
Abstract
Cancer invasion, metastasis, and therapy resistance are the crucial phenomena in cancer malignancy. The high expression of matrix metalloproteinase 9 (MMP9) is a biomarker as well as a causal factor of cancer invasiveness and metastatic activity. However, a regulatory mechanism underlying MMP9 expression in cancer is not clarified yet. In addition, a new strategy for anticancer drug discovery is becoming an important clue. In the present study, we aimed (i) to develop a novel reporter system evaluating tumorigenesis, invasiveness, metastasis, and druggability with a combination of three-dimensional tumoroid model and Mmp9 promoter and (ii) to examine pharmacological actions of anticancer medications using this reporter system. High expression and genetic amplification of MMP9 were found in colon cancer cases. We found that proximal promoter sequences of MMP9 in murine and human contained conserved binding sites for transcription factors β-catenin/TCF/LEF, glucocorticoid receptor (GR), and nuclear factor kappa-B (NF-κB). The murine Mmp9 promoter (−569 to +19) was markedly activated in metastatic colon cancer cells and additionally activated by tumoroid formation and by β-catenin signaling stimulator lithium chloride. The Mmp9 promoter-driven fluorescent reporter cells enabled the monitoring of activities of MMP9/gelatinase, tumorigenesis, invasion, and metastasis in syngeneic transplantation experiments. We also demonstrated pharmacological actions as follows: dexamethasone and hydrocortisone, steroidal medications binding to GR, inhibited the Mmp9 promoter but did not inhibit tumorigenesis. On the contrary, antimetabolite 5-fluorouracil, a gold standard for colon cancer chemotherapy, inhibited tumoroid formation but did not inhibit Mmp9 promoter activity. Notably, antimalaria medication artesunate inhibited both tumorigenesis and the Mmp9 promoter in vitro, potentially through inhibition of β-catenin/TCF/LEF signaling. Thus, this novel reporter system enabled monitoring tumorigenesis, invasiveness, metastasis, key regulatory signalings such as β-catenin/MMP9 axis, and druggability.
Impact Statement
Cancer invasion and metastasis have been shown to be driven by matrix metalloproteinase 9 (MMP9), whose expression mechanism is not clarified yet. In addition, a new strategy for anticancer drug discovery is becoming important. We established a novel reporter system evaluating tumorigenesis, invasiveness, metastasis, and druggability with a combination of three-dimensional (3D) tumoroid model and Mmp9 promoter. Using this reporter system, we demonstrated pharmacological actions of anticancer medications such as antimetabolite 5-fluorouracil (5-FU) and antimalaria medication artesunate (ART), which inhibited both tumorigenesis and β-catenin/MMP regulatory signaling. Our study impacts the translational fields of oncology, drug discovery, and organoid model.
Introduction
Cancer is one of the most serious diseases all over the world, and its metastasis and therapy resistance are leading causes of death. A new strategy for anticancer drug discovery is becoming an important clue. Success in anticancer drug discovery depends on the appropriate experimental tumor models and screening system. Organoid model and tissue engineering have been devised by scientific/medical motivation for regenerative medicine and drug discovery. In the field of cancer study, the 3D spheroid/tumoroid culture system has enabled us to develop a tumor-like organoid (tumoroid) in vitro, corresponding closely to native tumors in vivo.1–9 MMPs represent the most prominent family of proteinases associated with tumorigenesis, 10 tumor microenvironment, 11 angiogenesis,12,13 migration, invasion, 14 and metastasis. 10 Canonical roles for MMPs are to cleave and proteolyze substrate proteins at extracellular space. Proteolysis of extracellular matrix (ECM) and intercellular adhesion molecules by MMPs increases cellular motility, migration, and invasion abilities. 15 Proteolysis of ECM also promotes the release of cytokines, chemokines, and growth factors that activate the receptors and intracellular signaling pathways. MMPs also directly alter the activities of growth factors, cytokines, and chemokines by proteolysis. 16 High expression of MMP9 and MMP3 has been reported in human colon cancer in late stages with poor prognosis of patients.17–19 We recently showed that MMP9 and MMP3 are markedly produced by rapidly metastatic colon cells and targeted depletion of these MMPs successfully inhibited tumor growth, invasion, and metastasis. 10 In addition, intracellular MMPs play key roles in transcriptional regulation,20–22 intracellular proteolysis, and migration of cancer cells. 10 These studies prompted us to pursue potential signals contributing to the high expression of MMP9, whose promoter is markedly activated in metastatic cancer cells in vivo and in vitro. We also aimed to utilize the powerful MMP9 promoter for monitoring tumorigenic, invasive, and metastatic activities of cancer cells as well as the development of a novel drug discovery system.
Drug screening, as well as tissue/cell culture, had been often carried out using two-dimensional (2D) monolayer culture on plastic plates, which unintentionally altered the characteristics of cells. For example, 2D culture markedly induces forced expression of integrins, which is required for attachment to such plate materials. 23 In contrast, 3D organoid/tissue culture systems are able to replicate many aspects of 3D organs, tumors, and their environment and are thus more suitable for many physiological and pathological studies.1,24,25 One such feature of 3D organogenesis is an enhancement of stem cell phenotype. 2 We showed that 3D organoid/tumoroid culture of cancer cells enhanced stem cell phenotype with enhanced expression of stem cell marker genes. 2 It has been also shown that the tumor microenvironment consists of cancer cells, including cancer-initiating cells (CIC) also known as cancer stem cells (CSC), which can play key roles in tumorigenesis, recurrence, resistance, heterogeneity, and metastatic ability. 26 The features of stem cells in both organogenesis and tumorigenesis have been mediated by Wnt/β-catenin/TCF signaling.27–30 Wnt/β-catenin/TCF signaling is often activated in colon cancer by mutations in β-catenin or adenomatous polyposis coli, a repressive cofactor of β-catenin. 31 β-catenin appears to control the renewal of CIC/CSC fraction through regulation of proteins such as cyclin D1.32,33 β-catenin is also regulated by control of its rate of degradation that is mediated through phosphorylation by the kinase GSK3 on an N-terminal destruction motif. Binding of the ligand Wnt to surface receptors Frizzled and LRP-5/6 triggers Dsh to inhibit GSK3 activity, prevents degradation, and permits the influence of β-catenin in CIC/CSC renewal. 34 By analyzing Mmp9 promoter sequence, we found a β-catenin/TCF binding motif conserved between human and mouse in the proximal promoter sequences of MMP9. In the present study, we therefore aimed to investigate the β-catenin/TCF/MMP9 regulatory axis in tumorigenesis.
