
Abstract
We induced tissue repair following ischemic stroke in rats by delivering brain-derived neurotrophic factor (BDNF) directly to the brain. This protein is a potent modulator of plasticity and neuroprotection in the developing and adult central nervous system; however, the therapeutic potential of BDNF has been largely thwarted by its inability to cross the blood–brain barrier at an effective concentration. Herein, we demonstrate enhanced acute recovery of forepaw dexterity and enhanced hindlimb function at 7 weeks postinjury by delivering BDNF locally, with sustained release in vivo for up to 21 days. Using an encapsulation-free methodology, BDNF was dispersed in a hydrogel composed of hyaluronan and methylcellulose with poly(lactic-co-glycolic acid) nanoparticles and this composite was deposited epicortically, directly above the stroke lesion. BDNF delivery augmented plasticity, as evidenced by synaptophysin staining in the contralesional hemisphere of BDNF-treated rats, and presence of the vehicle reduced the lesion volume and prevented neuron loss in perilesional tissue. With local, sustained delivery directly to the brain, we demonstrate the benefit of BDNF in the treatment of stroke injury in a rodent model.
Impact Statement
We developed a biocomposite that can be mixed with brain-derived neurotrophic factor (BDNF) and dispensed onto the surface of the brain to provide sustained, local release of the protein using a procedure that avoids additional damage to neural tissue. The composite is simple to fabricate, and provides sustained release without nanoparticle encapsulation of BDNF, preserving material and protein bioactivity. We demonstrate that when delivered epicortically to a rat model of stroke, this composite allows BDNF to diffuse into the brain, resulting in enhanced behavioral recovery and synaptic plasticity in the contralesional hemisphere.
Introduction
Traumatic neurological events, such as stroke, activate plastic processes in the brain to promote limited recovery 1 ; yet, neurological deficits persist in the majority of stroke survivors2–4 due to the insufficiency of endogenous plasticity. It is hypothesized that enhancing neuroprotection and synaptic plasticity would lead to improved patient recovery as a result of improved neural networks. Brain-derived neurotrophic factor (BDNF) is a neuroprotective agent and a key regulator of plasticity in the healthy and injured brain. Upon binding to its receptor, tyrosine receptor kinase B (TrkB), several cascades are initiated that promote cell survival and differentiation, synaptogenesis and long-term potentiation (LTP), and upregulation of BDNF expression.5–8 Studies linking increased BDNF expression to physical activity 9 and rehabilitation practices10,11 have established the importance of its presence during the recovery of motor function after stroke. Immediately following stroke, BDNF expression is upregulated both in the ipsilesional and contralesional brain hemispheres; however, this increase in expression is short lived, 12 suggesting an opportunity for delivery of exogenous BDNF for brain repair.
Ploughman et al. 13 substantiated the role of BDNF in functional recovery by blocking BDNF expression following ischemic injury. Delivery of a BDNF antisense oligonucleotide to the contralesional hemisphere through an infusion pump for 28 days, beginning 4 days postinjury, negated the beneficial outcomes of rehabilitation therapy that are normally observed. 13 A similar effect was observed when BDNF was blocked after injury with a TrkB-Fc-loaded hydrogel that sequestered the BDNF present at the lesion site. 14 In the context of learning and memory, there is mounting evidence that BDNF is responsible for the formation and maintenance of LTP, a strengthening of synapses that contributes heavily to memory formation and storage.15,16 Patterson et al. 17 perfused recombinant BDNF in BDNF+/− mouse hippocampal slices and rescued LTP. Taken together, these experiments show that BDNF is necessary for recovery and exogenous BDNF can influence synaptic plasticity.
BDNF is unable to cross the blood–brain barrier (BBB) due to its large size and short half-life.18,19 This presents a significant challenge for the delivery of effective concentrations of BDNF to the peri-infarct tissue. Some groups have modified this protein with BBB-targeting vectors20,21 or incorporated it into polyion complexes that accumulate in the brain vasculature surrounding the lesion, 18 while others have used intraventricular injections or infusions to bypass the BBB.22,23 Direct injection of BDNF into the lateral ventricle demonstrated some beneficial effects; however, multiple injections, high dosing, or neural implants are usually required to maintain effective concentrations in the parenchyma and are highly invasive. 24 Transplantation of cells that express BDNF is another option, 25 but cell delivery to the central nervous system (CNS) is rife with challenges, including cell survival and/or uncontrolled proliferation, and requires direct tissue injection. 26 Off-target side effects associated with systemic delivery, increased risk of infection, and invasive procedures associated with tissue injection greatly limit the therapeutic efficacy of these approaches and call for a more refined methodology. 27
Biomaterials can offer an alternative to repeated bolus injections by controlling the release of a therapeutic molecule and facilitate less-invasive delivery practices. Both synthetic and natural polymers have been used to achieve sustained release of neurotrophins to varying degrees,28–34 although few studies have demonstrated sustained local release of BDNF in a stroke model. In a recent study by Cook et al., 35 a hyaluronan poly(ethylene glycol) crosslinked hydrogel was injected into stroke-injured brain tissue to sustain the release of BDNF in mice over a period of 3 weeks. At 9 weeks postinjury, there was an increase in axonal sprouting in the contralesional striatum of BDNF-treated animals and some motor map reorganization. 35 Despite the positive outcomes observed with BDNF delivery, there are broad challenges associated with direct injection of hydrogels into brain tissue. By their very nature, hydrogels swell and this can cause further tissue damage when injected directly into brain tissue. Since this hydrogel will swell by 20%, the 6 μL injected will occupy a volume of 7.2 μL until it begins to degrade. 36 While these studies were performed in mice, making functional recovery challenging to assess, 37 they highlight the importance of developing a less-invasive paradigm for BDNF delivery.
