
Abstract
Vascularization of large bone grafts is one of the main challenges that limit the clinical application of bone tissue engineering (BTE). In this way, cell cocultures, which involve the cross-talk between endothelial and osteogenic cells, have shown to be an effective strategy for in vitro prevascularization. Dental pulp represents an easily accessible autologous source of adult stem cells. A subset of these cells, named dental pulp pluripotent-like stem cells (DPPSCs), shows high plasticity and great capacity to differentiate into different tissues. Here, we suggested a combination of bone-like DPPSC and endothelial-like DPPSC to induce vascularized bone formation from a unique stem cell population. In addition, we evaluated the use of inorganic ions dissolved from S53P4 bioactive glass (BaG) in different medium compositions. Results show that endothelial medium with BaG extracts provides an effective way to enhance both endothelial and osteogenic processes, supporting the formation of vascular-like structures and mineralization simultaneously. Furthermore, 3D DPPSC cocultures in the same medium conditions demonstrated the formation of vessel-like structures that appear to be functional as indicated by the presence of an internal lumen. Overall, these results would provide a new promising system for the prevascularization of BTE constructs.
Impact Statement
In this study, we proposed for the first time the use of inorganic ions dissolved from BaG in a cell coculture system to induce vascularized bone formation in vitro. For that, we used dental pulp pluripotent-like stem cells from a single individual source obtained in a minimally invasive extraction manner. Moreover, we carried out all the experiments under xeno-free conditions, allowing the extrapolation of the results to the development of clinically orientated applications. Overall, these results would provide a new promising system to promote the success and survival of bone tissue engineering constructs after implantation.
Introduction
One of the critical problems of bone tissue engineering (BTE) is the development of a rapid vascularization after implantation to provide nutrients and oxygen to the osteoblast cells, as well as to remove CO2 and waste products. In the case of bone tissue, the vasculature also supplies the calcium and phosphate needed for mineralization.1,2 Hence, if the tissue-engineered construct does not contain or allow a rapid ingrowth of blood vessels, it would lead to hypoxia and death of the implanted cells.
It has been demonstrated that capillary-like structures created in vitro show a structure similar to capillaries in vivo3,4 that will anastomose rapidly and supply blood to the construct. Different osteoblasts and endothelial cells coculture systems have been studied to offer conditions for the endothelial cells to migrate and form a network of capillary-like structures through the osteoblasts. This movement and proliferation of endothelial cells to form capillary-like structures is known as angiogenesis, and is dependent on a variety of proangiogenic factors. 1 Moreover, it has been demonstrated that angiogenesis is able to regulate the recruitment of stem cells and their orientation to the osteoblastic lineage.1,5
One of the critical factors for the efficacy of these cocultures is the choice of the cell type. Currently, there are different studies that combine osteogenic and endothelial cells from different sources. Primary osteoblasts or human osteoblasts cell lines are commonly used as osteogenic cells. 6 Human umbilical vein endothelial cells (HUVECs), human dermal microvascular endothelial cells, or outgrowth endothelial cells are used as endothelial cells.7,8 Although primary osteoblast and endothelial cells can be isolated and cultivated relatively easily, it is difficult to obtain sufficient number of cells before they begin to change, loose specific phenotypes, and undergo dedifferentiation. 9 An optimal approach for clinical translation of coculture constructs would be the obtainment of both endothelial and osteoblastic cells from a single individual source in a minimally invasive extraction manner. Nonetheless, there are few reports on cocultures of cells originated from a single stem cell population.10,11 Mesenchymal stem cells (MSCs) are easily differentiated into bone-like cells; however, microcapillary-like structures are not frequently observed.10,11 On the contrary, it is known that pluripotent stem cells (PSCs), such as embryonic stem cells and induced PSCs, present high differentiation potential into both osteogenic and endothelial-like cells, forming microcapillary-like structures in vitro as well as neovascularization in vivo.12,13 PSCs show unlimited passage potential, mature phenotypes, and can be reimplanted into the same donor. However, they commonly show ethical problems or safety risks and, up to date, cocultures from PSCs targeting BTE approaches have not yet been reported.
A possible alternative could be the use of a new adult stem cell population from the dental pulp of human third molars with pluripotent-like properties. These cells, named dental pulp pluripotent-like stem cells (DPPSCs), express pluripotency markers until late passages, show chromosomal stability, and have demonstrated the ability to differentiate into cells from the three embryonic layers. 14 DPPSCs have higher capacity to differentiate into osteogenic and endothelial tissue than other dental pulp stem cells, and they are able to grow, attach, and differentiate onto different biomaterials.15–17 Therefore, DPPSC might be a promising candidate for coculture studies in BTE.
Originally developed for bone regeneration, bioactive glasses (BaGs) are currently investigated for BTE applications. BaGs belong to a group of surface reactive amorphous biomaterials, which are able to stimulate the osteogenic differentiation of stem cells displaying good biocompatibility and biodegradation properties. 18 In addition, it has demonstrated angiogenic effects, such as increased secretion of vascular endothelial growth factors (VEGFs) in fibroblasts, endothelial cells proliferation, and tubules formation. 19 S53P4 glass, commercially available as BonAlive®, is able to induce osteogenic differentiation of human MSCs cultured in direct contact with BaG surface as well as in indirect contact, suggesting that the ions from this BaG alone are capable of inducing osteogenic differentiation. 20 S53P4 BaG can also induce increased vascularization when incorporated in tissue-engineered scaffolds. 19
Here, we propose to study the combination of bone-like and endothelial-like DPPSCs under the effect of different medium compositions conditioned with inorganic ions dissolved from S53P4 BaG. This is the first study in which BaG ions are used to enhance both osteogenic and angiogenic characteristics simultaneously in a coculture system. In addition, we carried out all the experiments under xeno-free conditions, with human serum (HS) in the used media, which represents the natural growth environment of human-originated cells and allows the extrapolation of the results to the development of clinically orientated applications in BTE.
