
Abstract
Articular cartilage defects are common, causing significant pain and morbidities. To date, tissue engineering techniques using adipose-derived mesenchymal stem cells have offered a promising modality for cartilage repair. In this study, we investigated the effects of Eucomis autumnalis aqueous crude extracts on porcine adipose-derived mesenchymal stem cells (pADMSCs) to determine whether they can differentiate pADMSCs into chondrocytes. pADMSCs were prepared and treated with different concentrations of E. autumnalis aqueous crude extracts and transforming growth factor (TGFβ3) was used as a positive control in vitro. The cells were then stained with safranin O and toluidine blue to observe change in morphology. Micromass culture was used to assess chondrogenic differentiation. The deposition of glycosaminoglycan (GAG) chondroitin sulfate was assessed as well with dimethylmethylene blue dye assay. Microscopic observations showed that pADMSCs were adherent with fibroblastic morphology features and trypan blue staining showed that 96% of pADMSCs were viable. Micromass sections stained with alcian blue and safranin O showed blue and orange color deposition in between cells on the outer side of the sections, suggesting the presence of cartilage GAGs and collagens, respectively. We also noticed that E. autumnalis extracts at 5 and 15 μg/mL concentrations were able to induce chondrogenesis of pADMSCs, as observed in the morphology of the resultant cells. Treatment of cells with E. autumnalis extracts at all concentrations upregulated gene expression of SRY-type high-mobility group box 9 (SOX9) and downregulated gene expression of collagen type II and X. Increased GAG content was noticed on cells treated with E. autumnalis extracts at 5 μg/mL concentration compared to other treatment groups. These findings suggest that E. autumnalis crude extracts of at least 5 μg/mL concentration could possibly differentiate pADMSCs into chondrocyte-like cells. These results show the potential role that E. autumnalis could play in the differentiation factor.
Impact Statement
Eucomis autumnalis is one plant that is used by various traditional healers to alleviate the signs and symptoms associated with osteoarthritis. Although the exact mechanisms remain unknown, we hypothesized that this plant can induce chondrogenesis. In this work, we explored the potential for an aqueous crude extract from E. autumnalis to induce chondrogenesis in porcine adipose-derived mesenchymal stem cells (MSCs). The results reported in our article indicate that the aqueous crude extract from E. autumnalis was able to indeed induce chondrogenesis. Our research is relevant to communities that rely on plant-based remedies for their well-being.
Introduction
Mesenchymal stem cells (MSCs) offer an appropriate cellular model for studying articular cartilage repair. Since their discovery, many studies investigating the use of MSCs from different tissue sources have reported promising results in various models.1,2 Adipose-derived mesenchymal stem cells (ADMSCs) are considered a potential alternative source for cartilage repair.3,4 The use of ADMSCs for cartilage regeneration requires a multiparametric approach where biochemical and physiochemical factors, engineering and biomaterial, are holistically considered to improve or replace biological functions.3–6 In principle, the ADMSCs are first expanded in culture and differentiated into chondrocytes by a growth medium supplemented with transforming growth factor (TGFβ1, 3) or bone morphogenic protein-2 (BMP-2).7,8 These growth factors act on complex pathway systems to induce expression and regulation of cartilage matrix components such as aggrecan, glycosaminoglycans (GAGs), collagen type I, II, IX, and X, and SRY-type high-mobility group box 9 (SOX9).8,9
Although TGFβ1 and 3 and BMP-2 play a critical role during chondrogenesis, some studies have reported adverse effects associated with these growth factors during cartilage tissue repair.10–12 A study by Shlopov et al. showed that when a subpopulation of human articular chondrocytes was exposed to TGFβ, there was synthesis of MMP-13 (collagenase-3). 13 In synovial lining cells, TGFβ has also been shown to increase the synthesis of aggrecanases (ADAMTS4/5) and MMP-1, as well as the expression of proinflammatory cytokines, resulting in accelerated breakdown of the cartilage.10,11 Moreover, in a study conducted on ADMSCs, TGFβ was found to induce increased chondrogenic synthesis of collagen type X rather than collagen type II. 12
Given the wide range of growth factors and the confusion about their use in the maintenance of cartilage homoeostasis in vitro and in vivo, treatment of osteoarthritic joints remains a challenge. One potential solution would be to study medicinal plants that have apparently been used by traditional healers and herbalists to treat osteoarthritis (OA) or at least slow it down to a manageable degree. The coexistence of traditional and complementary medicine provides a fertile source for discovery of Western and African medicine, based on phytochemical leads. In South Africa, the Eucomis autumnalis plant is being used as herbal remedy for postoperative recovery. This genus has been proven to possess anti-inflammatory, antibacterial, and antihistaminic characteristics. 14 The factors above generated the necessity to explore the fundamental importance of using E. autumnalis aqueous crude extracts for porcine adipose-derived mesenchymal stem cell (pADMSC) viability and differentiation into chondrocytes (induce chondrogenesis). This approach is novel and could relieve burden of those suffering from early OA.