We developed a biomaterial-based strategy for local delivery of BDNF that avoids additional damage to neural tissue and allows sustained delivery of unencapsulated, bioactive protein to the brain. Our delivery vehicle consists of a hyaluronan and methylcellulose (HAMC) hydrogel in which we disperse poly(lactic-co-glycolic acid) (PLGA) nanoparticles. The HAMC hydrogel is shear thinning and inverse thermal gelling, allowing injection through a fine-gauge needle and in situ gelation at physiological temperature. 38 The anti-inflammatory action of the high molecular weight sterile sodium hyaluronate (HA) itself results in neuroprotection and decreased lesion volume. 39 Encapsulation of protein therapeutics in polymer nanoparticles is a commonly used method to achieve sustained release; however, the delicate nature of proteins makes them highly susceptible to denaturation and loss of bioactivity during this harsh process. Our system obviates the need for encapsulation by harnessing the electrostatic interaction between the positively charged BDNF and the negatively charged PLGA particle. By simply mixing the BDNF into the PLGA-HAMC composite, we created an easy and efficient system, well suited for in vivo application. 40 We deposited this BDNF-loaded composite on the surface of the cortex above the cortical stroke lesion, thereby avoiding injection into tissue and any possibility of additional damage associated with swelling. We show the efficacy of this novel epicortical delivery strategy in an endothelin-1 (Et-1) rat model of stroke in terms of tissue repair, behavioral recovery, and BDNF diffusion in stroke-injured rat brain tissue. Notwithstanding these exciting results, we acknowledge that BDNF will have to diffuse to greater distances in brain tissue with epicortical versus direct tissue injection, particularly in cases where the lesion is deeper than the cortex.
Materials and Methods
All reagents were acquired from Sigma-Aldrich (Oakville, Canada), unless otherwise indicated.
PLGA nanoparticle synthesis
PLGA nanoparticles were synthesized by water/oil/water double emulsion-solvent evaporation, as previously described, 40 in a laminar flow biosafety cabinet with sterile technique and materials. Briefly, 120 mg of PLGA (Lactel Absorbable Polymers; Durect Corporation; 50:50 lactide:glycolide, carboxy terminated, 0.15–0.25 dL/g in hexafluoroisopropanol [HFIP]), 4 mg of magnesium carbonate, and 0.05% Pluronic® NF-127 (BASF, Mississauga, Canada) were dissolved in 900 μL dichloromethane (DCM; Caledon Labs, Georgetown, Canada). One hundred microliters of 120 mg/mL bovine serum albumin (BSA) in artificial cerebrospinal fluid (aCSF) (149 mM NaCl, 3 mM KCl, 0.8 mM MgCl2, 1.4 mM CaCl2, 1.5 mM Na2HPO4, and 0.2 mM NaH2PO4, pH 7.4) was added and vortexed for 10 s to achieve 10% w/w BSA/PLGA in the final formulation. The mixture was then sonicated on ice for 10 min. This primary emulsion was added to 3 mL of a 2.5% w/v solution of poly(vinyl alcohol) (PVA) (30–70 kg/mol), vortexed, and sonicated on ice for 10 min. The secondary emulsion was added to a hardening bath of PVA and stirred overnight to allow the DCM to evaporate. The nanoparticles were washed four times by ultracentrifugation, lyophilized, and stored at −20°C.
Preparation of BDNF-loaded PLGA-HAMC composite
To prepare the BDNF-loaded PLGA-HAMC composite, PLGA nanoparticles were first dispersed in aCSF (0.1% BSA) to achieve 10% w/v in the final formulation. The nanoparticle solution was vortexed and bath sonicated for 5 min to ensure proper dispersion of the particles. Sterile methyl cellulose (MC, 300 kg/mol; Shin-Etsu, Tokyo, Japan) and HA (1.4–1.8 × 106 g/mol; NovaMatrix, Sandvika, Norway) were dissolved in the nanoparticle solution using a dual asymmetric centrifugal mixer (FlackTek, Landrum, SC) to a final concentration of 1.4% w/v HA and 3% w/v MC. Complete mixing was achieved by alternating vertical and horizontal speed mixing until composite appeared homogenous and was then stored overnight at 4°C. BDNF was added to the top of the gel to give a final concentration of 0.33 μg/μL in the composite and mixed using a protocol optimized to maintain bioactivity; after the addition of BDNF, the composite was speedmixed vertically for 10 s, horizontally for 10 s, centrifuged at 4°C for 10 s, and speedmixed vertically again for 10 s.
Animal approval
All animal work was approved by the University of Toronto Animal Care Committee and was conducted in accordance with the Guide to the Care and Use of Experimental Animals (Canadian Council on Animal Care). Male Sprague Dawley rats, 8–10 weeks old, and weighing between 225 and 350 g were used (Charles River, Quebec City, Canada). A total of 91 animals were used in these studies.
Behavioral task training
To assess forelimb dexterity, the Montoya staircase task was used. Animals were trained twice per day for 14 days before surgery to retrieve pellets with both paws from the staircase apparatus, as described by Ploughman et al. 41 Of the 21 pellets accessible to each forelimb, each animal needed to reach, grasp, and eat 15 with at least one forelimb to be considered fully trained and included in the study. The last four trials, 2 days before surgery, were averaged to generate baseline performance values expressed as a percentage of the maximum number of pellets that could be reached (i.e., 21). Handedness was determined based on the best-performing forelimb and the injury was thus given on the dominant side. The investigators were blinded and animals were sorted into the treatment and control groups so that average skill level remained consistent between the groups. The BDNF treatment group (n = 17) received the vehicle with BDNF mixed into the composite at the time of injury. The vehicle group (n = 18) received only the composite at the time of injury to control for effects of the hydrogel and nanoparticles. The injury group (n = 15) controlled for the effects of the injury alone. The sham group (n = 11) was included to control for effects of the anesthetic administration, scalp incision, and cap placement procedure.
