
Abstract
Large soft-tissue defects are challenging to reconstruct surgically. Expansion of soft tissue using an external volume expansion (EVE) device is a noninvasive method to improve such reconstruction; however, the underlying mechanism is unclear. In this study, we created fat flaps in Sprague-Dawley rats, applied an external force of 3 or 6 kPa using an EVE device, and investigated the migration and differentiation of adipose-derived stem/progenitor cells (ASCs). In addition, we performed finite element analysis to explore the stiffness of adipose tissue. An external force of 3 kPa promoted the migration and adipogenic differentiation of ASCs. By comparison, an external force of 6 kPa had a larger effect on migration of ASCs, but a smaller effect on adipogenic differentiation of ASCs. External force affected adipose tissue stiffness. In conclusion, external force generated by an EVE device increases the stiffness of adipose tissue, which influences the migration and differentiation of ASCs. The size of the external force can be altered according to the tissue stiffness required at particular time points to promote long-term adipose tissue regeneration.
Impact Statement
Stem cell therapy in clinic mostly requires the addition of exogenous stem cells, therefore the safety and controllability is always defective. In this study, the external force of external volume expansion regulates adipose-derived stem/progenitor cells (ASCs) migration and differentiation through tissue stiffness. Using tissue engineering without exogenous ASCs can promote long-term adipose tissue regeneration. The findings of this study provide theoretical support for clinical tissue engineering applications and improvements in stem cell therapy.
Introduction
Large soft-tissue defects caused by trauma or surgery are challenging to reconstruct surgically. Currently, reconstruction of these defects are always using allogeneic/synthetic fillers, fat grafts or autologous flaps.1–4 However, a recently developed external soft-tissue expansion system, external volume expansion (EVE) device, can achieve better results as a nonsurgical and noninvasive method. 5 The system stimulates the growth of breast tissue by applying distracting force and was first used in breast augmentation in 1999.6–9 However, its underlying mechanism is unclear.
To investigate this, we previously applied an EVE device in animals and found that it induces adipose tissue regeneration.3,10,11 Adipose tissue regeneration is dependent on tissue vascularization and adipogenic differentiation of adipose-derived stem/progenitor cells (ASCs). Therefore, the source, number, and differentiation capacity of ASCs are important for adipose tissue regeneration.12–14 In particular, migration and differentiation of ASCs are associated with long-term tissue regeneration.11,12
Cell migration is sensitive to environmental stiffness and is regulated by many factors. For example, external force induces cell migration. 15 Cells migrate from areas of low stiffness to areas of high stiffness.16,17 Some studies suggested that stiffness has a greater influence on cell migration than proteins.18,19 Tissue stiffness also regulates cell differentiation.20–23 External force generated by an EVE promotes tissue regeneration.10,11 Thus, external force may alter adipose tissue stiffness and thereby promote the migration and differentiation of ASCs.
We hypothesized that external force generated by an EVE changes the stiffness of adipose tissue and thereby regulates the migration and differentiation ASCs. To investigate this, we used an EVE device in Sprague-Dawley rats. We applied a high or low external force and explored the relationship between adipose tissue stiffness and external force. Rats were transplanted with CM-DiI (CiI)-labeled ASCs, and the migration and number of these cells were investigated. In addition, we investigated the differentiation capacity of ASCs.
Materials and Methods
Animals
All experiments procedures were approved by the Animal Ethics Committee of Nanfang Hospital and were performed in accordance with the guidelines of the National Health and Medical Research Council of China. One hundred male Sprague-Dawley rats weighing 350 ± 20 g were housed under a 12-h day/night cycle in SPF conditions, provided with standard food and water ad libitum.
EVE animal model
As previously described, 10 EVE animal model was established. In brief, 98 male Sprague-Dawley rats were anesthetized by isoflurane (30 mg/kg body weight). Then, exposed the fat flap through an incision on the right inguinal region, and transferred the fat flap to the middle of the abdomen after having separated it from the surrounding tissue. In the end, sutured the incision and marked the contour of the flap.
Seventy male Sprague-Dawley rats were randomly divided into two groups, namely, the High-EVE and Low-EVE groups, each of which contained 35 rats. A dome-shaped acrylonitrile-styrene copolymer device, 3 cm in diameter, 15 mL internal volume, was connected to a suction pump (low-vacuum suction unit DYX-1A; SMAF, Shanghai, P. R. China) via a flexible rubber tube. At 12 h after surgery, a suction of a constant pressure of 6 kPa (High-EVE group) or 3 kPa (Low-EVE group) was applied to the transferred flap (Fig. 1). Adipose tissue was collected from seven rats per group on days 0, 1, 3, 5, 7, and 9, fixed in 4% paraformaldehyde for 24 h, and stored at −80°C for protein expression analysis.

Schematic of the experimental setup. Color images are available online.