Materials and Methods
Plant procurement and preparation of crude aqueous extracts
E. autumnalis (Mill.) was harvested during summer (January to February) from the Eastern Cape Province, South Africa. The plant was identified at Schweickerdt Herbarium, University of Pretoria, and was given the voucher specimen number 123585. The bulb part of the E. autumnalis was collected and taken to the postharvest laboratory, where it was washed with water to remove soil and dust. After that, it was cut into small pieces and air dried at room temperature for 2 weeks. The dried material was ground in a Wiley mill grinder (Polymix PX-MFC 90D; LASEC, Cape Town, South Africa) and stored in the dark until extraction. Each ground sample was extracted separately by shaking the powder (30 g) for 6 h with 300 mL of distilled water (dH2O) on a Form Scientific orbital shaker (Labotech, Midrand, South Africa) at 200 rpm and filtered through Whatman No. 1 filter paper. 15 Water extracts were frozen at −70°C and lyophilized. Dried extracts were kept at −20°C until use.
Preparation of pADMSCs
Adipose tissue was harvested from the stifle (knee) joint of 3-month-old pig and digested with 0.1% collagenase type II (Life Technologies, Johannesburg, South Africa) at 37°C for 45 min. The resultant adipose cells were then used to generate ADMSCs, as previously described, with slight modifications. 16 In brief, the isolated cells were seeded at a density of 5 × 103 cells/mL in Dulbecco's modified Eagle's medium (Gibco, Life Technologies) supplemented with 10% fetal bovine serum (Life Technologies), 1% GlutaMAX-I (Gibco, Life Technologies), antibiotics (0.1% Gentamicin [Gibco, Life Technologies], and 0.1% Fungizone [Life Technologies]) in a T25 tissue culture flask and incubated at 37°C in a 5% CO2 atmosphere for 7 days. After the incubation period, the cells were observed under a microscope for spindle-shaped cells with fibroblast-like morphology, which were detached with trypsin (0.25% trypsin and 1 mM EDTA; Highveld Biological, Lyndhurst, South Africa) and prepared for in vitro assays.
Flow cytometric characterization
The porcine adipose tissue-derived cells were characterized for MSC qualities using flow cytometry according to the International Society of Cell Therapy. 17 After 7 days of culture, pADMSC viability was established before dilution to a cell density of 1 × 105 cells/mL. The cells were then incubated for 30 min at room temperature with CD90-FITC, CD44-PE, CD19-PE, and CD45-APC antibodies. All conjugated antibodies and markers were sourced from Abcam (Cambridge, United Kingdom) and supplied by BIOCOM Biotech (Pretoria, South Africa). The samples were analyzed using the MACSQuant Flow Cytometer Analyser (Miltinyi Biotech, Bergisch Gladbach, Germany).