Materials and Methods
Isolation, culture, and differentiation of DPPSC
Third molars of healthy patients were extracted for orthodontic reasons and obtained with informed consent from donors. All experiments were performed in accordance with the guidelines on human stem cell research issued by the Committee on Bioethics of the Universitat Internacional de Catalunya (study code BIO-ELB-2013-03).
DPPSCs were isolated and cultured as previously described.14–17 In brief, dental pulps were disaggregated with collagenase type I (3 mg/mL; Sigma) for 60 min at 37°C in continuous shaking. After washing with DPBS, the extracts were cultured in a specific medium composed of 60% Dulbecco's Modified Eagle's medium (DMEM) -low glucose (Life Technologies), 40% MCDB-201 (Sigma), 1 × SITE (Sigma), 10−4 M ascorbic acid 2-phosphate (Sigma), 1% penicillin/streptomycin (Life Technologies), 1% HS (BioWest), 10 ng/mL hPDGF-BB (R&D), and 10 ng/mL EGF (R&D) in flasks precoated with 100 ng/mL fibronectin and incubated overnight at 37°C in a 5% CO2 incubator. DPPSCs were detached at 30% confluence by adding 0.25% TrypLE Express Enzyme and repleated at 100–200 cells/cm 2 .
For osteogenic differentiation, DPPSCs were cultured in RPMI 1640 Glutamax (Life Technologies) supplemented with 5% HS (BioWest), 10 mM β-glycerol phosphate (Sigma), 50 μM L-ascorbic acid (Sigma), 0.01 μM Dexamethasone (Sigma), and 1% penicillin/streptomycin (Life Technologies) at 4 × 103 cells/cm2 in 24 well plates for 15 days. For endothelial differentiation, DPPSCs were cultured in EGM-2 bullet kit (Lonza) at 2 × 104 cells/cm2 in 24 well plates for 15 days. The 2% FBS of the EGM-2 bullet kit was replaced for 2% HS (Biowest). The medium was changed every 2 days in both differentiations.
Culture of HUVEC
HUVECs (Sigma-Aldrich) were maintained with EGM-2 BulletKit (Lonza) and cultured according to the manufacturer's protocol.
Cell viability and proliferation
Cell viability was evaluated qualitatively using Live/Dead staining probes (Live Technologies) at days 7 and 15 as previously described. 20 Images of the viable cells (green fluorescence) and dead cells (red fluorescence) were obtained using an Olympus IX51 phase contrast microscope with fluorescence optics. Cell proliferation was analyzed by determining the amount of total DNA with CyQUANT Cell Proliferation Assay Kit (Life Technologies) also at 7 and 15 days of culture according to the manufacturer's protocol. Fluorescence was measured at 480/520 nm with a microplate reader (Victor 1420 Multilabel Counter).
Immunofluorescences
For immunofluorescence (IF), cells were fixed with 4% PFA (Sigma) supplemented with 0.2% Triton-X 100 and blocked with 1% BSA for 1 h at 4°C. After PBS washes, cells were incubated with the primary antibodies against COL1 (Abcam; 1:100), OC (Abcam; 1:100), CD31 (Dako; 1:40), or vWF (Abcam; 1:500) diluted in 1% BSA overnight at 4°C. After washes, the corresponding secondary antibodies Alexa-fluor 488 or 568 (Abcam, 1:800) were added for 1 h at RT. Nuclei were stained with DAPI (1:1000). For 3D cocultures, cells were fixed with 4% PFA, washed with PBS, and the actin cytoskeleton was stained with phalloidin (Abcam; 1:1000) overnight at 4°C. Nuclei were incubated with DAPI (5:1000) for 1 h at RT. Images were obtained using an Olympus IX51 phase contrast microscope with fluorescence optics.
qRT-PCR analyses
The relative expression of endothelial and osteogenic marker genes was studied by quantitative reverse transcription polymerase chain reaction (qRT-PCR). The total messenger RNA was isolated using NucleoSpin RNA II kit (Macherey–Nagel) after which the isolated mRNA was reverse transcribed to cDNA with the High-Capacity cDNA Reverse Transcriptase Kit (Applied Biosystems). Data were normalized to the expression of housekeeping gene RPLP0. The sequences of the primers used are listed in Supplementary Table S1. The mixture contained 50 ng cDNA, 300 nM forward and reverse primers, and Power SYBR Green PCR Master Mix (Applied Biosystems). The amplifications were done in a CFX96 Real-Time PCR Detection System (Bio-Rad).
Alizarin Red S staining and quantification
Mineralization was determined by Alizarin Red S staining, which stains the calcium minerals in red. Cells were fixed with 70% ethanol for 1 h (−20°C) and stained with 2% Alizarin Red S solution (Sigma) for 10 min at RT. To quantify the results, the dye was extracted with 100 mM cetylpyridinium chloride (Sigma) during 3 h at RT, and the absorbance was measured at 544 nm (Victor 1420 Multilabel Counter).
BaG granules manufacture
BaG S53P4 granules were supplied by Åbo Akademi University, Turku, Finland. In brief, BaG S53P4 were prepared from batches of analytical grade reagents Na2CO3, K2CO3, CaCO3, MgO, CaHPO4·2H2O (Sigma), and Belgian glass quality glass sand. The batches yielding 300 g glass were melted in a platinum crucible for 3 h at 1360°C, cast, annealed, crushed, and remelted to ensure homogeneity. The oxide composition of the S53P4 is described and compared with the original Bioglass® 45S5 composition shown in Supplementary Table S2. Annealed glass blocks were crushed and sieved to yield granules of 500–1000 μm. The crushing was done according to the ISO 719 procedure without milling. The granules were washed with acetone in an ultrasound batch at least five times to minimize the fine grained particles attached to their surface. Finally, acetone was evaporated, and the particles were dried at 120°C.