Isolation and culture of ASCs
ASCs were isolated from inguinal adipose tissue of two healthy rats according to following protocols. Briefly, fat was obtained from bilateral inguinal fat pads, minced, and digested with phosphate-buffered saline containing 0.075% collagenase on a shaker at 37°C for 40 min. Digested tissue was centrifuged at 180 g for 5 min and the large fragments were filtered off. The cellular pellet (stromal vascular fraction [SVF]) was resuspended in erythrocyte lysis buffer and centrifuged at 180 g for 5 min. The SVF was partially plated and cultured in standard culture medium. Cells were used at passage 3–5.
Transplantation of CiI-labeled ASCs
To investigate the migration of ASCs upon application of the EVE, 28 male Sprague-Dawley rats underwent flap transfer and were attached to the suction pump with a pressure of 6 kPa (High-EVE group) or 3 kPa (Low-EVE group) for 12 h. Thereafter, the rats were transplanted with fluorescently labeled ASCs. These cells were generated by labeling ASCs with 50 μM CiI (Molecular Probes, Eugene, OR) at 37°C for 2 min and then incubating them on ice for 15 min before transplantation.
Histological examination
Tissue samples were fixed in 4% paraformaldehyde, dehydrated, embedded in paraffin, and stained with hematoxylin-eosin. Sectioned the tissue blocks, and examined under an Olympus BX51 microscope. Images were acquired using an Olympus DP71 digital camera.
Immunofluorescence analysis
Samples were stained with anti-CD34 (1:200; Santa Cruz Biotechnology) and anti-proliferating cell nuclear antigen (PCNA) (1:200; Abcam) primary antibodies, washed, and then labeled with donkey anti-goat or goat anti-rabbit immunoglobulin G secondary antibodies (Santa Cruz Biotechnology). DAPI (Sigma, St. Louis, MO) were used to stain nuclei. The number of ASCs (CD34+PCNA+) in at least seven fields per sample in a double-blinded fashion were counted by two authors under a confocal microscope (Leica TCS SP2; Leica Microsystems GmbH, Wetzlar, Germany).
Western blotting
Total cell lysates of adipose samples were prepared using M-PER Mammalian Protein Extraction Reagent (ThermoFisher Scientific). A BCA protein assay (ThermoFisher Scientific) was used to estimate the concentration of the protein. After separation by sodium dodecyl sulfate polyacrylamide gel electrophoresis using a NuPAGE electrophoresis system, protein extracts were transferred to immobilon polyvinylidene difluoride membranes (Millipore, Billerica, MA). Then, blocked membranes in 5% milk and immunoblotted them with the following primary antibodies: anti-CDC42 (1:1000; Cell Signaling Technology), anti-RhoA (1:1000; Cell Signaling Technology), anti-dynamin (1:1000; Cell Signaling Technology), peroxisome proliferators-activated receptors--γ (PPAR-γ) (1:200; Abcam), anti-cytosine-cytosine-adenosine-adenosine-thymidine enhancer binding proteins (C/EBP) (1:1000; Abcam), and anti-fatty acid binding proteins (AP2) (1:1000; Abcam). Thereafter, membranes were incubated with secondary antibodies. A WesternBreeze Chemiluminescent Detection Kit (ThermoFisher Scientific) was used to detect signals. GAPDH and tubulin served as internal controls.
Stiffness analysis
A BOSE ElectroForce® load testing device was used to analyze the stiffness. The load cell used to measure forces ranged from 0 to 225 N. The device's electromagnetic actuator allows an axial travel of 6 mm. Therefore, a preload was applied to remove slack from tissues and induce elongation. Each tissue sample was clamped and then a preload (25–30 N) was applied followed by ramp loading. The preload was selected based on preliminary testing. The ramp loading was set to 8 mm/s, and samples were expected to rupture during the application of this loading. The highest force achieved before the sutures tore was electronically recorded for each sample. Finally, the device calculated the tissue stiffness.
Finite element analysis
After analysis of tissue stiffness, simulations were performed to investigate how application of external force using the EVE device affected each tissue. Finite element (FE) simulation of rat adipose tissue was performed in microscopic conditions. The images were segmented and an FE mesh was generated. The model was solved using ABAQUS FEA (Dassault Systèmes Simulia Corp., Providence, RI) to determine the displacement field and tissue stiffness.
Statistical analysis
Complete randomization was used for grouping. Random numbers were generated using SAS 9.4 software (SAS Institute, Inc., Cary, NC) and then grouped according to a random number generator. All data were analyzed with IBM SPSS Version 20.0 software (IBM Corp., Armonk, NY). Results were presented as mean ± standard deviations. Comparisons between multiple time points were assessed using two-way analysis of variance. Comparisons between two groups at a single time point were assessed using independent Student's t-test. A two-tailed p-value less than 0.05 was considered statistically significant.
Results
Analysis of morphometry and sample volume
Fat flaps were hard in the High-EVE group from day 3 onward, but remained soft in the Low-EVE group. However, the appearance of fat flaps did not markedly differ between the two groups (Fig. 2A).