Et-1 stroke and drug delivery
Animals were placed in groups so that the average animal skill at the behavioral tasks was constant between groups. Rats were first anesthetized with isoflurane, shaved, and secured in a stereotaxic apparatus (Kopf Instruments, Tujunga, CA). An incision was made in the scalp and a retractor was inserted to hold the incision open. Sham animals were fitted with two curved 5.9 mm diameter polycarbonate disks on the surface of the skull, secured with bone glue (Loctite 454; Henkel Corporation, Rocky Hill, CT), and the wound sutured closed. In all other groups, a burr hole with a diameter of 2.7 mm was drilled using a trephine drill bit (Cat. No. 18004-27; Fine Science Tools, Inc., Vancouver, Canada), centered at anteroposterior (AP) +1.15 mm and mediolateral (ML) +3.0 mm. Et-1 (human/porcine, 1 μg/μL in ddH2O; Cat. No. ab120471; Abcam, Cambridge, MA) was loaded into a 10 μL Hamilton syringe with a 26 G, 45° bevel needle (Model 1701 RN; Hamilton) and injected using an automated pump (Pump 11 Elite Nanomite; Harvard Apparatus, Saint-Laurent, Canada). Two injections were made in the primary motor cortex and one in the striatum for a total of three Et-1 injections:
AP +2.3 mm, ML ±3.0 mm, dorsoventral (DV) − 2.2 mm AP 0 mm, ML ±3.0 mm, DV − 2.2 mm AP +0.7 mm, ML ±3.8 mm, DV − 6.9 mm
ML and AP coordinates are relative to bregma and dorsoventral coordinates are relative to the brain surface. The needle was lowered an additional 0.1 mm DV at each injection site and then raised DV +0.1 mm for the final DV coordinate. Once in position, the needle was left for 1 min to equilibrate. Following equilibration, Et-1 was injected at 0.5 μL/min until a volume of 1 μL had been injected. Following a pause of 1 min, an additional 1 μL was injected at the same rate. Once the injection was complete, needles were allowed to equilibrate for 3 min and then slowly withdrawn to prevent Et-1 backflow. A durectomy was performed using a surgical microscope to expose the cortical surface. For the injury group, a 2.3 mm diameter circle of medical-grade silicone sheeting (Cat. No. CUST-20001-005; BioPlexus, Ventura, CA) was placed on the tissue and a small piece of Surgifoam® gelatin sponge (Cat. No. 1972; Ethicon, Markham, Canada) was soaked in saline and placed in the trephine hole on top of the silicone sheeting. This was done to prevent soft tissue adhesion and swelling into the space created by removing the skull. Two 5.9 mm diameter polycarbonate disks were secured over the burr hole with bone glue and the entire assembly covered with Ortho-Jet™ BCA dental cement (Cat. No. 1334CLR; Lang Dental, Wheeling, IL).
Surgeries for the vehicle and BDNF groups followed the same procedure, with the only difference being the addition of BDNF to the composite formulation for the BDNF animals (2 μg/animal). Following durectomy, these groups were fitted with a 5.9 mm polycarbonate disk with a concentric 2.7 mm hole, secured over the burr hole with bone glue. They then received 6 μL of PLGA-HAMC composite, deposited directly onto the surface of the cortex in the space created by skull and disk. A second 5.9 mm polycarbonate disk with no opening was secured over top of the first disk with bone glue, the entire assembly was covered with dental cement (Ortho-Jet™; Lang Dental Manufacturing Company, Wheeling, IL), and the skin was sutured closed.
Behavioral task testing
Staircase testing was conducted for three consecutive days, twice per day, every other week (6 trials per time point). The last four trials were averaged to give the animals' scores at each time point, discarding the first two trials due to animal re-acclimation to the task. The first testing period began 4 days poststroke (week 1) to allow a deficit to be observed in injured animals before BDNF was expected to begin releasing from the composite. This injury baseline was expressed as a percentage of the uninjured baseline performance and animals were included in further analysis if they had a performance between 20% and 70% of uninjured baseline at week 1 testing as this indicated a sufficient deficit in the staircase task. 41
For the tapered beam task, animals were trained to cross a 1 m tapered beam elevated off the floor to reach a darkened goal box at the far end containing food pellets. Foot slips onto the ledge on either side were counted as faults if the limb appeared to be weight-bearing. Runs were recorded using a video camera and later analyzed in a blinded manner. Animals were required to traverse the beam such that the number of steps in the runs totaled >40 steps, usually taking between 4 and 6 runs. Animals were trained for 2 days before surgery, with the last day serving as the baseline uninjured performance. Animals were tested on the tapered beam task biweekly. Animals were included in behavioral analysis for the tapered beam task if they exhibited at least a 10% hindlimb slippage at 1 week postinjury and did not experience major complications during or after surgery. Within-group testing using a one-way analysis of variance (ANOVA) confirmed that the animals selected with our inclusion criteria exhibited a significant decline in performance at week 1 compared to their uninjured baseline performance.
Brain tissue preparation for histological analysis
Animals were sacrificed after the 7-week behavioral testing and brains were extracted and fixed in 4% paraformaldehyde at 4°C for 10 days, followed by cryoprotection in 15% and then 30% sucrose. Brains were snap frozen and cryosectioned coronally (30 μm).
Immunohistochemistry
Sections stained for synaptophysin (syn) and NeuN were first permeabilized for 15 min (1% Triton X-100 in phosphate-buffered saline [PBS]) at room temperature, blocked for 30 min (0.1% Triton X-100 and 5% BSA in PBS), and incubated with rabbit anti-synaptophysin (1/400 dilution in blocking solution, ab23754; Abcam, Cambridge, MA) primary antibody, and mouse anti-NeuN (1/500 dilution in blocking solution, MAB377; Millipore, Inc., Billerica, MA) primary antibody at room temperature for 2 h. Sections were then washed thrice for 5 min in PBS before incubation with Hoechst 33342 nuclei stain (1 μg/mL in blocking solution, Cat. No. 62249; Invitrogen, Inc., Burlington, Canada) and the highly cross-adsorbed secondary antibodies AlexaFluor 488 goat anti-rabbit IgG (1/500 dilution in blocking solution, A11034; Invitrogen, Inc.) and AlexaFluor 546 goat anti-mouse (1/500 dilution in blocking solution, A11030; Invitrogen, Inc.) for 1 h at room temperature. Sections were washed thrice for 5 min in PBS and mounted with ProLong® Gold Antifade Mounting Medium (Cat. No. P36934; Thermo Fisher Scientific, Mississauga, Canada).