The 3D culture system-based organoid/tumoroid formation is of general interest as well as useful for drug screening that targets resistant tumorigenesis with enhanced CIC/CSC phenotypes.2,25 We in the present study examined whether 5-FU, hydrocortisone (HC), and ART could inhibit in vitro tumorigenesis. 5-FU is the first-choice drug in the treatment of colon cancer due to its ability to inhibit the growth of cancer cells by incorporating its metabolites into DNA and RNA. 35 ART is part of the artemisinin group of drugs that treat malaria and is also active against cancer. 36 Recently, ART was identified as a selective inhibitor of cancer stemness. 37 We therefore hypothesized that ART could inhibit resistant/recurrent tumorigenesis through inhibiting stemness. HC is the pharmaceutical term for steroid hormone cortisol, in the glucocorticoid class of hormones, used as a steroidal anti-inflammatory drug (SAID) or an immunosuppressive drug. We examined this drug because (i) HC tended to inhibit in vitro tumorigenesis in our previous study 25 and (ii) HC binding to the glucocorticoid receptor (GR) represses activities of nuclear factor kappa-B (NF-κB) and AP1.38,39 We found consensus binding sequences of both GR and NF-κB in the Mmp9 promoter sequence, which we hypothesized could be repressed by HC.
Thus, the specific aims in the present study are (i) to develop a novel reporter system with a combination of 3D tumoroid model for drug discovery, (ii) to investigate whether a novel β-catenin/MMP9 regulatory axis could contribute to tumorigenesis with enhanced stemness, and (iii) to examine whether 5-FU, HC, and ART could inhibit in vitro 3D tumorigenesis in the tumoroid culture system.
Materials and Methods
Cell culture
A rapidly metastatic colon cancer cell line LuM1, its parental slowly metastatic cell line Colon26, and its nonmetastatic subline NM1110,40 were cultured in RPMI1640 with 10% fetal bovine serum (FBS) supplemented with penicillin, streptomycin, and amphotericin B or in mTeSR1 stem cell medium (STEMCELL Technologies, Vancouver, Canada)2,41 on 2D tissue culture plates (Corning, Corning, NY) or NanoCulture plates (NCPs) (MBL, Nagoya, Japan) that enable 3D culture.2,25,42
Chemicals and drugs
Lithium chloride (LiCl), dexamethasone (DEX), 12-O-tetradecanoylphorbol 13-acetate (TPA), and HC were purchased from Sigma-Aldrich (St Louis, MO). ART was purchased from Cayman Chemical (Ann Arbor, MI). AG490 was purchased from Tokyo Chemical Industry (Tokyo, Japan).
Reverse transcription quantitative real time polymerase chain reaction
Reverse transcription quantitative real time polymerase chain reaction (RT-qPCR) was performed as described. 2 Total RNA was prepared from biological quadruplicates. Polymerase chain reaction (PCR) was performed in triplicate. Primers used were mMmp9-Fw, 5′-CAGCCGACTTTTGTGGTCTT-3′ and mMmp9-Rv, 5′-GCTTCTCTCCCATCATCTGG-3′; mMmp2-Fw, 5′-GTCGCCCCTAAA ACAGACAA-3′ and mMmp2-Rv, 5′-GGTCTCGATGGTGTTCTGGT-3′; mMmp14-Fw, 5′-CCCTTTTACCAGTGG ATGGA-3′ and mMmp14-Rv, 5′-TTTGGGCTTATCTGGGACAG-3′; and b-actin-Fw, 5′-AACGAGCGGTTCCGATG-3′ and b-actin-Rv, 5′-GGATTCCATACCCAAGAAGGA-3′.
Promoter analysis and cloning
As described, 43 murine and human MMP9 promoter sequences were obtained from the Eukaryotic Promoter Database. Transcription factor binding sites were predicted using PROMO (ver 8.3). The two sequences were aligned using NCBI BLAST.
A 588-bp cDNA fragment of Mmp9 promoter between −569 and +19 was cloned via genomic PCR from a tail of a BALB/c mouse using the DNA isolation kit (Qiagen, Hilden, Germany). 44 A sense primer 5′-AAGGAGTCAGCCTGCTGGAGCTAGGGGTTTGC-3′ and an antisense primer 5′-GGGCCCGGTGAGGACCGCAGCTTCTTCTGGCTAACGCT-3′) were used for PCR, whose products were subcloned into TA vectors. The 588-bp Mmp9 promoter region was cut out using ApaI and inserted into a pZsGreen1-1 vector (Clontech, Palo Alto, CA). This Mmp9 promoter-driven ZsGreen reporter expression plasmid was designated pZsGreen/Mmp9-promo.
Electroporation and reporter assay
Electroporation transfection was performed as described.1,45 For transient transfection, 3.5 × 105 cells were transfected with 10 μg of pMmp9-ZsGreen or pCMV-GFP (NEPA Gene) using the NEPA21 electroporator (NEPA Gene, Ichikawa, Japan). The condition of electroporation was optimized for Colon26 and LuM1. 1 Poring pulse condition was a total of two pulses at 150 V for 5 ms pulse length with 50 ms interval between the pulses, 10% decay rate with + polarity. The transfer pulse condition was a total of five pulses at 20 V for 50 ms pulse length with 50 ms interval, 40% decay rate with ± polarity. After electroporation, cells were immediately suspended into RPMI1640 10% FBS, seeded at a concentration of 1.75 × 104 cells/100 μL in a well of 96-well 2D culture plates, and then cultured for 24 h. Alternatively, cells were mixed with pCMV-EGFP without electroporation as a negative control (NC). NucBlue® Hoechst33342 (Thermo Fisher Scientific, Waltham, MA) was dropped into the medium, and cells were incubated for 1 h to count total cell number. The ArrayScan™ High Content Screening (HCS) system (Thermo Fisher Scientific) was used for measurement of fluorescence. 2 The rate of ZsGreen-positive cells (Mmp9 promoter-activated cells) and Mmp9 promoter activities was normalized with transfection efficiencies of pCMV-EGFP. Mmp9 promoter activity was evaluated by an average fluorescence intensity per μm2 ( = pixel) of ZsGreen-positive cells in a well using ArrayScan HCS system.
