMSC seeding. On day 3, the OD values that were indicative of cell viability were 0.32 ± 0.03, 0.42 ± 0.04, and 0.49 ± 0.03 in the C group, S group, and S/P group, respectively (S/P group vs. C group, p < 0.01). On day 5, the OD values were 0.49 ± 0.07, 0.66 ± 0.06, and 0.72 ± 0.11, respectively (S group vs. C group, p < 0.01; S/P group vs. C group, p < 0.001). On day 7, the OD values were 0.78 ± 0.03, 0.87 ± 0.01, and 0.91 ± 0.05, respectively (S/P group vs. C group, p < 0.05) (Fig. 2). Therefore, at all the time points we tested, BMSCs grown in the scaffolds of the S/P group had a better cell viability than BMSCs in the other two groups.
SDF-1 and hypoxic preconditioning markedly elevated the protein levels of HIF-1α and CXCR4 in BMSCs seeded on the scaffolds
To evaluate whether the SDF-1/CXCR4 axis was activated by hypoxia, we performed Western blot assays to determine the protein levels of HIF-1α and CXCR4 in BMSCs at the early stage of cell/scaffold culturing. Data on protein levels were expressed as fold increases relative to the C group on day 1. As shown in Figure 8, HIF-1α levels in the S group were higher compared to the C group at each time point (C group: 1-fold on day 1; 1.08-fold on day 2; and 1.09-fold on day 3; S group: 1.48-fold on day 1; 1.64-fold on day 2; and 1.63-fold on day 3). More remarkably, HIF-1α levels in the S/P group were much higher compared to the S group and C group (S/P group: 2.47-fold on day 1; 2.63-fold on day 2; and 2.67-fold on day 3). Consistently, similar trends of CXCR4 expression in BMSCs of different groups were observed. At all the same time points checked, CXCR4 was more expressed in the S group (C group: 1-fold on day 1; 0.85-fold on day 2; and 1.01-fold on day 3; S group: 1.47-fold on day 1; 1.40-fold on day 2; and 1.17-fold on day 3), and the most expressed in the S/P group (S/P group: 3.66-fold on day 1; 4.04-fold on day 2; and 3.18-fold on day 3) (Fig. 9). Our results indicated that the expression levels of both HIF-1α and CXCR4 were markedly higher in BMSCs seeded on the scaffolds of the S/P group compared with the two other groups.

SDF-1 and hypoxic preconditioning promoted HIF-1α protein expression in rat BMSCs seeded in the scaffolds. Experimental procedures were the same as described in Figure 5. The protein expression of HIF-1α in rat BMSCs at 1, 2, and 3 days after seeding was detected by Western blot, and the expression levels were normalized to that of β-actin. HIF-1α, hypoxia-inducible factor 1 α.

SDF-1 and hypoxic preconditioning promoted CXCR4 protein expression in rat BMSCs seeded in the scaffolds. Experimental procedures were the same as described in Figure 5. The protein expression of CXCR4 in rat BMSCs at 1, 2, and 3 days after seeding was detected by Western blot, and the expression levels were normalized to that of β-actin. CXCR4, CXC chemokine receptor 4.
Transplantation of BMSC-seeded scaffolds of the S/P group significantly enhanced the in vivo bone repair of calvarial defects in rats
To further confirm the advantages of SDF-1 supplementation and hypoxic preconditioning in bone regeneration, we established the animal model of critical-sized calvarial defects in rats, and evaluated the effects of different scaffolds on in vivo bone repair. Bone regeneration within the defects was three dimensionally reconstructed using micro-CT. As shown in Figure 3, the mean area fraction of the newly formed bone within the surgical defect was about 8.85% ± 1.36% in the C group, 21.05% ± 3.45% in the S group, and 31.91% ± 2.79% in the S/P group at 7 days after surgery. At 14 days after surgery, the average area fraction of newly formed bone within the surgical defect was 21.13% ± 2.57% in the C group, 32.37% ± 2.27% in the S group, and 40.61% ± 3.88% in the S/P group. At 21 days after surgery, the average area fraction of newly formed bone was 30.23% ± 2.25% in the C group, 42.59% ± 2.89% in the S group, and 61.65% ± 5.30% in the S/P group. At all time points we checked, the C group presented significantly less bone formation compared to the S and S/P groups (all p < 0.001), while the bone formation in the S/P group was always better compared with the S group (p < 0.01 or p < 0.001). While the C group was still poorly healed after 3 weeks (Fig. 10a–c), the scaffolds of the S/P group had a much smaller translucent area (Fig. 10g–i), and the defect in the S/P group was almost fully repaired after 3 weeks (Fig. 10i). In addition, bone formation in the S/P group progressed evidently in a centripetal manner with new bone stretching from the margins toward the center. Therefore, BMSC-seeded scaffolds of the S/P group had the best performance, and significantly enhanced the in vivo bone repair of calvarial defects.