Imaging and quantification
Coronal sections spaced 300–600 μm apart were chosen between AP +4.0 mm and AP −1.0 mm relative to bregma to assess the effects of local BDNF delivery on the tissue. Images were taken with a 20 × objective on an Axio Scan.Z1 Slidescanner (Zeiss, Oberkochen, Germany), using consistent microscope settings for each section. Lesion area was defined as the area lacking NeuN+ staining, using the contralesional hemisphere as a reference, and the tracing of the lesion enabled the selection of regions of interest (ROI) for syn and NeuN staining.
NeuN staining was employed to assess the number of mature neurons in the peri-infarct tissue, using the same ROI as for syn quantification. The average size of a neuron in NeuN+ pixels was attained by sampling 100 randomly chosen neurons across 10 sections from different animals, which were traced and analyzed in ImageJ. The NeuN+ pixels in R1 and R2 were divided by this average to estimate the number of neurons present. To determine the lesion size, ImageJ software was used to measure the traced lesion area for 10 sections per animal. These areas were first multiplied by the intersection AP distance to determine the average volume of damage between sections and then added to determine total lesion volumes. This was done for both cortical and striatal lesions (SLs) separately, and these volumes added to give total lesion volume for each animal.
As a measure of synaptic plasticity, syn+ pixels were quantified in six sections per animal using Fiji ImageJ Software. Images were first converted to 8-bit and thresholded using the “Moments” thresholding algorithm, and then four 500 × 500 μm cortical ROI were chosen in both hemispheres. For the ipsilesional hemisphere, two ROI were defined directly adjacent to the lesion on either side of the cavity and were averaged to give the R1 values reported. An additional two ROI, 500 μm away from the lesion on either side of the cavity were averaged and reported as R2 values (Supplementary Fig. S1). Homotopic regions in the contralesional hemisphere were quantified in the same manner.
Analysis of BDNF diffusion in stroke-injured brain tissue
To determine BDNF concentrations in the brain after local delivery, surgeries were performed as described above. Animals were sacrificed after 4, 7, and 21 days postinjury and the brain tissue was harvested, snap frozen, and stored at −80°C. Tissue sections were prepared for diffusion analysis, as previously described. 43 Briefly, six serial 1-mm-thick coronal sections surrounding the stroke lesion were cut using a McIlwain tissue chopper (Mickle Laboratory Engineering Company, Surrey, UK). A Leica CM3050S cryostat system was then used to section these coronal slices dorsoventrally, each 0.3 mm thick. Tissue from each coronal section at the same depth was collected together in preweighed 2 mL polystyrene microtubes and homogenized to give average BDNF concentration at each depth spanning 300 μm, down to 3 mm below the surface of the tissue.
To extract BDNF from the tissue, 200 μL of homogenization buffer (20 mM Tris-HCl buffer pH 8, 137 mM NaCl, 1% vol/vol Triton-X 100, and 10% vol/vol glycerol) and 3–5, 1.0 mm zirconia beads (Cat. No. 11079110zx; Biospec Products, Bartlesville, OK) were added to each microtube. Tissue was homogenized for 30 s with a Mini-beadbeater 16 (Biospec Products), centrifuged at 4°C for 5 min at max speed, homogenized for another 30 s, and centrifuged a final time at 4°C for 15 min at max speed. Supernatant from the tubes was collected and an enzyme-linked immunosorbent assay (ELISA) (BDNF Emax® ImmunoAssay Systems; Promega, Madison, WI) was used to quantify BDNF in each segment of the tissue. Tissue from ipsilesional vehicle control brains was spiked with BDNF to generate the ELISA standard curve so as to account for endogenous BDNF levels and any effect of processing.
Statistics
Data are reported as mean ± standard deviation unless otherwise specified. Statistical analyses were performed using Prism 6.0 (GraphPad Software, Inc.). For behavioral task data analysis, multiple comparisons were performed using a one-way ANOVA within groups and the Tukey's post hoc correction for multiple comparisons. For immunohistochemical analysis, multiple comparisons were performed using a one-way ANOVA between groups with the Holm-Sidak post hoc correction for multiple comparisons. Reported p-values are adjusted for multiple comparisons.
Results
Short-term recovery of affected forepaw is enhanced with local BDNF delivery
The stroke lesion targeted the forelimb area of the primary motor cortex and thus, the Montoya staircase task was used to evaluate the performance of the injured forelimb in stroke-lesioned rats (Fig. 1). Only BDNF-treated animals showed a significant improvement in staircase performance at week 3 relative to the week 1 injured time point, demonstrating a beneficial effect. No significant improvements in performance were observed for the vehicle or injury groups in this task. Sham animals remained at baseline performance throughout the testing period.