Cloning of stable transfectant cells and reporter assay
LuM1 cells grown until 60% confluency were transfected with pZsGreen/Mmp9-promo using FuGENE6 reagent (Roche, Basel, Switzerland) in a 35-mm culture dish. Cells were reseeded at day 2 post-transfection period. G418 (0.5 mg/mL) was added a day later. Culture medium was replaced by fresh ones containing G418 every 3 days. Cells that survived for 10 days were reseeded in 96-well plates at a concentration of 0.5 cell/well. ZsGreen-positive single colonies were picked.
For reporter assay to study the effects of TPA, DEX, and SB431542 using stable cells, 5.0 × 103 cells were seeded onto a 96-well 2D culture plate in RPMI1640 supplemented with 10% FBS for 24 h, then drugs were added, and cultured for 24 h. Mmp9 promoter activity was evaluated by an average fluorescence intensity per μm2 of all cells in a well. Experiments were performed with three or four biological replicates.
Gelatin zymography
As described, 10 protein samples (10 μg) were mixed with an sodium dodecyl sulfate (SDS) sample buffer and then applied to a 10% acrylamide gel containing copolymerized 0.1% gelatin.
Invasion assay
As described, 10 in vitro invasion assays were performed using Matrigel-coated culture systems (Becton Dickinson), respectively. Cells (2.5 × 104 cells) were transferred into the upper chambers of Transwell® 24-well plates (Corning). The invading cells on lower surfaces of filters were fixed at the 24-h postcell transfer period. Fluorescence images were taken using the FLoid® Cell Imaging Station (Thermo Fisher Scientific).
Syngeneic transplantation
All animal experiments were performed according to the guidelines for the care and use of laboratory animals approved by Okayama University and the Japanese Pharmacological Society (OKU-2015659). As described, 10 5 × 105 cells were transplanted subcutaneously on the side of abdominal walls of 6–7-week-old female BALB/c mice. The mice were fixed by perfusion fixation with 4% paraformaldehyde at day 20 and 40 after transplantation. The primary tumor and lungs were resected and observed under a fluorescent stereomicroscope SZX-12 (Olympus, Tokyo, Japan) at the Central Research Laboratory, Okayama University Medical School.
3D tumoroid reporter assay
To investigate the effects of LiCl on the stable clone, cells were seeded at a concentration of 5 × 103 cells/well in a 96-well NCP or a 96-well 2D culture plate, were precultured in RPMI1640 with 10% FBS for 2 days, then LiCl was added, and cultured in RPMI1640 with 2% or 10% FBS for 2 days. Alternatively, cells were cultured in the presence of LiCl in RPMI1640 with 2% or 10% FBS for 3 days. To investigate the effects of 5-FU, HC, ART, and AG490, cells were seeded at a concentration of 5 × 103 cells/well in a 96-well NCP and cultured in mTeSR1 for 3 days with or without drugs. For counting cell aggregates/tumoroids, fluorescent intensity and area (μm2 = pixel) of each aggregate per well were calculated using the ArrayScan HCS system. Fluorescent areas greater than 300 μm2 were counted as cell aggregates/tumoroids. Mmp9 promoter activity was evaluated by an average fluorescence intensity per μm2 of all cells in a well as described above. Experiments were performed with three or four biological replicates.
Western blot analysis
Cytoplasmic and nuclear lysates were prepared using NE-PER nuclear and cytoplasmic extraction reagents (Thermo Fisher Scientific). As described, 10 protein samples were loaded onto 8 or 10% polyacrylamide gel and transferred to a poly vinylidene di-fluoride membrane by using the semidry method. Blocking and antibody reactions were done in blocking buffer containing 5% skim milk (Wako, Osaka, Japan) in Tris-buffered saline containing 0.05% Tween 20 (TBS-T). The anti-β-catenin (1:1000; Cell Signaling Technology, Danvers, MA), anti-histone H3 (1:1000; Cell Signaling Technology), anti-phosphorylated STAT3 (p-STAT) (1:1000; Cell Signaling Technology), and HRP-conjugated anti-GAPDH antibodies (1:5000; Wako, Osaka, Japan) were used.
Genetic alteration and gene expression in human cancer cases
Genetic alterations and gene expression of MMP9 were analyzed in whole data (11,1117 samples, 31 types of cancer) available in the TCGA database using cBioPortal version January 9, 2019.
Statistics
Data are expressed as mean ± SD unless otherwise specified. Statistical significance was calculated using GraphPad Prism (La Jolla, CA). Three or more mean values were compared using one-way analysis of variance with the pairwise comparison by the Dunnett's method, while comparisons of two were done with an unpaired Student's t-test. A p < 0.05 was considered to indicate statistical significance.
Results
The promoter activity and expression of Mmp9 were increased in high metastatic colon cancer cells
We first conceptualized to establish a novel, convenient reporter system that is useful for evaluation of cancer abilities of tumorigenesis and invasion in vitro and in vivo (Fig. 1A). For this concept, we combined the Mmp9 promoter reporter assay and 3D tumoroid culture system, enabling us to measure abilities of invasion and tumorigenesis, respectively. For this purpose, we first examined whether mRNA levels and promoter activities of Mmp9 were different among the three different cancer cell lines: murine colon cancer cell line Colon26, its high metastatic subline LuM1, and nonmetastatic subline NM11. The endogenous mRNA level of Mmp9 in the high metastatic LuM1 was 2456-fold higher than that in the parental Colon26 and 52-fold higher than that in the nonmetastatic NM11 cell (Fig. 1B). This result was consistent with the high expression of MMP9 in human colon cancer.