Treatment of pADMSCs with E. autumnalis
The pADMSCs were treated with the E. autumnalis aqueous crude extracts at concentrations of 5, 15, and 30 μg/mL in a complete cell culture medium. TGFβ3 (Sigma-Aldrich, St. Louis, MO) at 10 ng/mL served as positive control, while the complete cell culture medium served as negative control.8,12
Morphological assessment of pADMSCs
pADMSCs were seeded in six-well plates at 1 × 105 cells/mL dilution and then treated with different concentrations of E. autumnalis aqueous extracts and TGFβ3 for 21 days. The expression of cartilage matrix markers was investigated by staining with 1% toluidine blue and safranin O (Cell Path Services, Randburg, South Africa). The medium was changed every 3 days, accompanied by supplementation with the concentrations of the experimental drugs until the treatment period was reached. Change in pADMSC morphology was examined by light microscopy using the Leica DM IL LED Fluo microscope (Leica Microsystems, Wetzlar, Germany).
Quantitative real-time polymerase chain reaction
After 21 days of treatment, ribonucleic acid (RNA) was isolated from pADMSCs at an initial density of 1 × 105 cells/mL using the NucleoSpin RNA kit (Macherey-NAGEL, Duren, Germany) according to the manufacturer's instructions, and then the genomic DNA was removed using the rDNase solution. The Qubit 2.0 fluorometer (Life Technologies) was used to quantify RNA (Qubit RNA BR Assay Kit; Life Technologies). Single-strand complementary DNA (cDNA) was synthesized using the iSCRIPT cDNA synthesis kit (Bio-Rad Laboratories, Richmond, CA). Real-time quantitative polymerase chain reaction (PCR) was performed in triplicate with the Step One plus sequence detector (Applied Biosystems, Foster City, CA) and TaqMan gene expression assays (Applied Biosystems) following the recommended protocols. Collagen type II and X and SOX9 messenger RNA (mRNA) levels were normalized to housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH) levels and expressed relative to the control (untreated) culture levels calculated using the threshold cycle (CT) method. 18 TaqMan gene expression assays Ss03373342_g1, Ss03392406_u1, Ss04323770_g1, and Ss03375435_u1 for collagen type II, SOX9, collagen type X, and GAPDH, respectively, were predesigned by Applied Biosystems and sourced through Life Technologies.
GAG assay
After 21 days of pADMSC (1 × 105 cells/mL) treatment, the cells were washed with phosphate-buffered saline and digested with 1 mL of 200 μg/mL proteinase K (Bioline, Taunton, WA) before overnight incubation at 37°C. Afterward, the samples were heated at 96°C for 10 min and diluted (1:10) with TBE buffer. Following sample dilution, 10 μL of each sample was transferred into a 96-well plate in quadruplicate, then 250 μL of dimethylmethylene blue (DMMB) dye solution was added, and the plate was read at 525 nm with SkanIt MultiScan go (ThermoFisher Scientific, Fairland, South Africa). A standard curve of chondroitin sulfate (Sigma-Aldrich) was drawn and referenced to calculate the amount of GAG secreted by the pADMSCs.
Micromass culture (chondrogenic differentiation)
The pADMSCs were prepared for pellet micromass culture in 15 mL Falcon tubes at dilution 5 × 105 cells/mL and differentiated in chondrogenic medium for 21 days. The medium was changed after every 3 days, accompanied by supplementation with the experimental drugs (control, E. autumnalis at concentration 5 μg/mL and TGFβ3 at 10 ng/mL). Pellets were harvested at the end of the incubation period and fixed with 10% buffered formalin overnight at room temperature. Thereafter, the pellets were processed, embedded in paraffin wax, and sectioned at 3 μm thickness. The sections where then stained with 1% alcian blue to evaluate the deposition of mucopolysaccharides (GAGs) staining blue and then counter stained with 1% neutral red stain and safranin O to evaluate cartilage collagen deposition staining orange to red. The staining reagents were purchased from Cell Path Services and the images were taken with a Nikon Eclipse E100-LED microscope with the Digital Sight DS-L3 camera system (Nikon, Centurion, South Africa).
Statistical analysis
All experiments (n = 6) consisted of biological triplicates and an average of the results was used. Statistical analysis was performed using Microsoft Excel software 2010 and the mean, standard deviation, standard error, and significance were calculated. The Student t-test was employed to determine the significance (where *p < 0.05 and **p < 0.01) of the difference in means between the control and experimental groups.