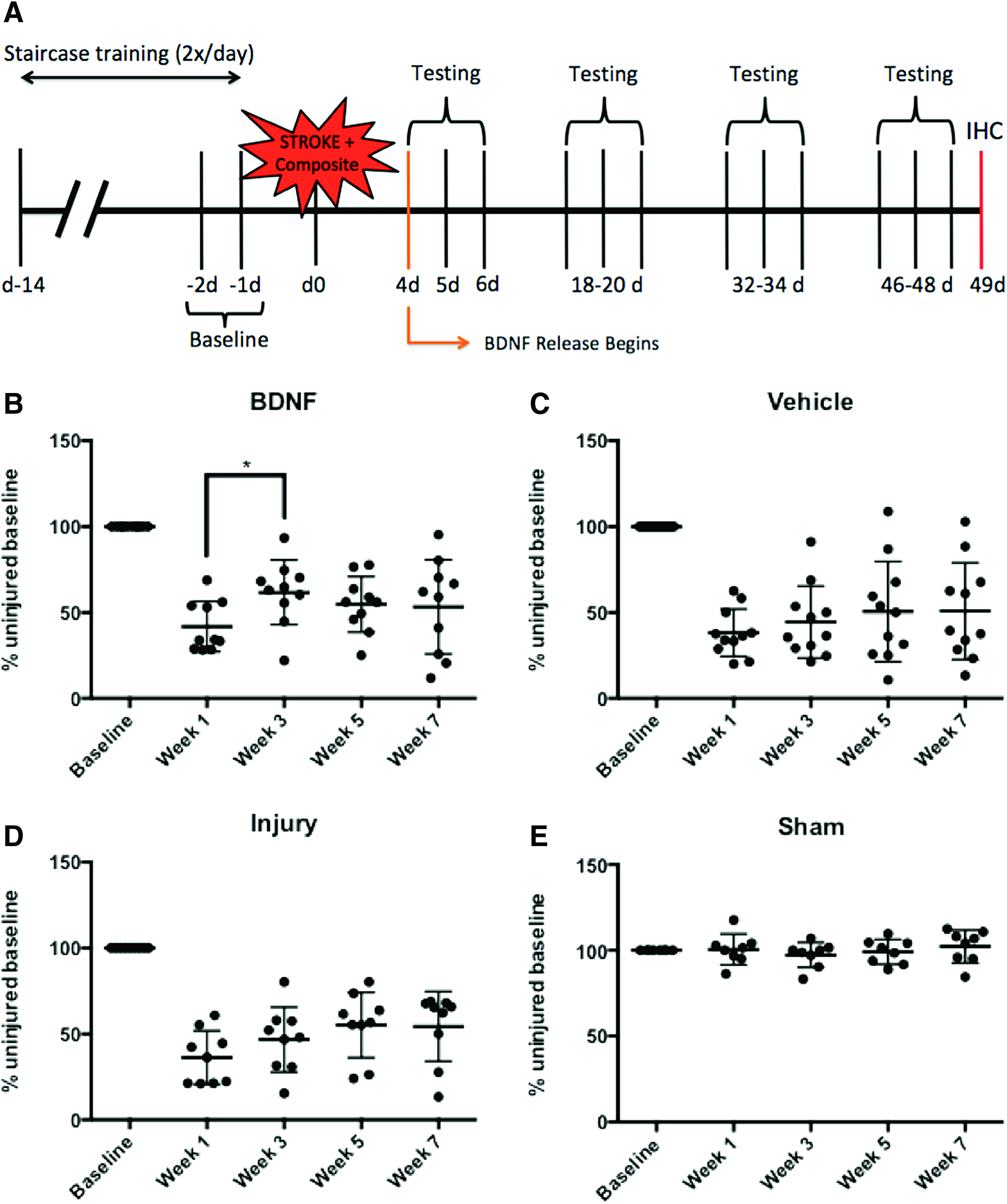
Local delivery of BDNF enhances early forepaw recovery following stroke injury. Data plotted as individual animal staircase task scores with mean ± SD shown, *p ≤ 0.05.
Recovery of affected hindlimb is enhanced with BDNF local delivery at 7 weeks
To assess hindlimb function, we used the tapered beam task (Fig. 2A). BDNF-treated animals exhibited recovery at both week 3 (p < 0.1, p = 0.0879) and week 7 (p < 0.05) relative to week 1 (Fig. 2B) in the affected hindlimb postinjury, suggesting that the effects of BDNF take some time to manifest in this task. Vehicle-treated animals exhibited a significant improvement in performance at week 3 relative to week 1, suggesting a transient benefit associated with HAMC, but this was not maintained to 7 weeks (Fig. 2C). Injury group animals did not show any significant improvement in performance at any testing point (Fig. 2D), and sham animals remained at their baseline level of performance (Fig. 2E).

Local delivery of BDNF and vehicle enhances hindlimb recovery following stroke injury. Data plotted as individual animal beam task scores with mean ± SD shown, #p < 0.1, *p < 0.05, **p < 0.01.
Stroke lesion volume is decreased by the presence of vehicle
NeuN staining was used to investigate the effect of local BDNF delivery on the stroke lesion volume. The lesion was defined as NeuN− and was traced using ImageJ software to quantify the area in each section (Fig. 3A). The total lesion volume (cortical and striatal combined) in BDNF and vehicle groups was significantly less compared with the injury group, indicating that vehicle itself was enough to reduce tissue loss. The cortical lesion (CL) volume in BDNF and vehicle groups was significantly decreased compared to the injury group, while the SL volumes were similar across all groups (Fig. 3B–D). As no significant difference was seen between the BDNF- and vehicle-treated groups, BDNF is unlikely to have contributed to this reduction of lesion volume.

Lesion volume, a proxy for tissue degeneration, is decreased in animals that received the vehicle with or without BDNF. Data plotted as individual mean ± SD, *p ≤ 0.05, **p ≤ 0.01.
Loss of mature neurons surrounding the lesion without vehicle
In addition to influencing plasticity, BDNF is known to have a prosurvival effect, acting mainly through the PI3K/Akt pathway.44,45 To determine whether our BDNF delivery strategy promoted neuronal survival in peri-infarct tissue, the number of NeuN+ cells was quantified in ROI (500 μm2) directly adjacent to the lesion (R1) and 500 μm away from the lesion (R2). Two regions for R1 and two regions for R2 were chosen on either side of the lesion and averaged together to give a representative sample of the tissue. Injury-only animals showed a significant loss of NeuN in R1, while the BDNF- and vehicle-treated animals showed no significant loss (Fig. 4).