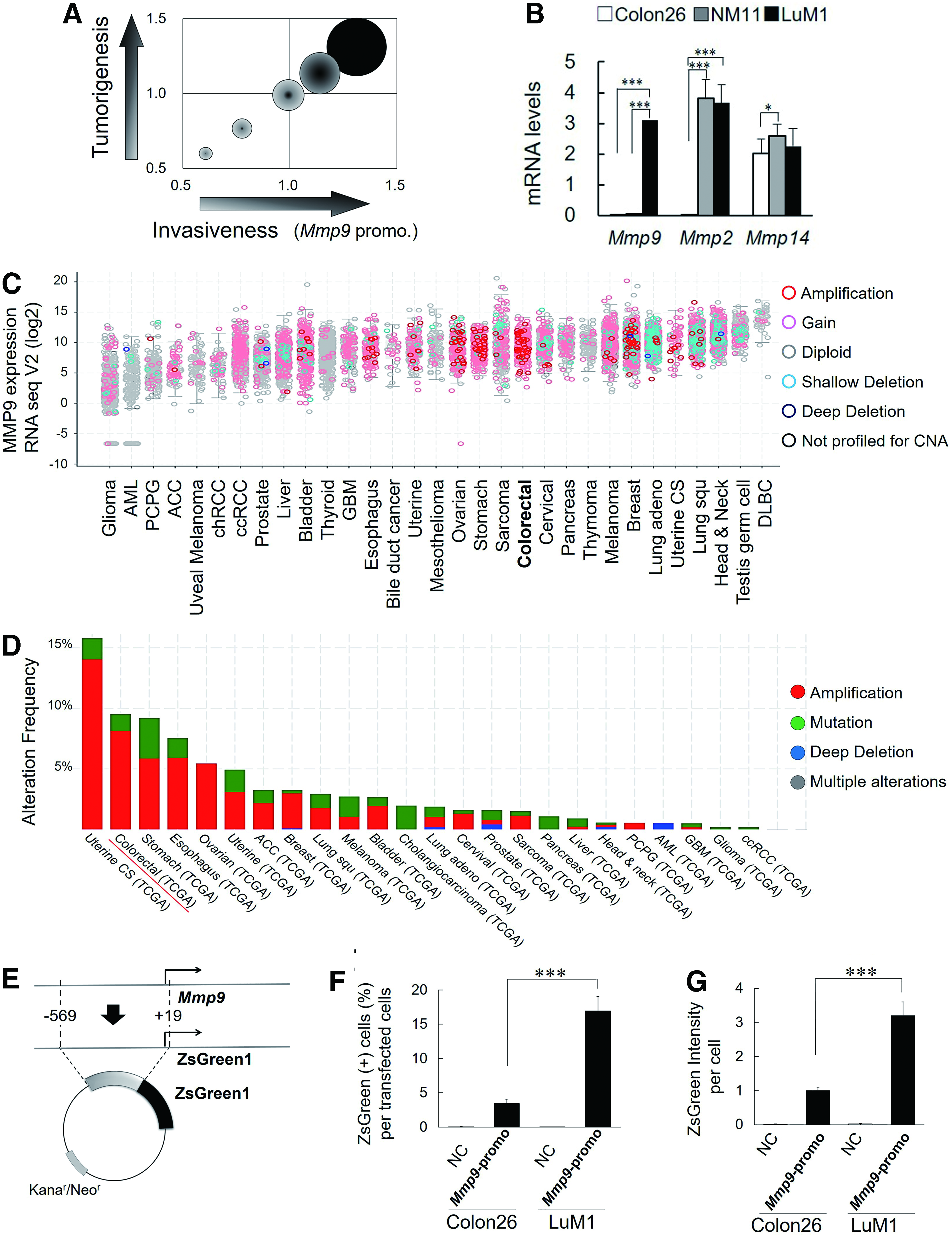
Increases in the promoter activity and mRNA expression of MMP9 in high metastatic colon cancer cells.
We next investigated the expression and genetic alteration of human MMP9 among various types of cancers, including colon cancer, using cBioPortal. The gain of MMP9 mRNA expression with genetic amplifications was frequently found in colorectal cancer cases compared with other types of cancer such as gliomas (Fig. 1C). Amplification of MMP9 gene in colorectal cancer was found in 8% of a total of 640 colorectal cancer cases (Fig. 1D). It was thus indicated that MMP9 is expressed at high levels in human and mouse colon cancer.
To investigate a mechanism underlying the high expression of MMP9, we next compared proximal promoter sequences between human and mouse. Of note, binding motifs of TCF/LEF, NF-κB, and GR, as well as a TPA response element (TRE), were conserved between human and mouse MMP9 promoter regions (Supplementary Fig. S1). For the establishment of a novel reporter system useful for syngeneic transplantation and metastasis in mice, we cloned cDNA corresponding to the murine Mmp9 promoter region between −569 and +19, which contains multiple transcription factor-binding sites, into ZsGreen fluorescence reporter plasmid (Fig. 1E). The 588-bp Mmp9 promoter was activated in 17% of LuM1 cells and 3.5% of Colon26 cells (Fig. 1F) (data were normalized with transfection efficiencies to each line shown in Supplementary Fig. S2), consistent with the different levels of Mmp9 mRNA between these cell lines. Mmp9 promoter activity per cell was threefold higher in LuM1 population than that in Colon26 population (Fig. 1G and Supplementary Fig. S2). Interestingly, the mRNA levels and promoter activities of Mmp9 in LuM1 versus Colon26 were not exactly coincident each other, suggesting that expression of MMP9 in cancer could be regulated at both transcriptional and posttranscriptional levels.
These data suggested that a potential regulatory mechanism is underlying the increased transcription of MMP9, specifically in high metastatic colon cancer.
Cloning of fluorescent reporter cells that enable monitoring of activities of MMP9/gelatinase, invasion, and metastasis
Recent studies showed heterogeneity of cancer cells, including CIC/CSC, in a single tumor. We therefore next cloned stable fluorescent cells that exert powerful promoter activities of Mmp9, gelatinolytic activities, invasiveness, and metastatic abilities. LuM1 sublines transfected with the Mmp9 promoter (designated LuM1/m9) #1 and #4 robustly expressed ZsGreen reporter that was driven by the Mmp9 promoter compared with either sublines #2 and #3 or the parental LuM1 cells (Fig. 2A). Gelatinolytic activities of 90-kDa gelatinase B (Gel. B, MMP9) and 60-kDa gelatinase A (Gel. A, MMP2) were higher in culture supernatants of LuM1/m9 sublines #1 and #4 than that of low metastatic Colon26 (Fig. 2B). Gelatinase activity of MMP9 of LuM1/m9 #4 was higher than that of the subline #1 (Fig. 2B, arrow). The LuM1/m9 #4 kept powerful fluorescence even after passages, whereas other sublines gradually lost their fluorescence. We therefore next examined whether the LuM1/m9 #4 cells possessed invasion abilities in vitro. We detected that the fluorescence of LuM1/m9 #4 cells invaded through matrix and pore at comparable levels with the nonfluorescent parental LuM1 (Fig. 2C).