BaG extracts preparation
The S53P4 BaG granules used in the extract preparations were first disinfected with 70% ethanol and allowed to dry at room temperature for 2 h. To leach ions from the BaG granules, 87.5 mg/mL granules were incubated for 24 h at 37°C in cell culture dishes with the corresponding extraction medium. The osteogenic and basal extraction medium contained RPMI 1640 Medium GlutaMAX™ and 1% penicillin/streptomycin (Life Technologies). The endothelial extraction medium contained EBM-2 supplemented with 0.1% Gentamicin/Amphotericin-B (Lonza). After incubation, the extracts were sterile filtered, and HS (Biowest) was added to the extract medium (5% in RPMI and 2% in EBM-2). The medium composition of RPMI extract medium and HS is referred to as basal medium extract (BM extract). To obtain osteogenic medium extract (OM extract), BM extract was supplemented with osteogenic factors (10 mM β-glycerol phosphate, 50 μM L-ascorbic acid, 0.01 μM Dexamethasone; Sigma). To obtain endothelial medium extracts (EM extract), the EBM-2 extracts with HS were supplemented with the endothelial factors of the EGM-2 Bulletkit (hEGF, VEGF, R3-IGF-1, ascorbic acid, hydrocortisone, hFGF-β, and heparin) to the manufacturer's concentrations. A schematic representation of the BaG extract preparation is shown in Supplementary Figure S1.
Two-dimensional cocultures
For bone-like DPPSC–HUVEC cocultures, DPPSCs were cultured in OM during 2 weeks to induce bone-like DPPSCs. Then, bone-like DPPSCs were seeded onto 24-well culture plates coated with fibronectin (Millipore) at a density of 3 × 105 cells/well and cultivated in OM. After 24 h, HUVECs were added at a density of 2 × 105. Cocultures were performed at a cell ratio of 3:2 (bone-like DPPSC:HUVEC) under different medium compositions.
For bone-like DPPSC-endothelial-like DPPSC cocultures, DPPSCs were cultured in OM and in EM during 2 weeks to induce bone-like DPPSC and endothelial-like DPPSC. Then, DPPSC cocultures were performed at a cell ratio of 3:2 (bone-like DPPSC:endothelial-like DPPSC). For that, bone-like DPPSCs were seeded onto 24-well culture plates coated with fibronectin (Millipore) at a density of 3 × 105 cells/well in OM. After 24 h, endothelial-like DPPSCs were added at a density of 2 × 105 cells/well using different xeno-free media compositions (OM; EM; a mixture of osteogenic and endothelial medium, OM/EM; basal medium, BM) supplemented with or without S53P4 BaG extracts (87.5 mg/mL; 0.08% w/v). Bone-like and endothelial-like DPPSCs were also cultivated as monocultures at the same density of cocultures. The medium was changed every 2 days. A diagram of the experimental design is provided in Figure 1.
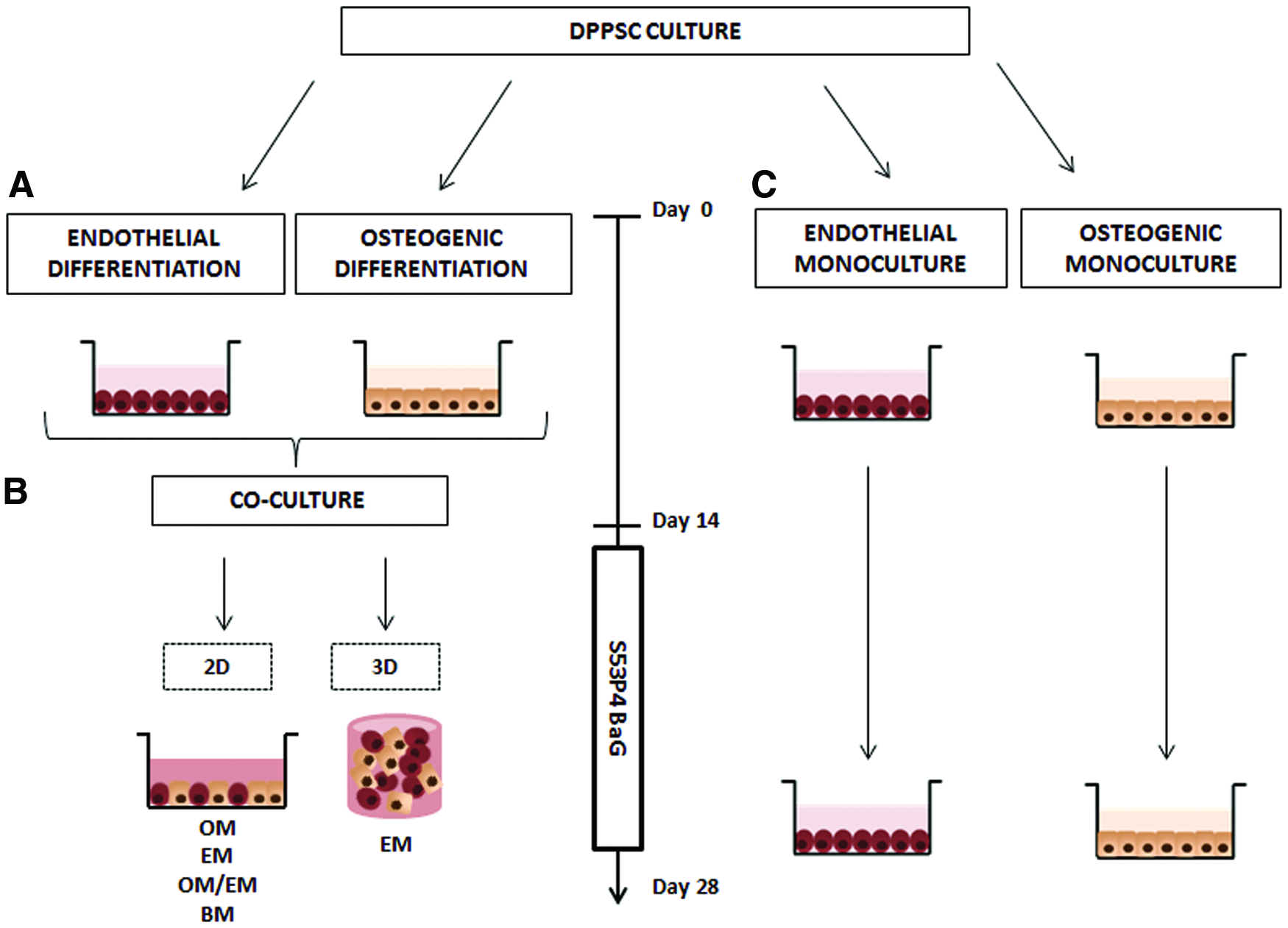
Experimental design of coculture systems used for bone-like tissue prevascularization.