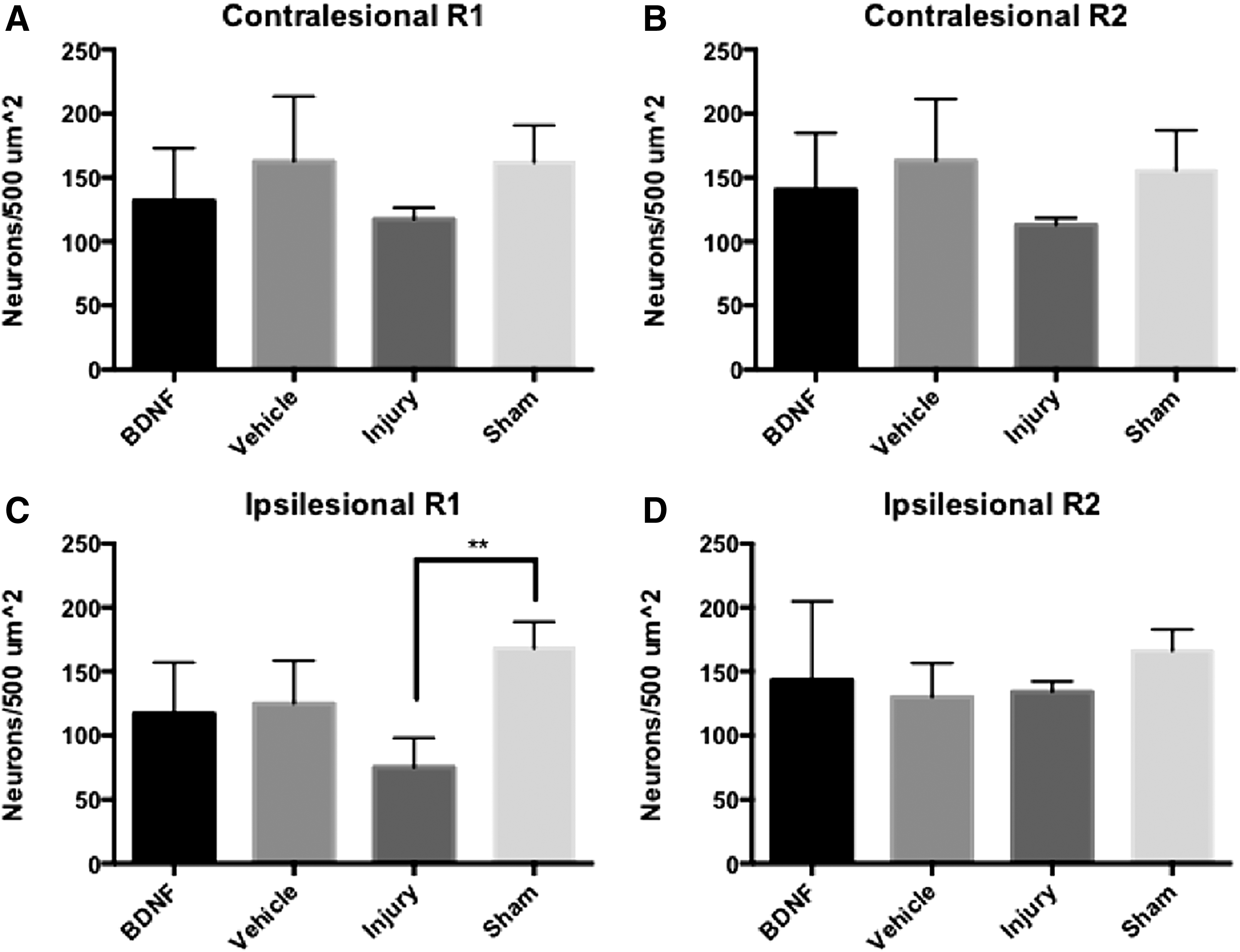
Injured animals experience loss of mature neurons in perilesional ROI. Data plotted as individual mean + SD, **p < 0.01.
Local delivery of BDNF increases synaptophysin expression in homotopic contralesional hemisphere
Given the reported role of BDNF as an agent of plasticity, we investigated synaptic plasticity with synaptophysin, a synaptic vesicle protein expressed presynaptically. 46 We observed significant increases in synaptophysin expression in the BDNF-treated animals in the contralesional hemisphere ROI R1 and R2 (Fig. 5). These increases were significant compared to vehicle and injury groups in region R1 and significant compared to vehicle, injury, and sham groups for region R2 (Fig. 5A, B). The ROI on either side of the lesion (in the ipsilesional hemisphere) did not display a significant upregulation of synaptophysin expression in any of the injured groups (Fig. 5C, D).

Local BDNF delivery results in significantly increased synaptophysin expression in the homotopic contralesional hemisphere. Data plotted as individual mean + SD, *p < 0.05, **p < 0.01.
Epicortical PLGA-HAMC composite allows sustained delivery of BDNF into the stroke-injured brain without encapsulation
To gain greater insight into the role of BDNF in repair, we characterized its tissue penetration in the brain at the behavioral testing time points (Fig. 6). At 1 day postdelivery, BDNF was not detectable in the tissue above endogenous levels at any depth. This is consistent with in vitro release profiles that exhibit a delay period before BDNF is released from the composite. 40 Diffusion into the tissue was first observed at 4 days postdelivery, with detectable concentrations throughout the collected depths. The highest concentration in the tissue occurred at 7 days postdelivery, and detectable concentrations were maintained out to 21 days. Thus, while the BDNF composite was applied at the time of injury, BDNF tissue diffusion, and hence treatment, was not immediate and allowed a statistically significant deficit to be created in the animals. When we delivered the BDNF composite 4 days postinjury to more closely simulate a clinical scenario, BDNF was detected in the tissue at 13 and 21 days postdelivery to a depth of 3 mm. Additional depth analysis past 3 mm revealed the presence of BDNF at 4.5 and 5 mm below the surface of the brain (Supplementary Fig. S2).