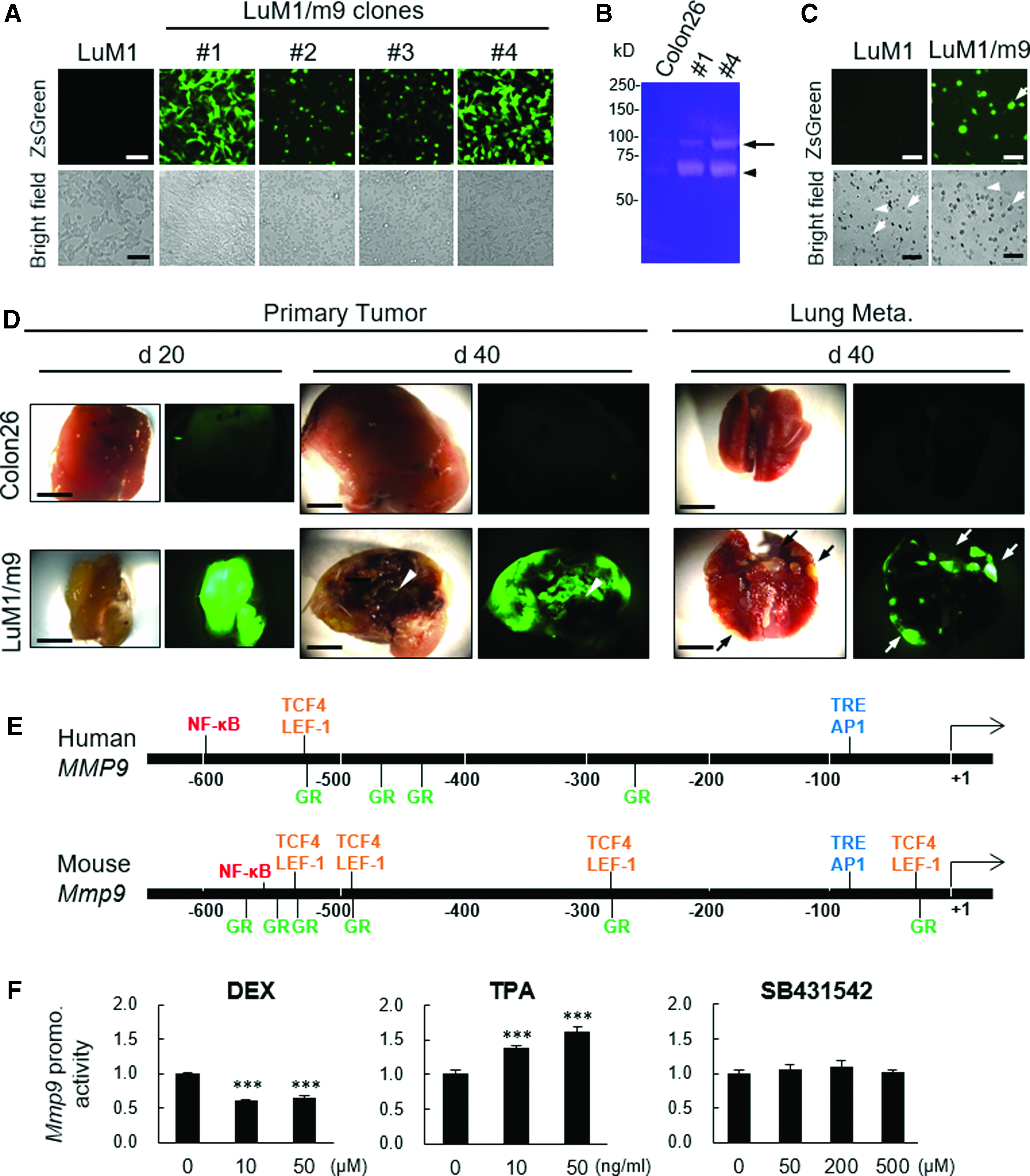
Stable reporter cells that monitor activities of MMP9/gelatinase, invasion, and metastasis. LuM1 cells were transfected with pZsGreen/Mmp9-promo and stable sublines were established.
We next examined whether LuM1/m9 #4 could have abilities of tumorigenesis and metastasis with detectable reporter fluorescence in vivo, using syngeneic transplant experiments. Subcutaneous primary tumors at day 20 and 40, as well as lung metastatic secondary tumors at day 40, showed powerful ZsGreen fluorescence. The tumor at day 20 was filled with ZsGreen-positive live cells (Fig. 2D, bottom left). In contrast, the nonfluorescent “necrotic” area was found in tumor at day 40 (Fig. 2D, bottom center). Lung metastatic secondary tumor nodules were found with powerful fluorescence at day 40 postinjection period (Fig. 2D, bottom right). We also examined tumors formed by the injection of nonfluorescent Colon26 cells as an NC, which enabled confirmation of LuM1/m9 tumor fluorescence positivity derived from the Mmp9 promoter-driven reporter gene activation.
Thus, we established a novel fluorescent reporter system for monitoring invasiveness, tumor growth, and metastasis.
Regulation of the Mmp9 promoter by DEX and TPA
It was suggested that MMP9 high expression and its powerful promoter activity are biomarkers of cancer activities in terms of invasiveness and metastatic potential. We therefore hypothesized that regulatory signaling for the Mmp9 promoter plays a key role in controlling cancer activities. By promoter analysis, we found that many transcription factor-binding sites were conserved between the proximal promoter sequences in human and mouse, including GR, NF-κB, and AP1, whose binding site was shown to be a TRE (Fig. 2E). To examine whether NF-κB and GR signaling and TPA regulate the Mmp9 promoter, we investigated whether TPA and DEX could alter the Mmp9 promoter activity in metastatic reporter cells. We used these materials because DEX is a type of corticosteroids that antagonize GR and used as an SAID 38 and TPA is a carcinogen that activates MMP9 gene via activation of multiple signaling pathways, including NF-κB, Raf/MEK/ERK, p38 MAPK, and RAC.46,47 DEX significantly decreased the activity of the Mmp9 promoter to ∼60% of the control at the concentrations of 10 and 50 μM (Fig. 2F, left). On the contrary, TPA significantly increased the activity of Mmp9 promoter in the metastatic cells at concentrations of 10 and 50 ng/mL (Fig. 2F, center). We also examined the transforming growth factor β receptor inhibitor SB431542 as an NC, since no SMAD binding sequence was found in the Mmp9 promoter region. As expected, SB431542 did not alter the promoter activity (Fig. 2F, right).