Three-dimensional cocultures
Endothelial and osteogenic-like DPPSCs (three bone-like: two endothelial-like DPPSC) were encapsulated in 0.5% of PuraMatrix (Corning) and cultured over 14 days in EM conditioned with BaG and EM control media. In brief, the viscosity of PuraMatrix stock solution (1% w/v) was reduced by vortexing and centrifuging. The cells were centrifuged at 1 × 106 cells/mL and resuspended in 10% sucrose at twice the final desired concentration. Equal volumes of PuraMatrix and 2 × cell/sucrose mixture were mixed, and 250 μL of the mixture was added to the center of the well (24 well plates). Gelation was initiated by running down the side of the well on top of the hydrogel. The medium was changed twice over the next hour to equilibrate the pH and every 2 days of culture.
VEGF quantification
The supernatants of cocultures and endothelial monocultures were collected and stored at −20°C. VEGF secretion was quantified by an enzyme-linked immunosorbent assay using the human VEGF Elisa Kit (Sigma) according to the manufacturer's protocol. The absorbance was measured at 450 nm (Victor 1420 Multilabel Counter).
Statistical analysis
Statistical analyses were performed with SPSS Statistics version 21.0 (IBM). The results were analyzed by applying the two-way analysis of variance (ANOVA) test for multiple factors. Confidence intervals were fixed at 95% (p < 0.05). GraphPad Prism was used to graph all the quantitative data presented as mean and standard deviation (SD). See figure legends for specific information regarding the number of technical (n) and biological replicates (N).
Results
BaG extracts maintain the viability of DPPSCs and enhance their differentiation toward osteogenic and endothelial lineage
Effect of BaG extracts in DPPPSC osteogenic differentiation
The effect of the BaG-conditioned medium on DPPSC was evaluated at 7 and 15 days of osteogenic induction. In the presence of OM-BaG, no dead cells were observed, and the morphology also appeared similar to standard OM conditions at day 7. However, there was a significant decrease of proliferation in OM-BaG cultures (Fig. 2A). By qRT-PCR, cultures in OM-BaG upregulated the expression of the early osteogenic markers OSX and DLX5 at day 7 and the expression of the later marker OC at day 15 (Fig. 3A). Moreover, IF stainings revealed that, in both culture conditions at day 7, COL1 was still predominantly located intracellularly and at day 15, it was secreted to the ECM (Fig. 3B). Nevertheless, the OM-BaG seemed to induce higher production of OC at day 7 (Fig. 3C).

Viability and proliferation of DPPSCs cultured in BaG extract media.

Effect of BaG extracts in the osteogenic differentiation of DPPSC.
Moreover, Alizarin Red S staining showed that OM-BaG induced extensive mineral formation compared with the standard OM. The difference was significant as seen after the quantification of the staining (Fig. 3D). In addition, some control experiments comparing the effect of BaG ions in DPPSC under BM and OM conditions were performed to determine whether the cellular response to the presence of BaG ions depends on the medium conditions and if BaG ions have some effect without differentiation supplements. Results show that in BM culture conditions the effect of BaG ions is negligible, while under OM the cells respond to BaG ions with an increase of osteogenesis. At day 15, only under OM culture conditions treated with BaG ions collagen was secreted to the ECM, and larger amounts of OC were observed (Supplementary Fig. S2).
Effect of BaG extracts in DPPPSC endothelial differentiation
The endothelial differentiation potential of DPPSC cultured in EM-BaG was also evaluated during 15 days of culture. Live/dead staining demonstrated that DPPSCs in EM-BaG survived as well as DPPSCs in standard EM. In fact, at day 7 the cell amount of EM-BaG cultures increased. The same phenomenon was also observed by the proliferation assay at day 7, revealing that the increase in cell amount was also statistically significant (Fig. 2B).
In general, an upregulation of all the endothelial markers (FLK1, vWF, and CD31) was observed by qRT-PCR at 7 and 15 days of culture in EM-BaG compared with cultures in the standard EM (Fig. 4A). At protein level, CD31 and vWF IF images showed that BaG extracts increased the expression of both endothelial proteins (Fig. 4B). Moreover, although there were no significant differences in VEGF secretion between both culture conditions, at 15 days of differentiation, EM-BaG seemed to induce more secretion of VEGF (Fig. 4C).