Results
Morphological and flow cytometric validation of the pADMSCs
Porcine adipose tissue-derived cells were assessed for features of MSCs using a microscope. Microscopic observations showed that the cells were adherent with fibroblastic morphology, presenting features known to MSCs (Fig. 1). The trypan blue staining showed that 96% of pADMSCs were viable.

Light microscopy examining the growth and morphological features of pADMSCs. Cultured cells showed a typical fibroblastic morphology of MSCs after 7 days of incubation. Magnification 10 × , scale bar represents 100 μm. MSCs, mesenchymal stem cells; pADMSCs, porcine adipose-derived mesenchymal stem cells.
Characterization of the cells was determined by flow cytometry, as shown in Figure 2. The results showed that the cells were indeed MSCs with minimal expression of hematopoietic cell surface antigen markers CD19 (2.27% ± 0.68) and CD45 (0.76% ± 0.31) and a marked increase in the expression of CD90 (95.4% ± 1.06) and CD44 (88.8% ± 0.79) (Fig. 2).

Analysis of the percentage expression of MSC surface markers CD19, CD44, CD45, and CD95 on the adipose-derived cells. The data are represented as the mean ± SD, n = 6. SD, standard deviation.
Morphological assessment of treated pADMSCs
In vitro monolayer cell culture was used to study the growth and morphological features of pADMSCs treated with crude aqueous extracts from E. autumnalis at concentrations of 5, 15, and 30 μg/mL for 21 days (Fig. 3). Safranin O staining of pADMSCs treated with TGFβ3 at 10 ng/mL and E. autumnalis aqueous crude extracts at 5 μg/mL revealed the presence of dark red arrears, reflecting high contents of collagen. Similarly, toluidine blue staining revealed the presence of blue-stained regions in cells treated with TGFβ3 at 10 ng/mL and E. autumnalis aqueous crude extracts at 5 μg/mL concentration, reflecting the presence of proteoglycan content. An observable decrease in cell volume and staining color intensity was noticed in cells treated with E. autumnalis at 15 and 30 μg/mL concentrations compared to other treatment groups (Fig. 3).

Safranin O and toluidine staining of pADMSC monolayer culture untreated
Cartilage markers gene expression analysis
The mRNA expression of SOX9 and collagen type II and X in pADMSCs treated with E. autumnalis aqueous crude extracts and TGFβ3 was assessed through real-time PCR after 21 days of treatment. Results showed that there was a dose-dependent decrease in the gene expression of SOX9 in pADMSCs treated with E. autumnalis aqueous crude extracts (Fig. 4A). When the pADMSCs were treated with 5 and 15 μg/mL of E. autumnalis aqueous crude extract, a 1.49 ± 0.04- and 1.22 ± 0.04-fold change in the relative gene expression of SOX9, respectively, was observed. The increase observed in the expression of the SOX9 gene in pADMSCs treated with 5 and 15 μg/mL of E. autumnalis aqueous crude extracts was significant (p < 0.05), relative to the untreated control. The results also showed that pADMSCs treated with 5 μg/mL of E. autumnalis aqueous crude extract had a 0.33-fold higher change in the gene expression of the SOX9 gene in comparison to those treated with TGFβ3 (Fig. 4A).