These results indicated that GR and NF-κB signaling is involved in Mmp9 expression in metastatic cancer cells, although another signal might also regulate the Mmp9 promoter, as investigated in the following studies.
β-catenin stability and 3D tumor growth enhance the Mmp9 promoter activity
The intratumoral milieu is being hypoxic when enlarged, and thus, the hypoxia-inducible factor 1 (HIF1) activates MMP genes.48–50 In addition, we recently reported that both hypoxic organoid/tumoroid formation and stemness-enhancing medium mTeSR1 increased stem cell properties.1,2 The mTeSR1 medium contains LiCl that activates β-catenin signaling by inhibiting GSK3.41,51 Therefore, we hypothesized that Wnt/β-catenin signaling for stem cell renewal could also mediate high expression of MMP9 during tumorigenesis. To test this hypothesis, we examined whether the 3D tumoroid-inducing milieu and LiCl could increase tumorigenesis as well as the Mmp9 promoter activity. LuM1/m9 #4 was seeded onto 2D culture plate or 3D NCP, and cultured in RPMI 1640 with 10% FBS (serum) or mTeSR1 (stem). After 2 days of incubation, the size of tumoroid and Mmp9 promoter-driven reporter fluorescence were measured using the ArrayScan HCS system. Tumoroids were enlarged with fluorescence in 3D tumoroid-inducing NCPs (3D-serum) and mTeSR1 medium (2D stem) and furthermore enlarged by their combination (3D stem) (Fig. 3A, B). Simultaneously, Mmp9 promoter activity was increased in 3D tumoroid-inducing culture (3D-serum) and mTeSR1 medium (2D stem) and further increased by their combination (3D stem) (Fig. 3C).
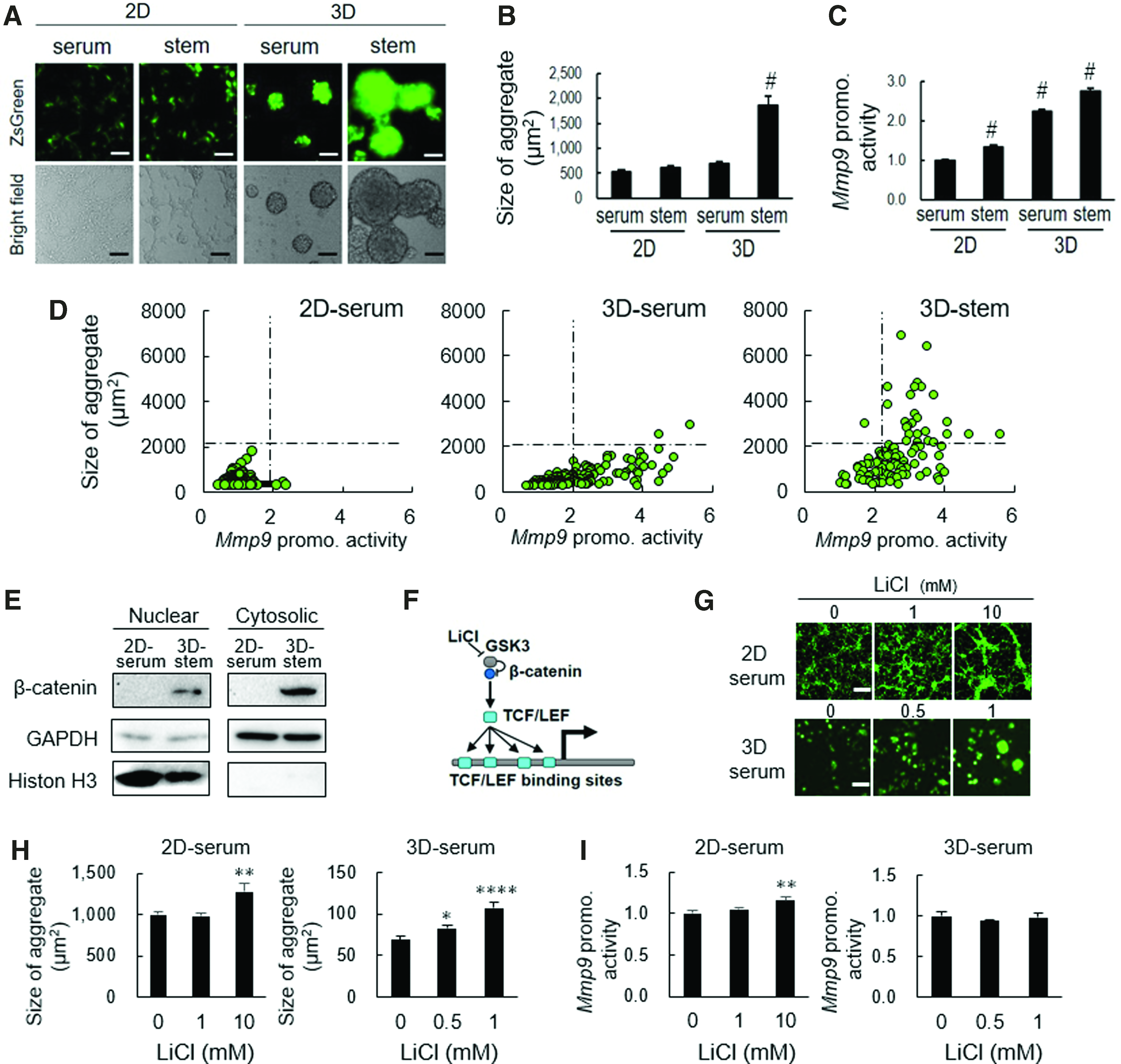
The stability of β-catenin and 3D tumor growth enhanced the Mmp9 promoter activity.