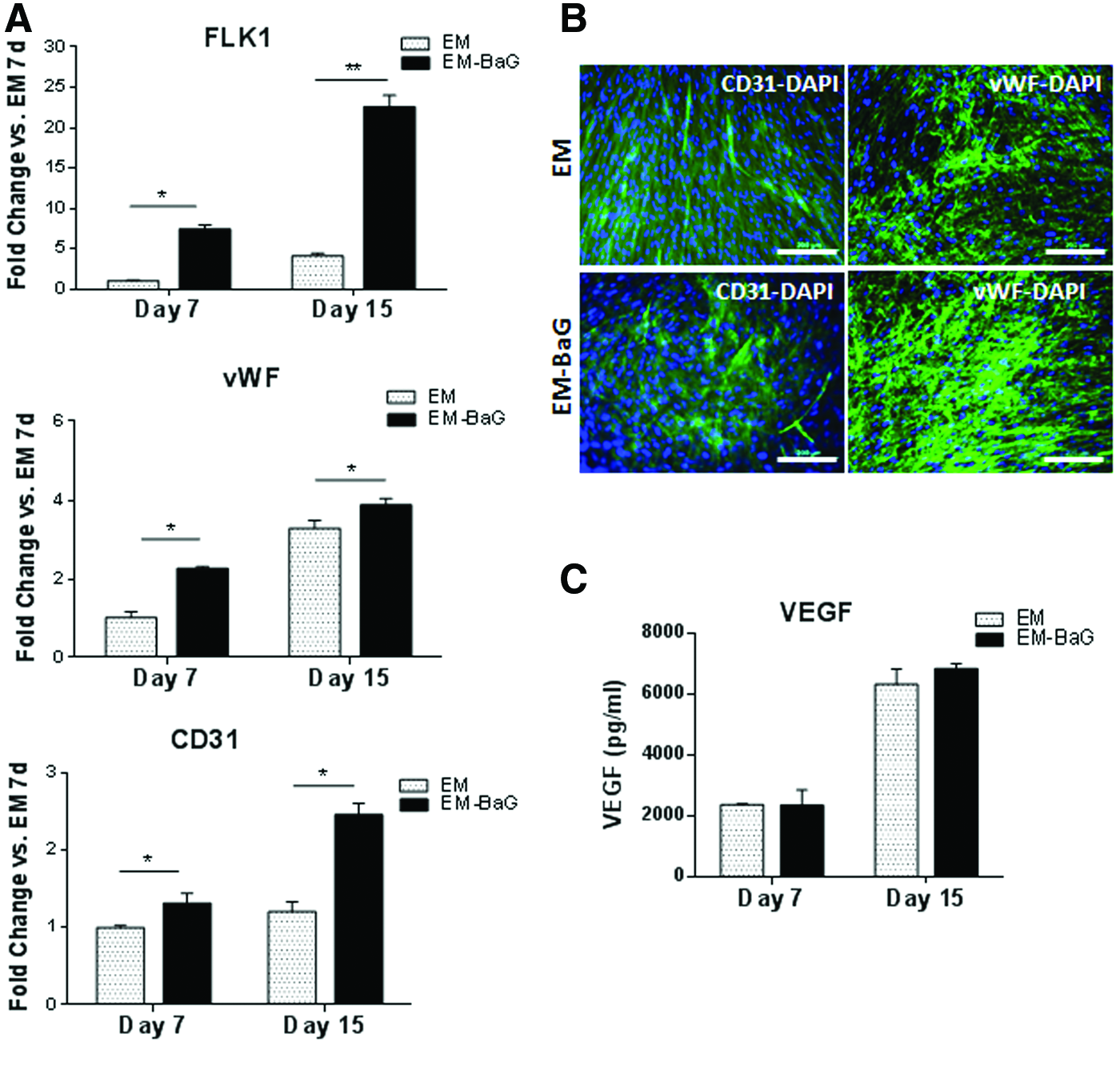
Effect of BaG extracts in the endothelial differentiation of DPPSC.
DPPSC coculture can enhance angiogenesis and osteogenesis simultaneously under the effect of BaG extracts.
DPPSCs show a good osteogenic and endothelial potential in xeno-free culture conditions, even better in the presence of BaG. Hence, we suggested the combination of both osteogenic and endothelial xeno-free DPPSC differentiations to induce vascularization of the bone-like tissue. In addition, we performed a preliminary study evaluating the effect of the coculture of DPPSC with a primary endothelial cell line, HUVEC (Supplementary Fig. S3).
Endothelial markers staining after 2 weeks of DPPSC coculture in different medium conditions showed that although vWF was expressed in a similar manner in all conditions, cocultures in EM with BaG extract resulted in a considerable formation of a vWF network (Fig. 5A). On the contrary, cocultures in OM or BM and monocultures in OM were characterized by the absence of CD31 expression (Fig. 5B). Thus, there was CD31 expression only under EM or OM/EM conditions. Moreover, the expression of CD31 was increased in cocultures with EM and OM/EM under the effect of BaG extracts resulting in a considerable formation of angiogenic-like structures, also observed at less magnification (Fig. 5D).
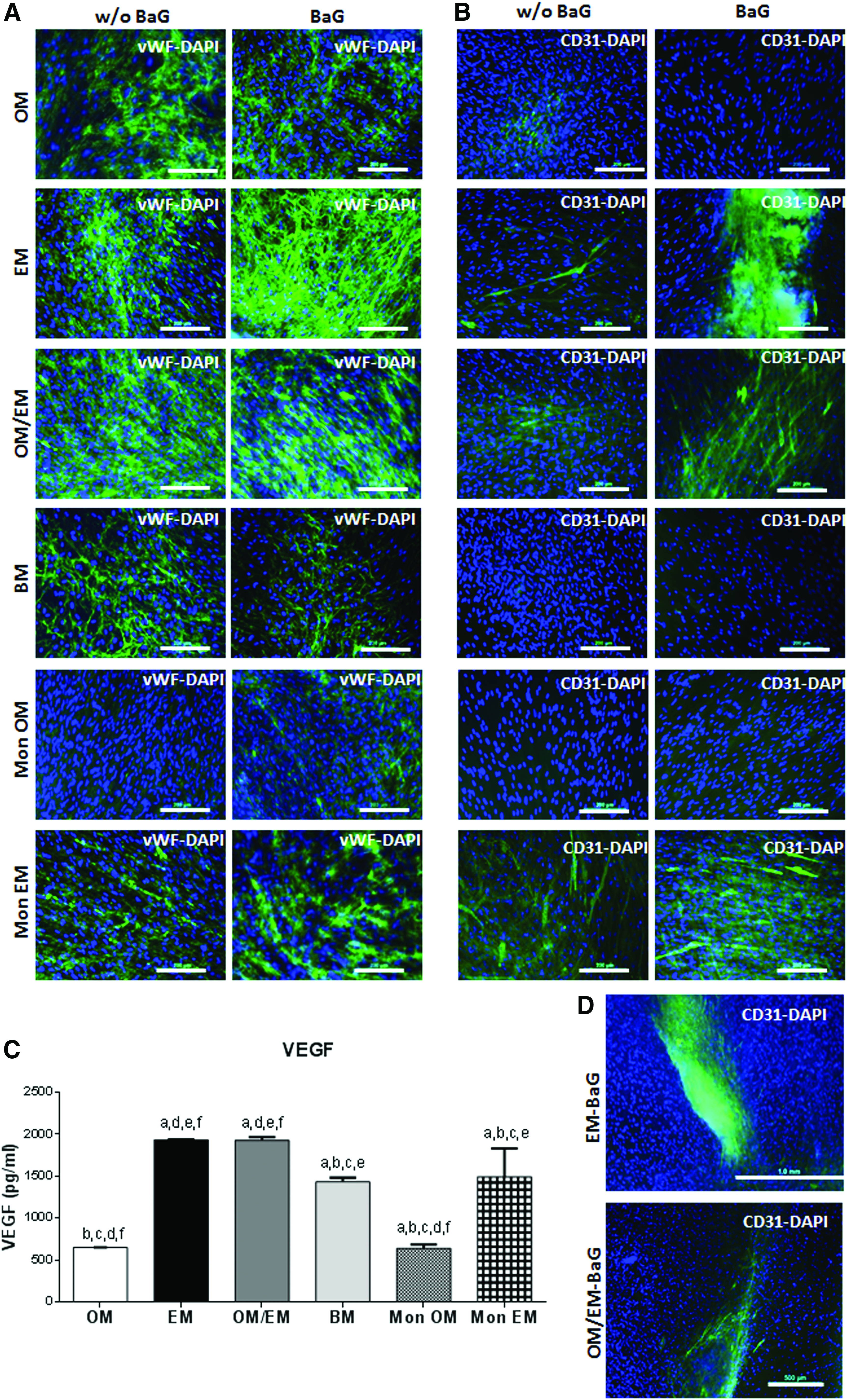
Effect of BaG extracts in the endothelial characteristic of 2D DPPSC cocultures.
