
Abstract
Osteocytes play a key role in orchestrating bone homeostasis and turnover and, therefore, in vitro investigations with osteocytes are of high relevance for biomaterial and drug testing in future. In this study, collagen type I gels and collagen gels modified with biomimetically mineralized collagen were tested as three-dimensional (3D) environment for the maintenance of the osteocytic phenotype of primary human osteocytes. After cultivation in different collagen gels, cells were analyzed microscopically for the osteocytic phenotype and gene expression of osteocyte marker genes osteocalcin, podoplanin (PDPN)/E11, phosphate regulating endopeptidase homolog, X-linked (PHEX), matrix extracellular phosphoglycoprotein (MEPE), dentin matrix protein 1 (DMP-1), and sclerostin (SOST). Directly after isolation from bone tissue the cells expressed all examined osteocyte markers. After 7 days of 3D cultivation in collagen gels the osteocytic marker genes MEPE and SOST were upregulated and the other marker genes still expressed. Modification of collagen gels with biomimetically mineralized collagen and strontium-doped mineralized collagen prevented the cell-seeded gels from shrinking. Osteocyte morphology was not affected by the gel modification. However, the isolation of RNA from the mineralized gel variants was heavily impaired. Alternatively, the osteocytic differentiation of human osteoblasts in the different collagen gels was examined. Primary human osteoblasts were embedded into the gels and cultivated under osteogenic stimulation. After 14 days of cultivation, embedded osteoblasts showed osteocyte-like morphology and positive staining for DMP-1. Early osteocyte marker genes, such as PDPN/E11 and PHEX, were expressed while the expression of the osteoblast marker gene alkaline phosphatase (ALPL) increased. This ALPL upregulation was partly prevented by modification of collagen gels with mineralized collagen.
Impact Statement
This research focuses on the in vitro three-dimensional cultivation of primary human osteocytes instead of rodent osteocyte cell lines. Stable in vitro cultures of these regulating cells provide the opportunity to establish co- and triple cultures with osteoblasts and osteoclasts to analyze the cross talk between these cell species and to establish in vitro bone models for the testing of bioactive molecules, growth factors, drugs, and biomaterials.
Introduction
Osteocytes, which are terminally differentiated osteoblasts embedded into the mineralized bone matrix, are the most abundant cell type in mammalian bone comprising 90–95% of the bone cells. 1 Osteocytes are the key player in the orchestration of bone remodeling 2 ; they are mechanosensory cells that coordinate adaptive skeletal changes in response to mechanical loading 3 and function as endocrine cells. 4 Despite their high abundance in bone tissue, their localization deeply embedded into the rigid bone matrix prevents easy observation and isolation of these cells. Owing to these difficulties, the majority of studies on osteocyte function are performed with murine cell lines such as MLO-Y45 and IDG SW3. 6
Only few groups have successfully isolated primary human osteocytes so far.7–9 Consistently, low yield of osteocytes after performing the tedious multistep isolation was reported. Another option to generate primary osteocytes in vitro is the osteocytic differentiation of osteoblasts in a suitable three-dimensional (3D) environment. Atkins et al. performed a couple of experiments in which primary human osteoblasts were cultivated in collagen gels under osteogenic stimulation. They observed dendritic morphology of the cells after 7 days of cultivation with increased expression of osteocyte marker genes after 21 days of cultivation. 10 Furthermore, a promoting effect of vitamin K on osteocyte morphology leading to longer cell processes 11 and an increased expression of osteocyte markers in the presence of strontium (Sr) ranelate 12 are reported. Instead of primary human osteoblasts Uchihashi et al. used cells of a mouse osteoblast cell line that were cultivated on the surface of collagen gels. 13 An active migration of the cells into the gel and an increase of osteocyte markers gene expression was reported.
In a previous article, the Atkins group has systematically investigated the transition of human osteoblasts to osteocytes based on both primary osteoblasts and a human osteoblast cell line (SaOS-2). 14 Both, primary human osteoblasts and SaOS-2 cells, showed increasing gene expression of osteocyte markers when cultivated on tissue culture polystyrene under osteogenic differentiation for several weeks. Interestingly, podoplanin (PDPN)/E11, which is one of the main osteocyte markers and responsible for cytological reorganization, was expressed only in primary cell cultures and not in SaOS cultures. Furthermore, regarding the SaOS cells, there were no huge differences between two-dimensional (2D) and 3D cultivation.