We next examined the scatter plot analysis of every single aggregate in terms of size (vertical axis) and Mmp9 promoter activities (horizontal axis). The 3D culture on NCP (3D-serum) markedly increased Mmp9 promoter activity as well as a tumoroid formation (Fig. 3D, compare graphs on the left and center). The add-on of mTeSR1 medium to the 3D culture system (3D stem) markedly increased the size of tumoroid (Fig. 3D, compare graphs on the right and center), consistent with the data shown in Figure 3A–C. Thus, these data suggested a synergistic effect of 3D culture and stemness-enhancing signaling on the tumoroid formation.
As the mTeSR1 medium contains LiCl, a stimulator of β-catenin signaling, we next examined β-catenin in the tumoroids and effects of LiCl on cell aggregation and Mmp9 promoter activities. β-catenin was found in both nuclear and cytoplasmic fractions of the 3D stem tumoroids of the LuM1/m9 cells, whereas this factor was not detected in the 2D serum culture condition (Fig. 3E), suggesting that LiCl in mTeSR1 medium might contribute to the stabilization of β-catenin through inhibition of GSK3β, an inhibitory factor of β-catenin (Fig. 3F). It has been shown that after stabilization, β-catenin translocates to nuclei, where this factor binds to TCF/LEF, 51 whose consensus binding sequences were found in the Mmp9 promoter sequence (Figs. 2E and 3F). The size of cell aggregates was increased by LiCl in either 2D serum or 3D serum conditions (Fig. 3G, H, and Supplementary Fig. S3), suggesting that the stabilized β-catenin/TCF complex might activate its target cyclin D gene that promotes cell proliferation. The Mmp9 promoter activity was increased by the administration of LiCl in the 2D serum condition (Fig. 3I), although the activity was not altered in the 3D stem tumoroid (Supplementary Fig. S3), suggesting that the Mmp9 promoter might be occupied by hypoxia-HIF1 signaling in such large tumoroids. These findings suggested that the stabilized β-catenin/MMP9 regulatory axis was activated during tumor growth (while the intratumoral hypoxia-MMP9 regulatory axis could be simultaneously activated when the tumor was enlarged). It was also indicated that the combination of NCP-based 3D culture and mTeSR1 stemness-enhancing medium or LiCl is useful for the development of tumoroid studies.
ART inhibited tumorigenesis and the Mmp9 promoter of metastatic colon cancer cells in vitro
Our reporter system has been demonstrated to be useful for monitoring cancer invasiveness, metastasis, 3D tumorigenesis in vitro, and key signalings such as NF-κB and Wnt/β-catenin. Therefore, it is supposed that this reporter system could also be suitable and useful for drug screening with an advantage of 3D tumoroid formation within the culture medium. We examined the effects of ART, 5-FU, and HC on in vitro tumorigenesis using this 3D reporter system as a pilot study. LuM1/m9 #4 was seeded and simultaneously treated with 5-FU, HC, or ART within mTeSR1 stem cell medium in NCPs. After 3 days of incubation, the size of tumoroid and Mmp9 promoter-driven reporter fluorescence were measured using the ArrayScan HCS system (Fig. 4A). The average size of organoid and the average activity of the Mmp9 promoter in tumoroid were depicted as a scatter plot. 5-FU significantly inhibited tumorigenesis to ∼50% of the control but did not decrease the Mmp9 promoter activity at a concentration of 10 μM (Fig. 4B, bottom right; Supplementary Fig. S4). ART significantly inhibited both the Mmp9 promoter activity and tumorigenesis to ∼85% and 40% of the control, respectively, at a concentration of 20 μM (Fig. 4B, bottom left; Supplementary Fig. S4). HC (2 μM) inhibited the Mmp9 promoter activity to ∼20% of the control but did not inhibit in vitro tumorigenesis (Fig. 4B, top left; Supplementary Fig. S4). Lower concentrations of ART (2 or 4 μM) and 5-FU (1 μM) did not alter either the size of tumoroids or the Mmp9 promoter activities (Supplementary Fig. S4).

ART inhibited tumorigenesis and the Mmp9 promoter of metastatic colon cancer cells in vitro.
We next depicted scatter plots of all cell aggregates per well in terms of their size (vertical axis) and Mmp9 promoter activities (horizontal axis). It was shown that 5-FU and ART inhibited in vitro tumorigenesis compared with control or HC (Fig. 4C). HC and ART tended to attenuate the Mmp9 promoter activities compared with the control or 5-FU, although tumoroids with high Mmp9 promoter activity were still present even after treatment with HC or ART (Fig. 4C).
Since many STAT binding sites were found in the Mmp9 promoter region, as shown in Supplementary Figure S1, we examined STAT inhibitor AG490 and detection of p-STAT, often active in stem cells. However, AG490 altered neither the in vitro tumorigenesis nor the Mmp9 promoter activity. The p-STAT3 was barely detectable in the 2D serum and 3D stem conditions (Supplementary Fig. S5).
Thus, these findings indicate that (i) ART inhibits tumorigenesis and Mmp9 promoter potentially through inhibition of β-catenin, (ii) HC inhibits Mmp9 promoter by a potential mechanism underlying HC/DEX-dependent repression of GR/NF-κB complex, and (iii) 5-FU inhibits tumorigenesis.