The production of VEGF was increased in EM and EM/OM cocultures and in EM monocultures (Fig. 5C). By qRT-PCR, an upregulation of the endothelial markers (FLK1, vWF) was observed in DPPSC cocultures in either EM or OM/EM BaG conditions (Fig. 6A). Concretely, the expression levels of FLK1 increased in EM with the highest levels in BaG extracts conditioned media. On the contrary, vWF was upregulated in EM and EM/OM cocultures also with higher levels in the presence of BaG extracts.
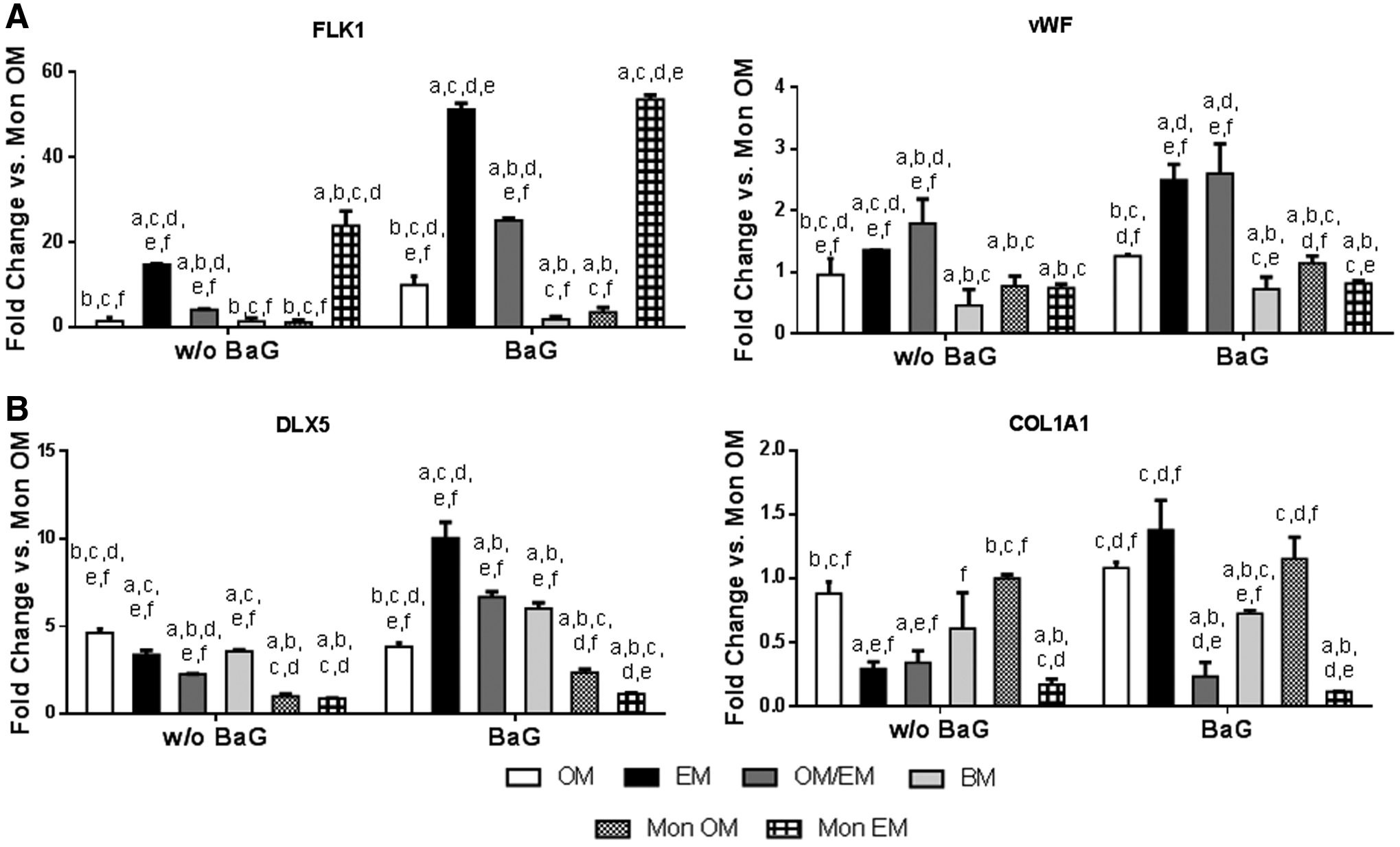
Effect of BaG extracts in the expression of osteogenic and endothelial marker genes of 2D DPPSC cocultures. Gene expression after 14 days of culture by qRT-PCR of
In addition, the coculture of DPPSCs might have effects on osteogenic differentiation. By qRT-PCR, standard cocultures showed the highest expression of the osteogenic markers DLX5 and COL1A1 in OM and BM conditions. However, there were changes in the expression profile of the osteogenic genes in response to BaG treatment. In general, BaG extracts upregulated the expression of the osteogenic markers in all cocultures, reaching the highest levels in EM conditions (Fig. 6B). Finally, Alizarin Red S staining showed that cocultures in EM with BaG extracts induced extensive mineral formation. The difference was significant compared with the other culture conditions as seen after quantification (Fig. 7).