Representative micro-CT images showing the effects of different scaffolds on bone formation in rats with calvarial defects.
Discussion
One of the major challenges in cell scaffold-based tissue engineering is to seed the cells inside the scaffold as homogeneously as possible. Usually, the cells that form a tissue layer on the outer surface significantly inhibit the growth of the cells inside the scaffold, which is a major hurdle to effective tissue engineering. 5 In this study, we generated the scaffold by cross-linking of chitosan and gelatin, and confirmed that the supplementation of SDF-1 in the scaffolds together with preconditioning of rat BMSCs significantly enhanced the migration of BMSCs to the inner sections of the scaffolds. Our study suggests that the hypoxic preconditioning-induced activation of the SDF-1/CXCR4 signaling plays an essential role in mediating the successful adhesion and migration of BMSCs in the scaffolds.
The SDF-1/CXCR4 axis has been revealed to play an important role in the recruitment and retention of progenitor cells (PGCs) in ischemic tissues for promoting tissue regeneration. 15 In our previous study, we locally delivered SDF-1 to fractured sites and intravenous engrafted BMSCs in rats under the condition of traumatic brain injury. Using cell fluorescence labeling, we observed that CXCR4-positive cells can home to the fractured site and accelerate healing due to the SDF-1 gradient. 10 Based on these results, we hypothesized that the combination of the scaffold and SDF-1/CXCR4 axis may provide an effective method to promote scaffold cellularization. Scaffolds made of C/G provide good microenvironments for cell attachment and growth due to the pore structures. Hence, C/G scaffolds were selected in this study to determine whether hypoxic preconditioning and the SDF-1/CXCR4 axis were beneficial for scaffold cellularization and in vivo bone regeneration.
In the preexperimental observation of cell adhesion to the scaffolds (Fig. 1), the experiment was done by mechanically forcing the BMSCs to adhere to the scaffold, and there was no significant difference among the three groups. Nevertheless, when the cells were dropped to the scaffolds, their attachment to the scaffold occurred in the absence of the exogenous forces. In that case, differences were observed among the three groups regarding the attachment, migration, and growth of BMSCs on the different scaffolds (Figs. 2, 5, and 7). The better performance of scaffolds in the S/P group may be due to the upregulation of adhesion molecules when cells are under hypoxic stress. 16
Our live/dead staining demonstrated that rat BMSCs survived after seeding to the scaffolds of the three groups, demonstrating that the experimental operations can effectively remove the glutaraldehyde residues on the scaffold and avoid their cytotoxicity. Nevertheless, the cells in the S/P group had better viability and higher cell growth rate than cells in the other groups. We speculated that, on one hand, this was due to the activation of hypoxia-induced SDF-1/CXCR4 signaling, which promoted the stable adhesion of BMSCs to the scaffold and thus provided essential basis for cell growth; on the other hand, hypoxic pretreatment itself may accelerate the survival and proliferation of stem cells and play cytoprotective effects, as previously reported. 17
Hypoxic preconditioning has been used as an approach to enhance PGC survival, adhesion, and migration in vitro. 18 Rosová et al. 17 demonstrated that exposure of MSCs to hypoxia activated the Akt signaling pathway to preserve cell viability and cell cycle rates. Hu et al. 19 and Pasha et al. 20 showed that hypoxic preconditioning improved the myocardium repair capabilities of MSCs. It is well known that hypoxia upregulates both HIF-1α and CXCR4 expression in cells such as endothelial cells, monocytes, and BMSCs, through HIF-1α activation and transcript stabilization, and increases the chemotactic response to SDF-1. Tang et al. 21 demonstrated that hypoxic preconditioning increased CXCR4 expression in an HIF-1α-dependent manner. Dai et al. 22 showed that exposure of endothelial progenitor cells to hypoxia upregulated CXCR4 expression and enhanced chemotaxis. Marquez-Curtis and Janowska-Wieczorek 23 showed that the SDF-1/CXCR4 axis can be manipulated to improve cell recruitment and tissue repair. All of these findings are consistent with our results. Our Western blot analysis showed that HIF-1α and CXCR4 protein levels were highly expressed in the S/P group. Nevertheless, as reviewed by Lin et al., 24 a number of factors can be used to modulate the migration of MSCs. Therefore, to optimize the migration process, those additional factors should be further explored in combination with SDF-1 in future studies.