The aim of this study was to generate a primary human osteocyte network in vitro, which can be applied for studies on osteocyte function. Therefore, both mentioned strategies—3D cultivation of primary osteocytes and 3D osteocytic differentiation of human osteoblasts—were compared. Primary osteoblasts and osteocytes were isolated from bone tissue. To increase the yield of primary osteocytes a modified isolation protocol was used, providing resting phases between the digestion/demineralization steps of the bone matrix. 15 The focus of the osteocyte cultures was the maintenance of the osteocytic phenotype during further cultivation in collagen gels, whereas for the osteoblast cultures the differentiation of the cells into osteocytes should be achieved. We used collagen gels for 3D environment of both strategies and modified them by addition of biomimetically mineralized collagen, with and without Sr modification. Our hypothesis was that mineralized collagen might trigger osteocytic differentiation of osteoblasts and might delay the dedifferentiation of primary osteocytes since its composition is similar to natural bone tissue.
Materials and Methods
Isolation of primary human osteocytes
Osteocytes were isolated from human femoral heads of three osteoarthritic patients undergoing total hip replacement at the University Hospital Carl Gustav Carus Dresden (Germany) after informed consent (approval by the ethics commission of TU Dresden, EK 303082014). The procedure was performed based on a protocol of Prideaux et al. with some modifications. 15 Spongious bone fragments were repeatedly digested (collagenase II treatment) and demineralized (EDTA treatment). Resting steps of the particles in between the treatments increased the yield of osteocytes.
Isolated primary osteocytes after at least six digestion steps were cultivated on collagen-coated TCPS for 2 days with Minimal Essential Medium, α-modification (α-MEM), 2% fetal calf serum, 2 mM
Osteoblasts were isolated after two collagenase II digests and were expanded in α-MEM, 15% fetal calf serum, 2 mM
Preparation of collagen gels and modified collagen gels
Pure collagen gels were prepared by mixing nine parts of acid collagen solution (3 mg/mL rat collagen; Amedrix, Esslingen, Germany) with one part of 10× Hank's Balanced Salt Solution (HBSS) and neutralizing with 1 M NaOH. Furthermore, β-glycerophosphate was added after neutralization to a final concentration of 10 mM in the gel.
Modified collagen gels were prepared by the addition of mineralized collagen to the neutralized collagen solution. Biomimetically mineralized bovine collagen was prepared as already published. 16 In brief, acid-soluble collagen I (isolated from bovine tendon and kindly provided by Syntacoll, Saal/Donau, Germany) was dissolved in 10 mM hydrochloric acid and mixed with a 0.1 M calcium chloride solution. The pH was adjusted to 7 by addition of Tris(hydroxymethyl)-aminomethan buffer (TRIS buffer) and phosphate buffer and the mixture was warmed to 37°C for 12 h. Under these conditions the collagen fibril reassembly and formation of nanocrystalline hydroxyapatite (HA) occurs simultaneously. The reaction was performed under air, so carbonated HA was formed. The product—a nanocomposite consisting of homogeneously mineralized collagen fibrils—was collected by centrifugation as a wax-like colorless material. A modification of biomimetically mineralized collagen was generated by the introduction of Sr2+ during simultaneous mineralization and collagen fibril assembly as previously described. 17 50% of the Ca2+ ions applied in the mineralization setup were replaced with Sr2+ leading to a Sr content of 32 mol% related to the Ca2+ content. After centrifugation the materials were freeze-dried, grounded with a mortar, and sterilized with supercritical CO2. 18 The powder was added to the neutralized collagen solution in a concentration of 2 mg/mL.
Cultivation of osteocytes and osteoblasts in collagen gels
Primary osteocytes and primary osteoblasts were embedded in collagen gels and modified collagen gels (which were prepared as described in the previous paragraph) at a concentration of 1 × 105 cells/mL. For microscopic investigations, gels of 200 μL collagen in 48-well plates and for the isolation of RNA gels of 1 mL in 24-well plates were prepared. Cell-laden collagen samples were allowed to gel for 30 min at 37°C in the incubator before culture medium was added. Culture medium for osteocytes consisted of α-MEM, 2% fetal calf serum, 2 mM
Fluorescence microscopy
Gels were fixed with a 4% solution of neutral buffered formaldehyde for 1 h at room temperature. After permeabilization using 0.1% Triton X-100 in phosphate-buffered saline (PBS) for 5 min and five washes with PBS, samples were blocked using 1% bovine serum albumin (BSA) in PBS for 30 min. A solution of 1 μg/mL rabbit monoclonal (Clone DMP-1) anti-DMP-1 antibody (M176; TaKaRa Bio, Kusatzu, Japan) in PBS containing 1% BSA was incubated with the samples overnight at 4°C. Antibody solution was removed and after five washes with PBS the samples were incubated with a solution of 20 ng/mL DAPI (4′,6-diamidin-2-phenylindol; Invitrogen), 25 μL/mL AlexaFluor 488® phalloidin (Invitrogen, part of Thermo Fisher) and 4 μL/mL of goat anti-rabbit AlexaFluor 546 phalloidin (Invitrogen, part of Thermo Fisher) overnight at 4°C protected from light. After removal of the staining solution, the samples were finally washed with PBS and imaged with a Keyence BZ 9000 fluorescence microscope.