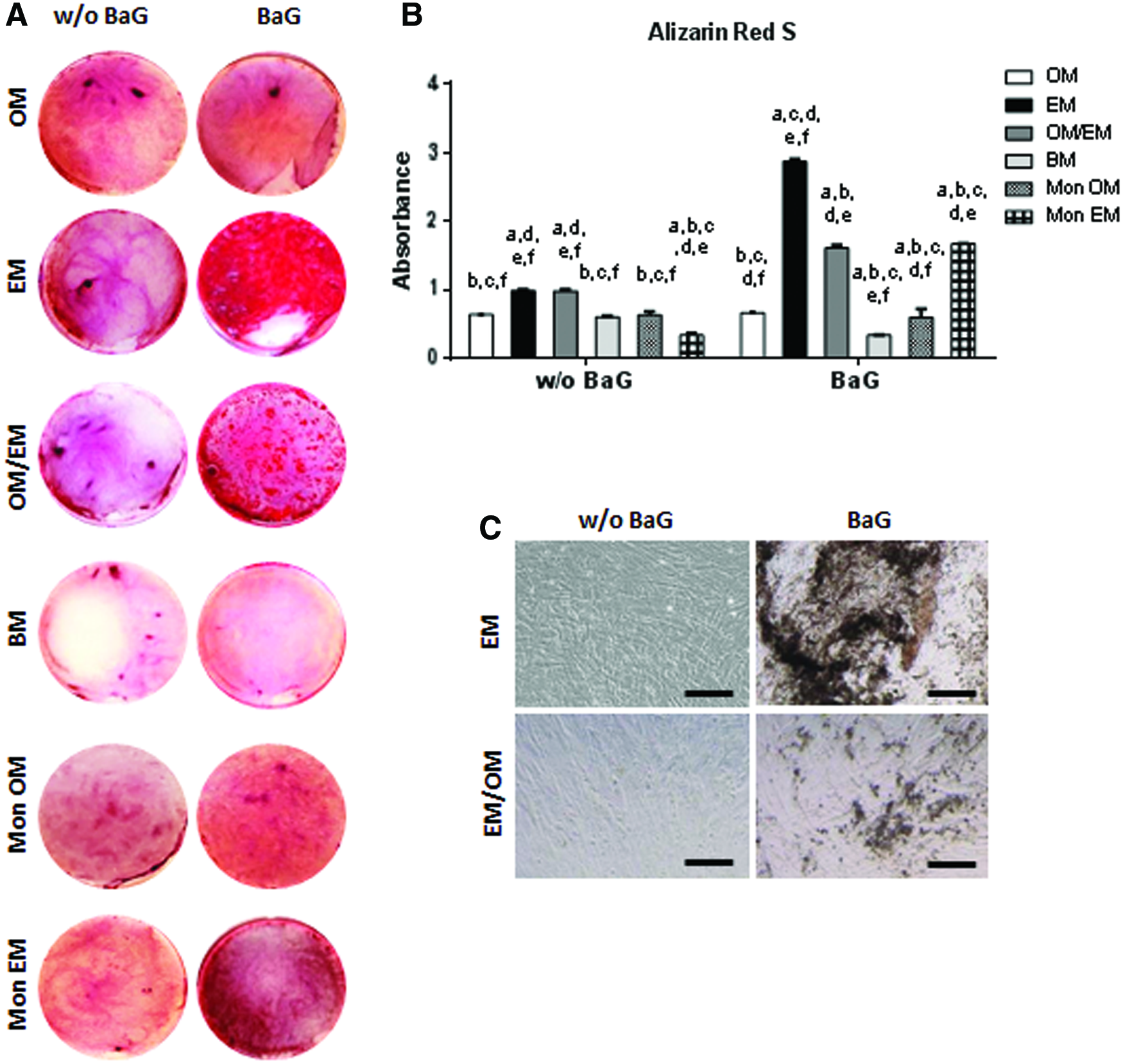
Effect of BaG extracts in the mineralization of DPPSC cocultures.
Three-dimensional DPPSC cocultures can support the formation of vascular-like structures under the effect of BaG
DPPSC monocultures and cocultures were analyzed in a 3D peptide–hydrogel system. For coculture experiments, we compared EM and EM-BaG, because they demonstrated the highest osteogenic and endothelial potential in the 2D experiments.
Cell viability of PuraMatrix encapsulating DPPSC monocultures showed that both osteogenic and endothelial culture conditions supported the viability of the cells (Fig. 8A). On the contrary, while DPPSC in EM acquired an elongated morphology, DPPSC in OM remained in a more rounded form. In fact, in EM, some interconnected endothelial cell structures were found. Phalloidin staining after 14 days of coculture (Fig. 8B–D) showed that a significant amount of cells was aligned in an interconnected cell structure, forming a complex network with a series of branching points. In addition, under the effect of BaG, DPPSCs seemed to form tubular-like structures in the presence of an internal lumen (Fig. 8C, D).

Cell alignment and tubular-like structures in DPPSC 3D cocultures.
Discussion
BaG and their dissolution ions have been shown to be advantageous biomaterials for BTE due to their osteogenic and angiogenic potential. 21 Reactions on the BaG surface induce the release of soluble ions such as Si, Ca, and P, leading to favorable cellular responses promoting rapid bone formation. 22 In this study, we have investigated, for the first time, the effect of S53P4 BaG extracts on the osteogenic and endothelial differentiation of DPPSCs and their coculture.