In addition, we evaluated the bone repair ability of the scaffolds in vivo. In this study, days 7, 14, and 21 were selected as the time points for micro-CT analysis, as previous studies have indicated that the contribution of circulating cells to fracture callus peaked at 2 weeks and declined rapidly at 3 and 4 weeks. 25 Our results indicated that there was obvious bone formation at 7 days. At 14 days, the defect area was almost covered with new bone in the S/P group, which reflects a significantly stronger osteogenic ability compared with the other two groups. There may be three reasons for the enhanced bone regeneration ability by hypoxic preconditioning and SDF-1 signaling. First, SDF-1 may increase the survival rate of transplanted cells. In ischemic tissue, cells encounter severe hypoxic conditions, ranging from 0.4% to 2.3% O2, which often results in cell apoptosis and thus limits the therapeutic potential. Nevertheless, these lethal effects may be prevented by mild hypoxic pretreatment, that is, 2–3% O2, for a short period of time before transplantation into ischemic tissues. Previous studies have demonstrated that severe hypoxia (0.5% O2) can lead to apoptosis.17,19 Second, SDF-1 delivered to the scaffold leads to high SDF-1 concentrations and hence establishes a gradient between the local site and surrounding tissues. This induces host stem cell mobilization and homing to the affected site. 26 Moreover, SDF-1 promotes endothelial progenitor cell migration to the ischemic tissue for local vascularization, which is necessary for bone regeneration. Nevertheless, the exact mechanisms for promoting bone regeneration by SDF-1 are unclear and need to be elucidated before its clinical applications.
This study has some limitations and much work is still needed before any clinical application. We assumed that the release kinetics of SDF-1 was the same under normal and low oxygen conditions. Hypoxic pretreatment should not affect the concentration gradient formed by SDF-1 since the atmospheric pressure was the same in both groups. We will examine it in future studies to determine whether this could be a modifiable factor affecting the efficacy of the scaffold. Second, the conditions of hypoxia used in previous studies were 0.5% O2 for 24 h,17,19 but we worried that severe hypoxia may induce cell apoptosis. Therefore, we chose mild hypoxia (3% O2) and appropriately prolonged the time of treatment (3 days). Third, H&E staining was performed to roughly evaluate the migration of BMSCs into the inner parts of the scaffold. In the S/P group, the migration ability of BMSCs into the inner parts of the scaffold was found enhanced. As for the extent to which this migration ability was specifically enhanced, quantitative experiments will have to be performed. Fourth, the micrographs in Figure 5 show differences in cell size among groups. We speculate that because SDF-1 was not present in the C group, a signal that mediated cell adhesion was lacking. As a result, a large number of cells were apoptotic in the C group due to inability to adhere, and the remaining cells were not stably attached to the scaffolds. Therefore, these cells in the C group showed smaller size and impaired proliferation.
In conclusion, our work demonstrated that under hypoxic preconditioning, SDF-1 markedly promoted cell migration to the central sections of the C/G scaffold in vitro and enhanced bone regeneration in vivo. Our study suggests that hypoxic preconditioning in combination with exogenous SDF-1 supplementation is a promising approach for scaffold cellularization.
Footnotes
Acknowledgment
This work was supported by grants from the National Natural Science Foundation of China (81501868).
Disclosure Statement
No competing financial interests exist.