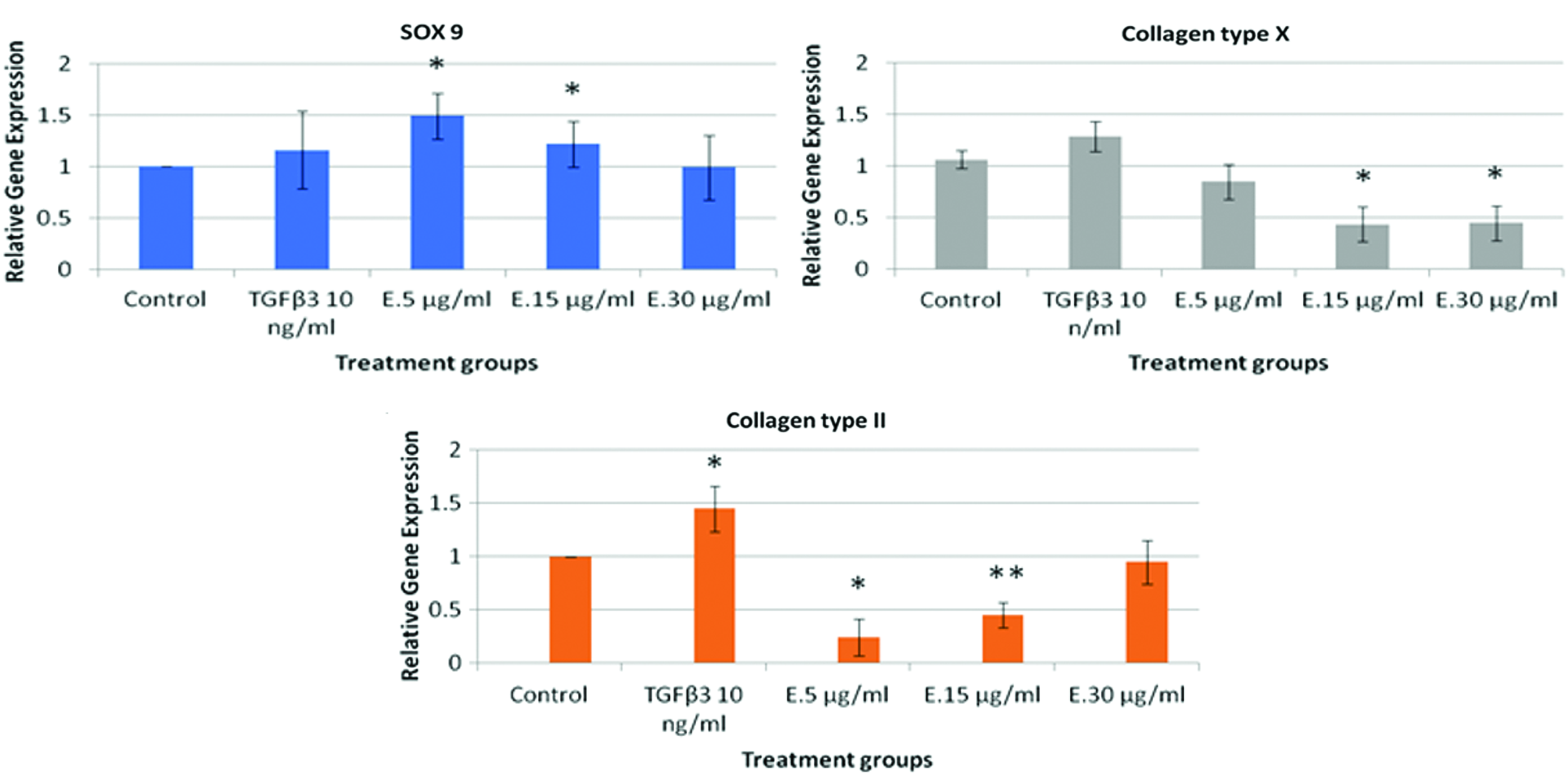
An effect of E. autumnalis aqueous crude extracts on the gene expression of cartilage differentiation markers. The pADMSCs were cultured for 21 days with or without (control) treatment groups. The difference in significance between the control and treatment groups was shown by * means p < 0.05 and ** means p < 0.01. E = E. autumnalis aqueous crude extracts. Color images are available online.
The expression of genes collagen type II and X was downregulated in pADMSCs treated with E. autumnalis aqueous extracts (Fig. 4B, C). The gene expression of collagen type X was significantly (p < 0.05) downregulated in pADMSCs treated with 15 and 30 μg/mL of E. autumnalis aqueous crude extract in comparison to the untreated sample (Fig. 4C). The relative gene expression of collagen type II in pADMSCs treated with 5 and 15 μg/mL of E. autumnalis aqueous crude extracts was 0.24 ± 0.03 and 0.45 ± 0.01, respectively. The difference in relative gene expression in pADMSCs treated with 5 and 15 μg/mL of E. autumnalis aqueous crude extracts was significantly (p < 0.05) lower in comparison to that observed in the untreated cells (Fig. 4B).