RNA isolation
Cell-seeded collagen gels (six 1 mL-gels per condition and time point) were incubated with collagenase II solution (235 U/mg; Biochrom, Berlin, Germany, part of Merck, Germany) (3 mg/mL in α-MEM, 10% fetal calf serum, 2 mM
cDNA synthesis and quantitative real-time polymerase chain reaction
cDNA was generated using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher) according to manufacturer's instructions. In total 50–100 ng RNA was transcribed, according to the yields of RNA per experiment.
Polymerase chain reaction (PCR) reactions were set up using the TaqMan Fast Advanced Master Mix (Thermo Fisher) and TaqMan Gene Expression Assays for glyceraldehyde-3-phosphate dehydrogenase (GAPDH), alkaline phosphatase (ALPL), bone gamma-carboxyglutamate protein (osteocalcin, BGLAP), PDPN (E11/g38), phosphate regulating endopeptidase homolog, X-linked (PHEX), matrix extracellular phosphoglycoprotein (OPF 45, MEPE), dentin matrix protein 1 (DMP-1), and sclerostin (SOST) (Thermo Fisher) according to manufacturer's instructions.
PCR was run with an Applied Biosystems® 7500 fast Real-Time PCR system (Applied Biosystems).
Relative gene expression (fold-change) was calculated using the 2−ΔΔCt method and normalized to GAPDH expression.
Statistical differences of different groups were calculated at the level of ΔCT values using t-test for the comparison of two groups and one-way ANOVA followed by post hoc Tukey test to determine multiple comparisons (Origin 9.1, OriginLab).
Results
Maintenance of osteocyte phenotype in collagen gels and modified collagen gels
Primary osteocytes, embedded in collagen gels, and collagen gels modified with mineralized collagen, and Sr-doped mineralized collagen showed osteocytic morphology and positive staining for DMP-1 (Fig. 1). In all gel variants cells showed dendritic processes, which are typical for osteocytes. In the mineralized gel variants, the processes showed a tendency to be shorter compared with the processes of osteocytes in pure collagen gels. There were no changes in cell morphology between 7 and 14 days of cultivation (data not shown).