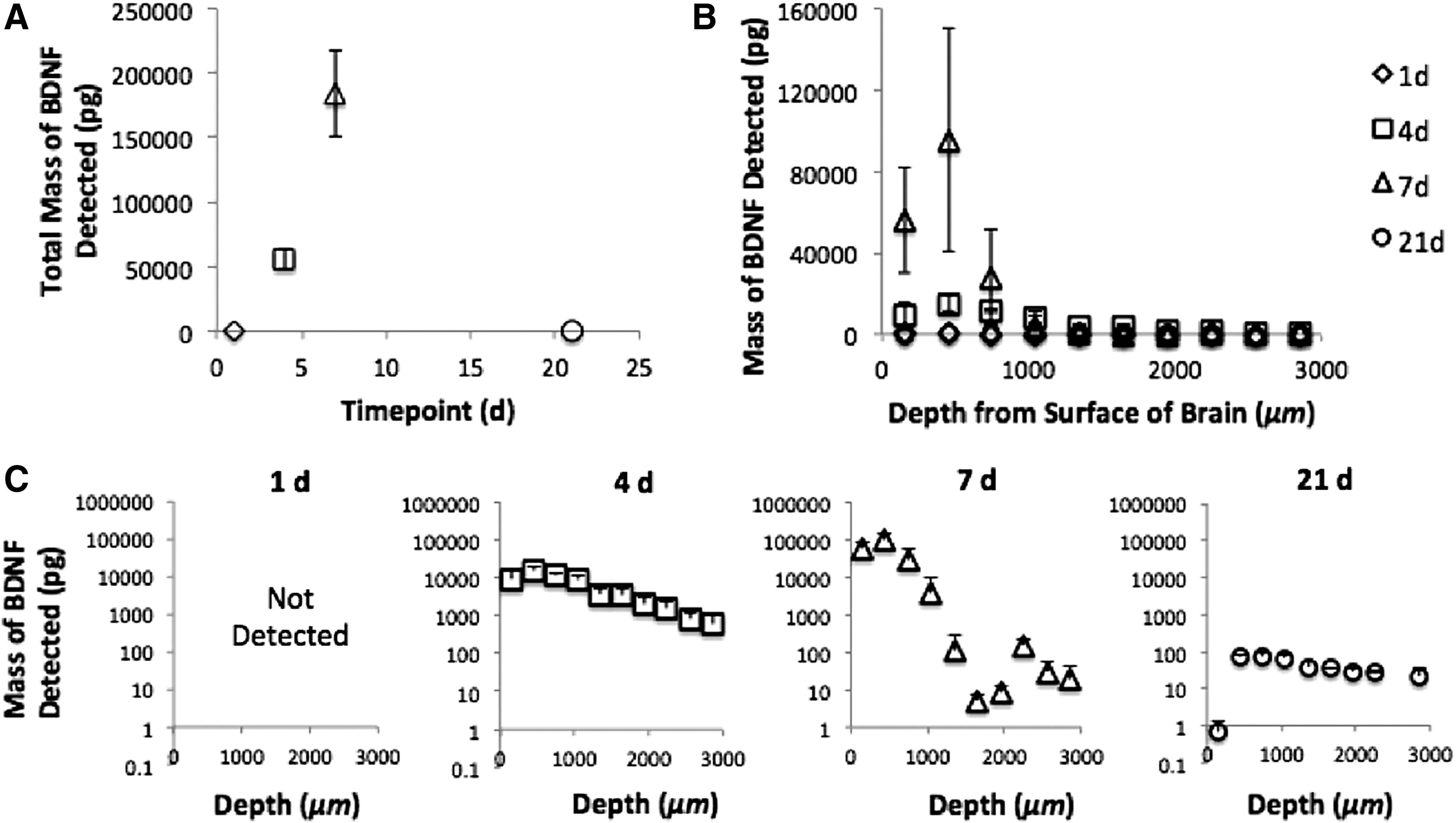
BDNF diffuses into stroke-injured rat brain tissue from the PLGA-HAMC composite deposited on the surface of the cortex, superficial to the lesion.
Discussion
In the first few hours following ischemia, the BDNF gene is upregulated in both peri-infarct and homotopic contralesional tissue, but this spike in expression quickly diminishes to baseline levels by 24 h postinjury. 12 We demonstrated that local, sustained delivery of exogenous BDNF to the stroke-injured rat brain has beneficial behavioral and tissue effects, and that this can be achieved using our novel delivery strategy, which is less invasive than current methods. As functional recovery is the overall goal of any stroke treatment strategy, behavioral testing was an integral part of our investigation. With our Et-1 stroke injury, we targeted the striatum and the primary motor cortex. The location and severity of injury are easily controlled with the Et-1 model, making it very reproducible and widely used in rodent studies of ischemia.13,35,42,47 Our cortical coordinates were largely specific to the forelimb, although these coordinates are also known to affect hindlimb function due to the close proximity of the areas, 42 and so we included both the staircase reaching task and the tapered beam tasks in our behavioral assessments.
We observed an early benefit of local BDNF delivery in the staircase task at week 3. This task requires a fine degree of motor control of the affected limb and since animals were not exposed to fine motor use or training scenarios outside of staircase task testing, it is not surprising that these circuits were not further enhanced throughout the course of the testing period. In contrast, performance in the tapered beam task reflected necessary daily movement. Relative to week 1, BDNF-treated animals recovered function at week 3 (with p < 0.10) and week 7 (with p < 0.05). Interestingly, vehicle-treated animals showed functional recovery at week 3 as well (with p < 0.01), yet this effect was not sustained. Immunohistochemical staining at the week 7 time point revealed tissue effects that gave additional insight into these behavioral outcomes.
In BDNF- and vehicle-treated animals, we observed preservation of tissue in the CL site and the absence of mature neuron loss in the perilesional tissue. These effects can be attributed to the epicortical placement of the composite, directly superficial to the CL, as high molecular weight HA is known to be anti-inflammatory 48 and our HAMC hydrogel has similar anti-inflammatory properties in the CNS. 39 These findings are consistent with previous studies that demonstrated the ability of the PLGA-HAMC composite to reduce lesion volume when administered epicortically above the lesion site in a mouse model of stroke. 39 By modulating the immune response, the HAMC hydrogel limits the extent of damage, thereby creating a more permissive environment for plasticity. This immune modulation may have allowed more endogenous plasticity to occur in the short term, resulting in the observed recovery of both BDNF- and vehicle-treated animals in the beam task at week 3; however, long-term effects observed at week 7 in the BDNF-treated animals indicate a benefit from the sustained presence of exogenous BDNF, possibly because this provided greater neuroprotection and encouraged synaptic plasticity in the surviving neurons.