Determination of the amount of GAG-chondroitin sulfate
A DMMB assay was done to quantitate the amount of GAG-chondroitin sulfate content produced by pADMSCs treated with E. autumnalis aqueous crude extracts after 21 days. Results showed that pADMSCs stimulated with 5 μg/mL of E. autumnalis aqueous crude extracts and TGFβ3 produced 25.5 ± 0.02 and 25.3 ± 0.02 μg/mL of GAG-chondroitin sulfate, respectively (Table 1). There was relatively similar production of GAG content between treatment groups. The untreated control group had the lowest GAG content compared to the treatment groups TGFβ3 and E. autumnalis aqueous extract at 5 μg/mL concentration. pADMSCs treated with E. autumnalis aqueous extracts at 5 μg/mL concentration had the highest GAG content when compared to all treatment groups, as shown in Table 1.
Measurement of Glycosaminoglycan Content Secreted by Porcine Adipose-Derived Mesenchymal Stem Cells After Treatment with Eucomis autumnalis Aqueous Extracts and Transforming Growth Factor β3 on Day 21
The difference in significance compared between treated groups and the untreated control shown by a means p < 0.05.
GAG, glycosaminoglycan; TGF, transforming growth factor.
Micromass culture determining chondrogenic differentiation
The presence of GAGs and collagens synthesized in the cartilage matrix was evaluated by staining pADMSC micromass culture pellet sections after 21 days of treatment with E. autumnalis at 5 μg/mL and TGFβ3 at 10 ng/mL (Fig. 5). Histological staining of the sections of pADMSC cell mass using alcian blue staining demonstrated the presence of GAGs as indicated by the blue color (Fig. 5A). Results showed that the micromass treated with TGFβ3 had a wider blue-stained zone in comparison to that treated with the E. autumnalis extracts (Fig. 5A). Nonetheless, the micromass treated with E. autumnalis had a more intense blue stain, mostly on the outer section and between cells, in comparison to the negative control (Fig. 5A). Safranin O staining of the micromass sections was positive for collagens, as indicated by the orange color on the outer side of the pellet section for both treatment groups, except the negative control (Fig. 5B).

Discussion
MSCs have been reported to provide an alternative to conventional procedures of therapy. One of the common diseases in which MSC-based therapies may hold great promise is OA.2,19 The use of MSCs in cartilage regeneration relates to applications that involve growth factors, tissue engineering, and regenerative medicine. The growth factors act on complex pathway systems to induce cell proliferation, expression, and regulation of cartilage matrix components during chondrogenic differentiation.8,19 To realize the therapeutic potential of MSCs for cartilage regeneration, this study explored the fundamental importance of using E. autumnalis aqueous crude extracts for pADMSC viability and differentiation into chondrocytes (induce chondrogenesis).