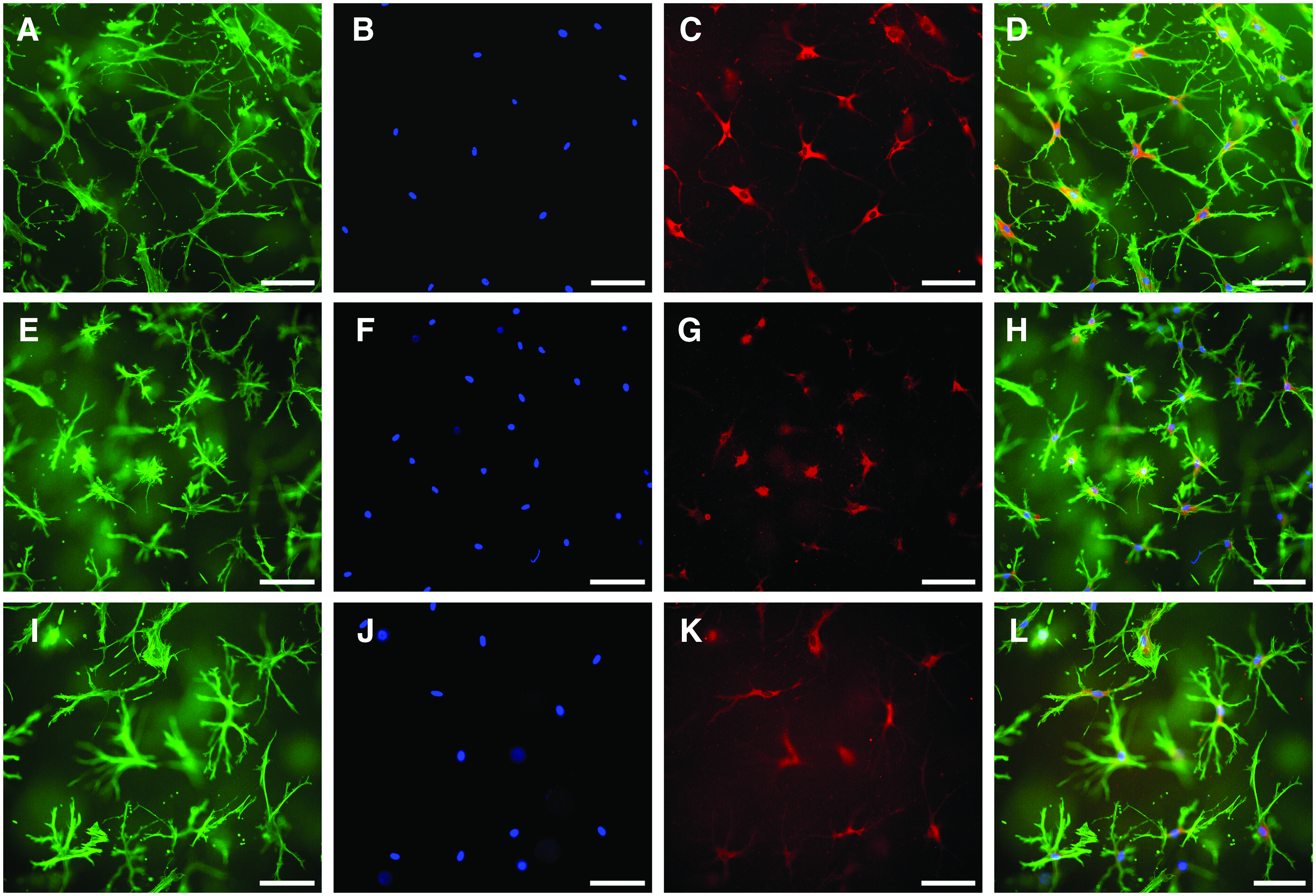
Human osteocytes embedded into collagen gels
During cultivation in collagen gels gene expression profile of the osteocytes changed (Fig. 2). Gene expression of the osteogenic marker ALPL further decreased as well as osteocalcin expression, which is a marker of late osteoblasts and early osteocytes. PHEX expression remained quite stable, whereas PDPN/E11, MEPE, and SOST expression even increased. DMP1 expression was significantly downregulated during cultivation in the collagen gel.

Relative expression of osteoblast marker gene ALPL and osteocyte marker genes in primary osteocytes that were cultivated for 7 days in collagen gels compared with d0. Data of two different cell donors are presented in the left and right diagram. Six samples were analyzed per donor, with two samples pooled for RNA isolation. Mean ± upper/lower limit. Significant differences to day 0 (*p < 0.05, **p < 0.01). ALPL, alkaline phosphatase.
Gene expression of osteocytes embedded in mineralized collage gels was not analyzed, since the yield of RNA obtained from these collagen constructs was not sufficient to obtain conclusive results.
Differentiation of human osteoblasts into osteocytes in collagen gels and modified collagen gels
Human osteoblasts, obtained after two collagenase digestion steps of bone particles showed osteocytic morphology after 14 days of cultivation in different collagen gels under osteogenic stimulation (Fig. 3). There was also a weak DMP1 staining detectable. In some cases, gels shrank during cultivation (Fig. 4), and only gels that maintained their full size were included into the investigations. Interestingly, modified gels, both with mineralized collagen and Sr-doped mineralized collagen, did not show any shrinkage during the whole observation period.