Discussion
Our studies established a novel reporter system that enables to monitor tumorigenesis in vivo and in vitro, metastasis, invasiveness, and is useful for studies of signaling, transcription, and drugs that alter the promoter activity of MMP9, which is one of the key biomarkers as well as a causal factor in cancer progression. We showed that the 588-bp Mmp9 promoter-driven reporter cells (LuM1/m9) were profoundly gelatinolytic and invasive (Figs. 1 and 2). This Mmp9 promoter reporter monitored cancer cells at both primary and secondary tumors in vivo (Fig. 2D). The quantitative analysis of the tumoroid size and the reporter activities enabled efficient pharmaceutical studies (Figs. 3 and 4). Although numerous factors and signaling pathways have been shown to involve CIC/CSC,2,25,52–56 to elucidate key factors that could promote tumoroid growth with enhanced stemness and increased Mmp9 promoter activity, we analyzed the MMP9 promoter sequence and then found that a TCF/LEF-binding site was conserved between human and mouse (Fig. 2E). It has been shown that Wnt/β-catenin/TCF signaling is essential for self-renewal of CIC/CSC. 29 Our present study showed that β-catenin was stabilized in the 3D stem tumoroid culture condition, presumably through inhibition of GSK3β by LiCl supplemented in mTeSR1. Such stabilization of β-catenin could trigger nuclear translocation of this factor (Fig. 3E). To elucidate whether the β-catenin-TCF/LEF signal is crucial for activation of the Mmp9 promoter, we used LiCl, which can activate β-catenin-TCF/LEF signaling. Indeed, LiCl exposure increased Mmp9 promoter activity as well as cell aggregates. Such effects of LiCl on the growth of cell aggregates and on activation of the Mmp9 promoter were observed in early stages of tumorigenesis, in which β-catenin/TCF might activate its target gene cyclin D1. 33 Thus, our data touch on the Wnt/β-catenin regulation of MMP9 in colon cancer. This is consistent with a recent report that Wnt3a expression is associated with MMP9 expression in the primary tumor and metastatic site in recurrent or stage IV colorectal cancer. 18 Another study also reported the β-catenin regulation of MMP9 in melanoma using a β-catenin inhibitor lycorine. 57 However, in the present study, LiCl was ineffective when the tumoroid grew larger, which mimics the later stage of tumors. The interior of tumors is hypoxic, in which HIF1 activates MMP9 gene and other oncogenes.1,2,52,58–61
Therefore, targeting Wnt/β-catenin signaling might be effective only at the early stage of tumorigenesis, whereas targeting hypoxia-HIF1 signaling might be effective for enlarged hypoxic tumors at the later stage. Regardless of the activities of β-catenin/TCF signaling or hypoxia-HIF1 signaling, we showed that siRNA-targeted depletion of MMP9 and MMP3 was effective to inhibit tumorigenesis and metastasis. 10 Moreover, not only extracellular proteolytic MMPs but also intracellular MMPs could contribute to cancer transcriptional regulation and migration.10,20–22
Our studies also indicated that 5-FU was effective to decrease tumorigenesis of the MMP9 high colon cancer cells. 5-FU resistance has been shown in colon cancer and its mechanism is still unclear. 62 Nevertheless, a recent study reported that overexpression of MMP9 and Rab1B predicts poor survival and good response to chemotherapy in patients with colorectal cancer. 17 This report is consistent with our finding that 5-FU was effective to decrease tumorigenesis of the MMP9 high colon cancer cells (Fig. 4). In clinical cases, 5-FU has been applied mostly in combination with other substances. 63 Therefore, a combination of therapeutics, found to be effective, may solve resistance and recurrence of cancer. Our pilot study also revealed some effectiveness of ART on Mmp9 promoter activity and tumorigenesis (Fig. 4), in which β-catenin was stabilized. It has been shown that the anticancer activity of ART correlated with the inhibition of hyperactive Wnt/β-catenin signaling pathway.64,65 Moreover, it was shown that ART inhibited the expression of several MMPs, including MMP2 and MMP7 mRNA/proteins. 66 These studies were consistent with our findings that ART inhibited both the Mmp9 promoter and in vitro tumor formation, in which β-catenin-TCF/LEF signaling was active. Therefore, ART is useful as an anticancer medication that inhibits tumorigenesis, invasiveness, and recurrence, and resistance of cancer through targeting the Wnt/β-catenin/MMP regulatory axis.
Due to practical and ethical limitations in human experimentation, animal models have been essential in cancer research. However, the average rate of successful translation from animal models to clinical cancer trials is less than 8%. 67 Animal models can provide a key source of in vivo information, but alternative translational methods have emerged that may eventually replace the connection between in vitro studies and clinical applications. Human tissue models in cancer research, including patient-derived xenograft mice, are widely used to address questions in cancer research. 68 The latest animal models of human colon cancer have opened up new doors for continuing cancer research for not only understanding the colon cancer pathogenesis but also aiding in the development of newer chemotherapeutic drugs as they mimic the human disease closely. 69 However, each reported model has some limitations. A limitation using human cancer cells is that researchers ought to transplant the human-derived cancer cells to immunodeficient mice such as nude mice or severe combined immunodeficient mice, in which studies on tumor immunology are limited. Our reporter system was established using mouse metastatic colon cancer cells that are useful for syngeneic transplantation to immunologically normal mice, in which researchers could analyze tumor milieu and immunology. Moreover, we have developed the 3D tumoroid system using mouse-derived cancer cells as well as human-derived cells.1,2 Growing cells as 3D models are more analogous to their existence in vivo, for example, akin to a tumor, and being able to be cocultured with other cells and cellular components that naturally occur in their milieu may be more clinically relevant. 9 Therefore, the LuM1/M9 reporter system established with the BALB/c mouse-derived cancer cells in the present study could be useful for the 3D tumoroid model cocultured with other types of cells such as immune cells and for syngeneic transplantation to investigate the tumor microenvironment and tumor immunology.
In conclusion, this novel fluorescent reporter system for dual monitoring of tumorigenesis and Mmp9 promoter activity in vitro and in vivo is useful for studies of invasiveness, metastasis, signaling pathways, pharmacology, and drug discovery. It was also demonstrated that ART inhibited in vitro 3D tumorigenesis and Mmp9 promoter activity, potentially via inhibiting Wnt/β-catenin signaling.
Footnotes
Acknowledgments
This article is dedicated to the memory of one of the coauthors, Professor K.K., who passed away on May 29, 2016. This work was supported by JSPS KAKENHI, grant no. 17K11643 (C.S., T.E.), JP17K11642 (T.E.), JP17K17895 (Y.O.), JP17K11669 (K.O., C.S., T.E.), JP16K11863 (K.O.), JP18K09789-KN (T.E.), JP26293067 (K.K.), and JP26670815 (K.K.) and by the SUZUKEN memorial foundation (T.E.).
Authors' Contributions
K.K., C.S., and T.E. conceptualized and designed the study. C.S., K.K., N.S., and T.E. devised the methodology. K.K., T.E., M.T., N.S., C.S., and K.O. prepared the resources. C.S., Y.O., N.S., K.O., M.I., R.K., and Y.H. carried out the experimentation. C.S. carried out formal analysis. T.E. and C.S. interpreted data. T.E. and C.S. wrote the article. T.E., C.S., and K.O. revised and edited the article. All authors reviewed the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.