This study has shown that the porcine isolated adipose tissue cells were indeed MSCs with minimal expression of hematopoietic cell surface antigen (Fig. 2). The pADMSCs had a surface attachment capacity with viable and proliferating cells, as characterized by the International Society of Cell Therapy.17,20 Evaluation of chondrogenic differentiation by staining methods revealed the presence of cartilage collagen and proteoglycan on monolayer culture by staining with safranin O and toluidine blue, respectively (Fig. 3). This finding suggests that E. autumnalis aqueous crude extracts at 5 μg/mL concentration were able to differentiate pADMSCs into chondrocytes. The results were in accordance with some other studies that showed that MSCs have chondrogenic potential through staining methods.21,22
The cell culture method employed in this study exhibited the required MSC qualities, namely the ability to self-renew and differentiate into cartilage tissue, the expression of surface markers CD90 and CD44, and the lack of expression of CD19 and CD45.7,20 Based on these observations, the expression of the cartilage-specific markers SOX9 and collagen type II and X was evaluated by quantitative gene expression (Fig. 4). TGFβ3 has been reported to be the most important inducer of chondrogenesis by acting on SOX9, which in turn regulates the expression of cartilage proteins.8,9,19 Our study showed that E. autumnalis aqueous extracts at a concentration of 5 μg/mL significantly expressed SOX9 compared to the untreated and positive controls, as shown in Figure 4A. Interestingly, E. autumnalis aqueous extracts at all concentrations downregulated the expression levels of collagen type II mRNA (Fig. 4B), an essential component of articular cartilage. Previous reports showed that expression of collagen type II is an early marker of MSC chondrogenic differentiation. 12 Thus, the reduction in the expression of collagen type II might not contribute to cartilage regeneration and deserves further research. A follow-up study showed that expressions of collagen type II mRNA levels were upregulated after 14 days of pADMSC treatment with E. autumnalis (unpublished observation). We, furthermore, showed that there was less expression of collagen type X mRNA levels (a hypertrophic marker during MSC differentiation) on cells treated with E. autumnalis aqueous crude extracts at all concentrations versus TGFβ3 (Fig. 4C). Monolayer data show a reduction in cell number at high doses of E. autumnalis aqueous extracts; this might reason for a dual decrease in the expression of collagen type II and X mRNA. This study once again proved the literature findings that TGFβ3 can induce terminal differentiation of pADMSCs with transient cartilage properties.12,19 During assessment of GAG secretion by pADMSCs treated with E. autumnalis aqueous crude extracts and TGFβ3, we noticed that the cells treated with plant extracts at a concentration of 5 μg/mL had the highest content of chondroitin sulfate, as shown in Table 1. At higher concentrations of the plant extract, the amount of chondroitin sulfate is lower compared with untreated pADMSCs. These data, supported by Figure 3, further suggest that E. autumnalis aqueous extracts at higher concentrations are less effective. There are different types of GAGs of specific physiological significance. In cartilage, chondroitin sulfate is usually found attached to proteins as part of a proteoglycan and it is mostly important, as it provides a structural component to the cartilage and is resistant to compression. 23 Maintaining micromass pellets in culture was difficult in this study. The cell aggregates were delicate and technically difficult to feed. However, we managed to prove that our plant extract induced pADMSCs to form cartilage-like matrix, embedded with chondrocyte-like cells. Interestingly, cell pellets treated with the positive control seemed to have a compact matrix compared to E. autumnalis-treated and untreated control groups, observed in Figure 5. This finding was in correlation with previous studies on evaluating chondrogenesis on micromass culture.12,24 From a clinical standpoint, further investigation is needed on optimization of scaffold-free chondrogenesis protocols to generate a stable tissue for clinical use. These findings are crucial to the study and prove the hypothesis that the medicinal plant E. autumnalis indeed has the capacity to influence pADMSC chondrogenesis and increase cell proliferation, characteristics known to be related to the growth factors TGFβs and BMPs.8,25
Conclusions
The key findings in our study were that E. autumnalis aqueous crude extract of at least 5 μg/mL concentration (1) improves ADMSC attachment, (2) induces expression of cartilage-specific marker SOX9 and decreases expression of hypertrophic marker collagen type X, and (3) differentiates ADMSCs into cartilage-like matrix. These effects are shown in Figures 3–5, respectively. More experimentation is required to evaluate the effects of the plant extract mechanism of action and its potential effects on animal models (in vivo). In addition, we could isolate the pure active morphogenic compounds from the plant extract responsible for the chondrogenic effects. The results from the proposed future studies, added to this study, may lead to significant improvements in MSC-based therapies for cartilage defect regeneration. Our study presents better understanding of the potential effect E. autumnalis could play as a differentiation factor during chondrogenesis. However, the question of standardization of the extraction methods as a prerequisite for obtaining comparable results must be addressed in future for a broad application of the extracts.
Footnotes
Acknowledgments
This work was financially supported by the National Research Foundation and Technology Innovation Agency Grants. The funders have no role in the study design, data collection and analysis, or the decision to publish the article. We thank Vleis 1 abattoir for providing porcine knees and our colleagues for technical and review support.
Disclosure Statement
The authors have no conflict of interests relevant to this article and no competing financial interests exist.