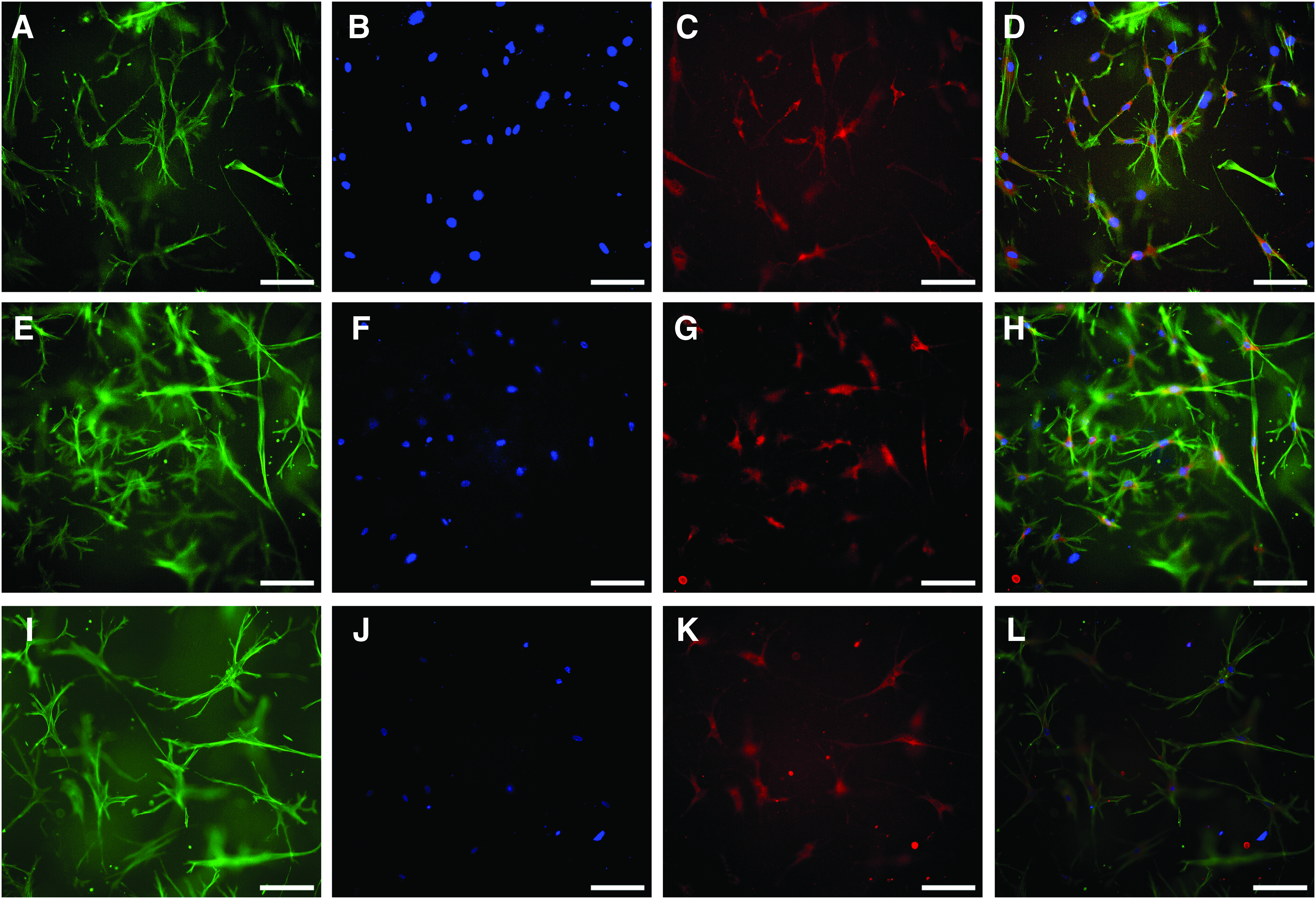
Human osteoblasts were embedded into collagen gels
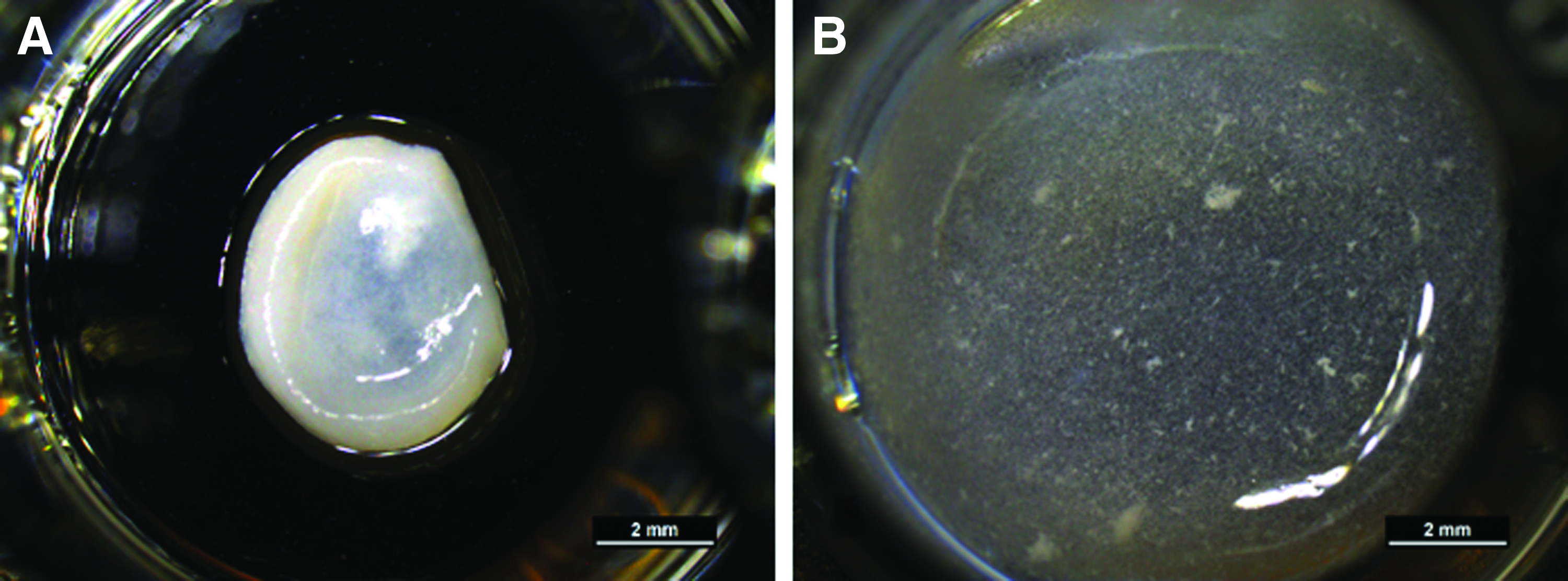
Shrinkage of collagen gels. Primary osteocytes were embedded into collagen gels and collagen gels modified with 2 mg/mL mineralized collagen and cultivated for 7 days. In some cases, the pure collagen gels
In all experiments with nonmodified collagen gel, the expression of the osteoblast marker ALPL significantly increased (Fig. 5A). We furthermore observed an increase of osteocyte markers PDPN/E11 and PHEX. In contrast, BGLAP gene expression was significantly downregulated in all experiments during cultivation in collagen gels. Gene expression of the osteocyte markers MEPE, DMP-1, and SOST was not detected. Further cultivation of the osteoblasts in collagen gels for up to 4 weeks revealed a further increase of both ALPL and PHEX gene expression (Fig. 5B). However, even after 4 weeks of osteogenic differentiation of osteoblasts in collagen gels gene expression of the osteocyte markers MEPE, DMP-1, and SOST was not detected.

Expression of osteoblast marker gene ALPL and osteocyte marker genes in osteoblasts that were cultivated in collagen gels for 14 days (
In the case of mineralized collagen gels similar gene expression profiles were found (Fig. 6). ALPL, PDPN, and PHEX expression increased compared with d1, and BGLAP expression significantly decreased, whereas later osteocyte markers DMP1, MEPE, and SOST were not detected. Interestingly, ALPL expression was reduced when the cells were cultivated in collagen gels modified with mineralized collagen compared with pure collagen gels. Although ALPL expression was significantly upregulated in all gel modifications for the cells of one donor, the effect was not statistically significant for the cells of the other examined donor. ALPL expression was significantly lower in the Sr-doped mineralized collagen gel compared with the pure collagen gel (p = 0.00518) and mineralized collagen gel (p = 0.0304).