Unexpectedly, we observed greater plasticity in the contralesional versus ipsilesional hemisphere as determined by synaptophysin staining. Increases in synaptophysin expression are associated with synaptogenesis and synaptic strengthening, and used to quantify synaptic plasticity.44,49 Consistent with others, we observed increased synaptophysin expression in BDNF-treated brains following stroke, although these differences were in the contralesional, and not the ipsilesional, hemisphere. Given that the glial scar contributes to the inhibitory environment, it may have prevented observable plasticity in ipsilesional tissue at 7 weeks postdelivery. The absence of glial scar in the contralesional hemisphere results in a more permissive environment for plasticity and this may explain why effects were only observed here.
Of the studies that have used synaptophysin as an indicator of synaptic plasticity in ischemic injury, few have investigated it in the contralesional hemisphere; however, the contralesional hemisphere is known to provide support in a compensatory manner, 50 and increases in the number of synapses and the volume of dendritic processes have been reported in layer V of the contralesional forelimb motor cortex following ischemia. 51 Reorganization of motor circuits takes time, even when rehabilitation therapy is used to promote cortical plasticity and recovery, 52 and thus, examination at different time points would reveal varying tissue effects. Studies in both humans53,54 and rats 55 that have investigated the role of contralesional activity in relation to functional outcome have found that contralesional activity is transiently increased in the short term, and that greater recovery requires a subsequent increase in the ipsilesional activity at chronic time points. Eventually, balanced hemispheric activity may be the key to optimal recovery. 56
The contralesional plasticity we observed in BDNF-treated animals is likely a product of multiple pathways, including synaptic plasticity of interhemispheric axonal projections, diffusion of exogenous BDNF across the corpus callosum, and/or redistribution of exogenous BDNF by CSF flow (Fig. 7).

Potential pathways of BDNF influence. Contralesional effects could be governed by interhemispheric axonal sprouting initiated by BDNF treatment in the ipsilesional hemisphere, anisotropic diffusion guidance of delivered BDNF along the corpus callosum, and/or redistribution of the delivered BDNF through CSF flow in the subarachnoid space. CSF, cerebrospinal fluid; NP, nanoparticles. Color images are available online.
The epicortical placement of the composite directly above the CL allows BDNF to reach the peri-infarct tissue, as demonstrated by tissue penetration data. As we were targeting plastic processes in the peri-infarct tissue, the observed tissue penetration was sufficient. Even without modification or additional diffusion facilitators, BDNF diffused to a depth of at least 3 mm from the epicortical composite. This diffusion depth comprises the cortical layer of tissue and includes the interface between cortex and corpus callosum. With BDNF diffusing to this depth in the tissue, it is possible that the anisotropic orientation of the white matter tracts of the corpus callosum facilitated protein diffusion to the contralateral hemisphere. 57 BDNF delivered to the ipsilesional hemisphere also likely initiated the MEK-ERK-CREB cascade in peri-infarct tissue, which upregulated endogenous BDNF expression in the ipsilesional hemisphere. The additional neuroprotection this provided would have contributed to the survival of perilesional neurons, while encouraging synaptogenesis and synaptic strengthening of interhemispheric axonal projections from these neurons. Cook et al. 35 demonstrated this effect in stroke-injured mice that received a BDNF-loaded hydrogel, injected into the stroke cavity tissue. They showed that BDNF-treated animals exhibited less loss of cortical interhemispheric projections and increased interhemispheric axonal sprouting within the corticostriatal systems. 35 In addition, BDNF diffusion to the contralesional hemisphere may have been facilitated by CSF flow in the subarachnoid space and the lateral ventricles. 57
Although the focus of our work has been stroke, the applicability of this delivery system for BDNF is evident for other neurological diseases and injuries. BDNF dysregulation has been implicated in many CNS pathologies, including Alzheimer's disease 7 and Parkinson's disease, 58 and effective delivery of this therapeutic without causing additional tissue damage could unlock new therapeutic options for these hard-to-treat conditions.
Conclusions
We validate a novel strategy for eliciting tissue repair and recovery of function following ischemic stroke in rats. Local delivery of unencapsulated BDNF with our PLGA-HAMC composite results in short-term recovery in forelimb function, as well as hindlimb recovery at week 7 testing. The PLGA-HAMC vehicle demonstrated beneficial effects when delivered at the time of injury, preserving mature neurons in the perilesional tissue and decreasing lesion volume, likely through the anti-inflammatory properties of the HA. At 7 weeks postdelivery, synaptic plasticity in the homotopic contralesional hemisphere was increased, as indicated by a substantial increase in synaptophysin expression. Our PLGA-HAMC formulation, loaded with unencapsulated BDNF, resulted in detectable concentrations of BDNF in the tissue to a depth of 3 mm below the surface for 21 days. We demonstrate the benefits of an epicortical strategy for the treatment of stroke and suggest its applicability to other neurodegenerative diseases.
Footnotes
Acknowledgments
We are grateful to the Natural Sciences and Engineering Research Council of Canada (NSERC Discovery to M.S.S., CGSD to J.M.O.) and the Canadian Institutes of Health Research (CIHR Foundation to M.S.S., CIHR Project to C.M.M. and M.S.S.) for funding our research. We are grateful to Matthew Jeffers and Dr. Dale Corbett (University of Ottawa) for thoughtful discussions and advice on the Et-1 stroke injury model in rats. We thank members of the Shoichet laboratory for thoughtful review of this article.
Disclosure Statement
The authors have no commercial associations or conflicts of interest to disclose; however, they acknowledge having composition of matter patents on HAMC.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.