The first results demonstrated that DPPSC response to BaG ions depends of the medium conditions, it is necessary to add supplements to enhance differentiation. Moreover, BaG-conditioned media maintained the viability of DPPSCs, and enhanced both osteogenic and endothelial differentiations. However, at the beginning of the process (day 7), the proliferation rate was decreased under osteogenic conditions, while in the endothelial induction it was increased. Similar to our results, the combination of the inorganic ions from BaG and OM has been shown to enhance osteogenesis and mineralization in MC3T3-E1, fibroblasts, and adipocyte stem cells (ASCs).20,23,24 Controversial results exist in concern to proliferation: the proliferation of MC3T3-E1 and human osteoblasts was increased when they were cultured with 45S5 extracts,24,25 whereas in SAOS-2 with MBG85 extracts or ASC with S53P4 extracts, the proliferation was decreased.20,26 This large variation in the studies conducted with BaG extracts could be explained by the experimental setup differences (e.g., glass compositions, medium supplements, cell types).
On the contrary, our results of endothelial differentiation are in agreement with other studies that have reported a mitogenic response and an increase of angiogenic markers through both direct and indirect contact of human endothelial cells with BaG.19,27 We used a small concentration of BaG (0.08% w/v) for stimulating the endothelial potential of DPPSC. Previous reports have shown that the angiogenic potential of BaG is dose dependent. Hence, concentrations of 45S5 BaG superior to 0.1% w/v inhibited VEGF secretion from fibroblasts, possibly due to cytotoxic effects related to either increased concentration of ion dissolution products or increased pH. 28 Here, we demonstrated that S53P4 BaG extracts at 0.08% w/v could potentially provide an effective way to differentiate DPPSC toward both osteogenic and endothelial lineage.
The intimate functional relationship between bone vascular endothelium and osteoblasts might be crucial for bone development and remodeling. 29 Osteoblasts and osteoprogenitor cells are always located adjacent to endothelial cells of blood vessels at the site of bone formation. 30 Therefore, the communication between osteogenic and endothelial cells is the theoretical basis of the in vitro coculture systems for the prevascularization of bone tissue. It is known that during coculture, endothelial cells secrete soluble factors, such as BMP2, bFGF, ET1, or IGF, which affect the migration, proliferation, and differentiation of the osteogenic cells.31–33 Simultaneously, osteogenic cells produce high levels of VEGF, which has been shown to increase proliferation, differentiation, and angiogenesis of the endothelial cells. 34 However, some studies revealed significant differences in the stability and function of the cocultured cells depending on the culture medium, which seems to correlate with the secretion of the endothelial and the osteogenic factors. 35 In general, it has been reported that BM and OM are beneficial for the expansion and the differentiation of osteoblast-like cells. In contrast, survival and maintenance of endothelial cells are highly dependent on EM. Nevertheless, there is no consensus on the optimal media composition for the coculture of both cell types.2,35 Some reports demonstrated an increase in proliferation, VEGF production, and high expression of bone differentiation markers in HUVEC–bone-like cells cocultures under EM, BM, or EM/BM,5,36,37 some of them also demonstrated the formation of microcapillary-like structures.5,37
There are few studies on cocultures of cells originated from a single stem cell population.10,11 Here, once we demonstrated the effect of S53P4 BaG ions on DPPSC differentiations, cocultures of bone-like and endothelial-like DPPSCs with inorganic ions dissolved from BaG were investigated as a new strategy to induce vascularized bone tissue in vitro from a unique cell source. In general, the expression of endothelial markers and the formation of angiogenic structures were improved in the presence of EM and BaG (in either EM or EM/OM conditions) correlating with a higher release of VEGF. Besides, the differentiation of DPPSC toward the osteogenic lineage was more effective in EM conditions. Therefore, it suggests that the interaction between both the cell types was dependent on culture medium, and the dissolution of S53P5 BaG ions in the EM enhances the prevascularization and, consequently, accelerates the osteogenic development.
A 3D hydrogel scaffold might offer an optimal environment for cell-to-cell communication, cell migration, and cell functionality promoting the formation of vessel-like structures. 29 Three-dimensional DPPSC cocultures with EM and BaG were also performed in a peptide–hydrogel system demonstrating the formation of tube-like structures characterized by the definition of a vascular-like lumen. Fuchs et al. confirmed that when cocultured with human osteoprogenitors on plastic surface, endothelial cells are organized in vascular-like structures, but these do not exhibit a lumen at the center. In contrast, when cultured in 3D silk fibroin scaffolds, they form capillary-like networks containing vascular lumen, 38 a critical feature of angiogenically activated cells for establishing a functionally active blood supply. 29 Dissanayaka et al. also demonstrated a coordinated migration of undifferentiated DPSC and HUVEC in PuraMatrix, which supported cell survival, migration, and capillary network formation. 39 Although further studies testing the efficacy of 3D DPPSC cocultures using more accurate methods are necessary to demonstrate their effect in both angiogenesis and osteogenesis, here we observed a direct interaction between bone-like and endothelial-like DPPSCs in 3D conditions supporting the formation of vascular-like structures under the effect of BaG ions. Taken together, S53P4 BaG extracts in xeno-free EM could be an effective way to enhance both osteogenic and endothelial processes simultaneously, providing a new promising system for the in vitro vascularization of BTE constructs.
Footnotes
Acknowledgments
This work was partially funded by the Universitat International de Catalunya (UIC Barcelona) and the University of Tampere (UTA). UIC acknowledges Agència de Gestió d'ajuts Universitaris i de Recerca, Generalitat de Catalunya (SGR 2014) n° 1060. The authors R. Núñez-Toldrà and B. Bosch were funded by the predoctoral grant Junior Faculty awarded by the Obra Social La Caixa and the UIC. The author R. Núñez Toldrà was also funded by a travel grant awarded by Boehringer Ingelheim Fonds. The work at UTA was funded by Tekes, the Finnish Funding Agency for Innovation, the Academy of Finland, and the Competitive State Research Financing of the Expert Responsibility area of Tampere University Hospital.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.