Expression of osteoblast marker gene ALPL and osteocyte marker genes BGLAP, PDPN, and PHEX in primary osteoblasts, which were cultivated for 14 days in collagen gels and collagen gels modified with mineralized collagen under osteogenic stimulation. Data of osteoblasts from two different donors are shown in diagram
Discussion
In this study, primary human osteocytes were cultivated in vitro in different collagen gels. Collagen as the major structural protein of bone tissue is frequently used to support in vitro cultivation of osteocytes. Murine osteocyte cell lines are maintained on collagen-coated dishes, as proposed by the group of Bonewald, 5 and this cultivation can also be used to maintain the morphology of primary human osteocytes for a few days. 8 3D cultivation of osteocytes embedded in collagen gels seems to support the osteocytic phenotype even better. It was demonstrated that primary murine osteocytes show higher expression of osteocyte markers SOST and FGF-23 when cultivated embedded in collagen gel instead of on top of collagen-coated plates. 19 However, the expression of osteocyte markers PDPN and DMP-1 was maintained to a higher extent in the 2D culture, and also in 3D environment the cells dedifferentiated over time. In this study, primary osteocytes were embedded in collagen gels, and after 7–14 days of cultivation the cells showed still an osteocytic phenotype and a positive staining for DMP-1. Some osteocytic markers, such as PDPN/E11, MEPE, and SOST were even upregulated during 3D cultivation in collagen. Interestingly, also ALPL gene expression, which was quite low at from the beginning of the experiment, was further downregulated during cultivation of osteocytes in the 3D gels, suggesting that cultivation inside a collagen gel supports the osteocytic phenotype rather than the osteoblastic phenotype. Unfortunately, also DMP1 gene expression was downregulated during 3D collagen cultivation of osteocytes, which is in contrast to the positive DMP1 immunostaining of the cells. We suppose that even low DMP1 gene expression leads to a positive signal in the immunostaining. Prideaux et al. embedded primary human osteocytes into collagen gels for 5 days and observed dendritic morphology of the cells with long extensions, similar to our study; however, they did not perform immunostaining for DMP1 or gene expression analysis. 8
An elegant method to obtain primary human osteocytes might be the in situ differentiation of osteoblasts into osteocytes in collagen or gelatine gels, as described by several groups.10,13,14,20,21 In this study, primary human osteoblasts, embedded into collagen gel and cultivated for up to 28 days under osteogenic stimulation with dex, AAP, and β-GP showed typical osteocyte morphology and at least weak positive staining for the osteocyte marker DMP-1. However, of the examined osteocyte markers, only PHEX and PDPN/E11 were upregulated, indicating a beginning osteocyte differentiation. We were not able to detect noteworthy expression of osteocyte markers MEPE, SOST, and DMP-1, irrespective of the positive DMP-1 staining. PHEX and MEPE are enzymes that are involved in the regulation of phosphate homeostasis and extracellular matrix mineralization, and their expression highly increases during the maturation of osteoblasts to osteocytes. 22 It has furthermore been shown that inhibition of mineralization in late osteoblasts/preosteocytes can be induced by PHEX downregulation and MEPE upregulation, with MEPE being an inhibitor of PHEX. 23 We suggest incomplete maturation of osteoblasts in this experimental setup with only early osteocyte markers such as PDPN/E11, which is responsible for the elongation of osteocyte dendritic processes. 24
Accordingly, there was a strong upregulation of the osteoblast marker ALPL, and ALPL expression increased further between d14 and d28 of cultivation. Increased ALP activity or ALPL gene expression during cultivation of an osteoblast cell line (MC3T3-E1) in collagen or gelatin gels was also reported by others.20,21 However, during maturation of the osteoblasts and subsequent osteocytic differentiation ALP expression should be downregulated. 25 Probably, the cultivation time of maximum 28 days was too short to trigger appropriate osteoblast maturation, and it would be necessary to cultivate the constructs for longer periods in future studies.
Furthermore, the addition of dex, which is responsible for the strong increase of ALP, should be reduced in concentration or completely omitted in further studies. The lack of gene expression of osteocytic markers SOST, MEPE, and DMP-1 in differentiating osteoblast 3D cultures in this study is in contrast to already published studies. Other groups showed an upregulation of osteocyte markers DMP-1 and SOST after cultivation of osteoblast cell lines in collagen gels for 35 days,13,14 but also at earlier time points of cultivation. Besides the slightly longer cultivation time in the cited studies, both groups did not add dex to their mineralizing cultures. Possibly, the exclusion of dex could be helpful not only to suppress ALPL expression, but also to support expression of late osteocyte markers. When biomimetically mineralized collagen was present in the collagen gels ALPL expression of the cells increased less, compared with pure collagen gels, suggesting modified collagen gels as a promising environment for the osteocytic differentiation of osteoblasts. In this study besides mineralized collagen, a Sr-doped mineralized collagen was applied to modify collagen gels. It has been shown before that the addition of Sr ranelate to long-term (up to 42 days) monolayer cultures of primary osteoblasts under mineralizing conditions stimulated the expression of osteocyte marker genes DMP-1, SOST and BGLAP. 12
However, in this study osteoblasts that were embedded into collagen gels, modified with Sr-doped mineralized collagen, did not show any expression of these late osteocyte markers after 14 and 28 days of cultivation. Possibly the Sr concentration, which was released during cultivation from the gels, was not high enough to stimulate osteocytic differentiation. Sr2+ release from Sr-doped mineralized collagen was quantified with 6–8 μg/mg, 17 resulting in an initial Sr concentration of ∼0.1 mM for the modified collagen gels, containing 2 mg mineralized collagen per gel. Stimulation of osteocytic differentiation was observed in the presence of 5–10 mM Sr2+, 12 which is two magnitudes higher compared with our experiments.
Nevertheless, modification of collagen gels with biomimetically mineralized collagen has several advantages. The addition of mineralized collagen prevents cell-seeded collagen gels from shrinking. Shrinking of cell-laden collagen hydrogels has been observed before, when fibroblasts were cultured in collagen gels. 26 This phenomenon is explained by tension which is exerted by cell processes onto the collagen lattice. It has furthermore been observed that contraction of the collagen matrix increases with increasing cell number. With higher collagen content, shrinkage is diminished. In the case of osteocyte cultures, the cell number in the gels is quite low, and we have observed gel shrinkage only in a few samples of each experiment.
However, the addition of biomimetically mineralized collagen completely prevented shrinkage, which might be the result of an increased collagen content leading to a higher stiffness of the samples. A recent article describes the modification of collagen gels with small amphiphilic molecules, which also resulted in the prevention of shrinkage, 27 suggesting that interactions of cells with the collagen matrix can be modulated by different molecules. A further promising result of this study is the downregulation of ALPL expression in cells, which were cultivated in mineralized collagen gels. This result was quite surprising, since HA is osteoconductive, supporting the osteogenic differentiation and formation of new bone. Sr-substitution of HA even increases the potential to support osteogenic differentiation: Primary rat osteoblasts, which were cultivated on Sr-substituted HA ceramics, showed higher ALP activity compared with cells on pure HA ceramics. 28 Human mesenchymal stroma cells cultivated on Sr-doped HA cements showed higher ALP activity compared with cells on nondoped HA cements. 29 Recently, it has been shown that another calcium phosphate phase—octacalcium phosphate promotes the transition of osteoblasts to osteocytes and mineralization in vitro. 30
One reason for the decreased ALPL expression in the presence of mineralized collagen could be the diminished Ca2+ concentration in the cell culture medium. It has been demonstrated before that scaffolds from biomimetically mineralized collagen absorb Ca2+ ions from cell culture medium in vitro. 31 This is due to the conversion of Ca-deficient HA, which is embedded as nanocrystals into the collagen fibrils. In this study, the Ca2+ concentration in the cell culture supernatants of gels, modified with mineralized collagen, was significantly lower compared with the Ca2+ concentration of supernatants from pure collagen gels (Fig. 7). Low Ca2+ concentrations (0.15–0.19 mM, compared with 0.64–0.8 mM in the control medium) significantly reduced ALP activity of human SaOS-2-osteoblasts. 32

Ca2+ content of cell culture medium. Primary osteoblasts
Similar results were obtained with fetal rat calvaria osteoblasts; lowering the Ca2+ concentration from 1.2 mM to 0.8 and 0.5 mM significantly decreased the ALP activity of these cells. 33 The authors conclude that osteoblasts express a functional calcium receptor, regulating their activity. Interestingly, an upregulation of BGLAP in the presence of low Ca2+ concentrations was detected, which is in accordance with our results, where BGLAP expression is upregulated in the presence of mineralized collagen.
Conclusions
Primary osteocytes, embedded into collagen gels, show typical osteocytic morphology. Osteocytic morphology was also observed for osteoblasts, which were cultivated under osteogenic stimulation in collagen gels for 2 weeks. However, gene expression of osteocytic markers DMP-1, SOST, and MEPE was detected in osteocyte cultures but not in cultures of differentiated osteoblasts. Furthermore, in direct comparison osteoblast marker ALPL was higher in differentiated osteoblasts compared with primary osteocytes, whereas the early osteocyte marker osteocalcin was expressed to a higher level in osteocytes cultures compared with differentiated osteoblasts. Therefore, for further investigations, such as co-culture and triple culture studies with other bone cells the application of freshly isolated primary osteocytes is favored. Modification of collagen gels with biomimetically mineralized collagen prevents cell-laden gels from shrinking and suppresses ALPL gene expression; however, the formation or maintenance of the osteocytic phenotype is not better supported by the mineralized gel variants and RNA isolation from mineralized collagen gels is impaired.
Footnotes
Acknowledgments
We are grateful to the German Research Foundation (DFG) for funding this study (BE 5139/3-1) and thank Ortrud Zieschang for excellent technical assistance.
Disclosure Statement
No competing financial interests exist.