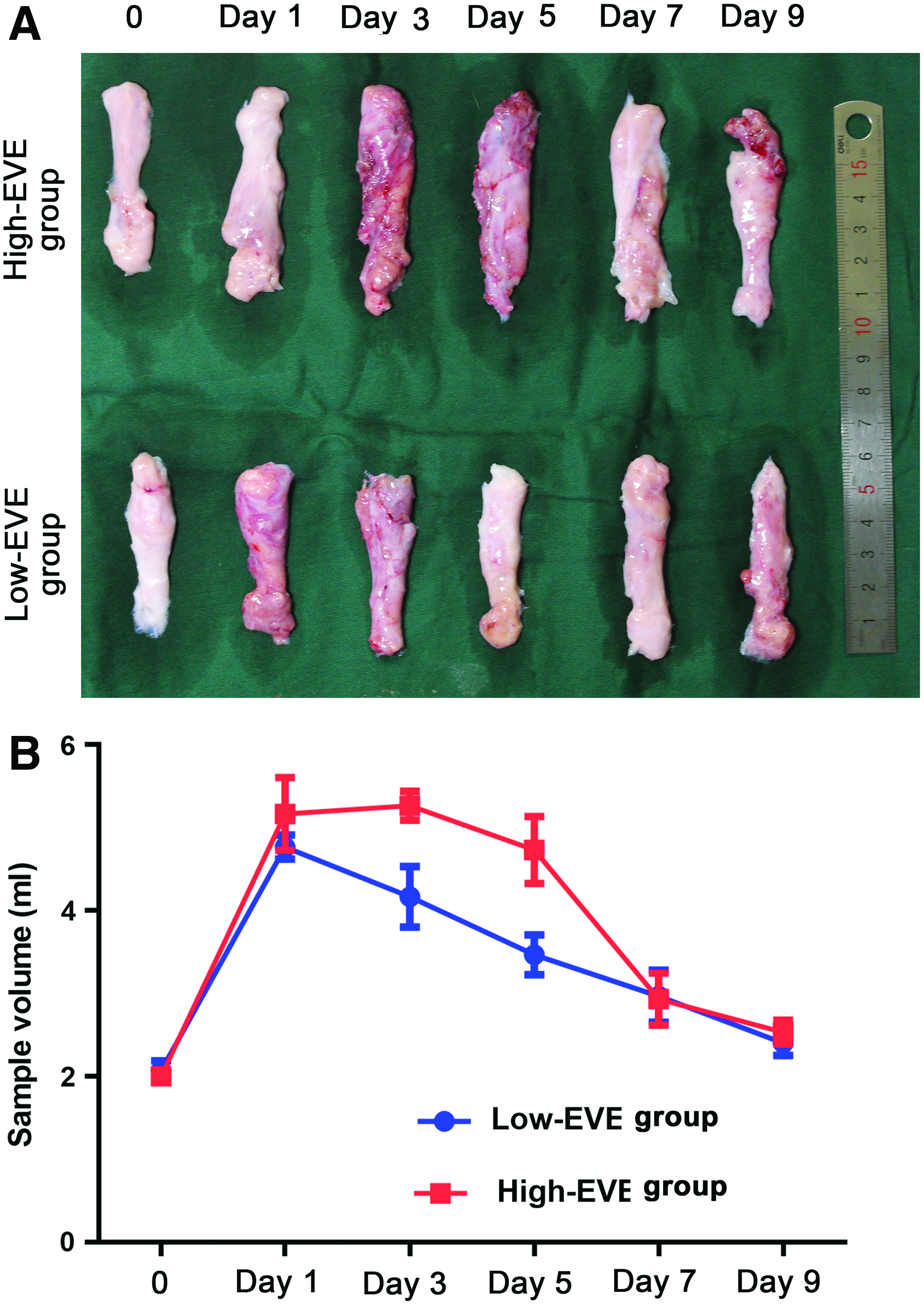
Gross morphology and volume of fat flaps.
The adipose tissue volume was increased on day 1 in both groups, began to decrease on day 5 in the High-EVE group and on day 1 in the Low-EVE group, and did not significantly differ between the two groups on days 7–9 (Fig. 2B).
Histologic evaluation
Histological analyses on day 9 revealed that fat flaps had an abnormal structure with a high degree of fibrosis in the High-EVE group compared with normal fat flaps. Although fat flaps also had an abnormal structure in the Low-EVE group, the degree of fibrosis was lower than in the High-EVE group (Fig. 3).

Histological analysis of normal fat flaps and those in the High-EVE and Low-EVE groups on day 9. Color images are available online.
Migration of ASCs
Immunofluorescence analysis detected many CD34+ stem cells on day 1 in the High-EVE group. A high number of these cells were also detected on day 3, but some were PCNA+ (Fig. 4A). The number of CD34+ stem cells remained high on day 3 in the Low-EVE group (Fig. 4A). Quantitative analysis showed that the number of CD34+PCNA− cells peaked on day 1, decreased on day 3, and remained stable thereafter in the High-EVE group (Fig. 4B). Meanwhile, the number of CD34+/PCNA− cells peaked on day 3, decreased on day 5, and remained stable thereafter in the Low-EVE group (Fig. 4B).
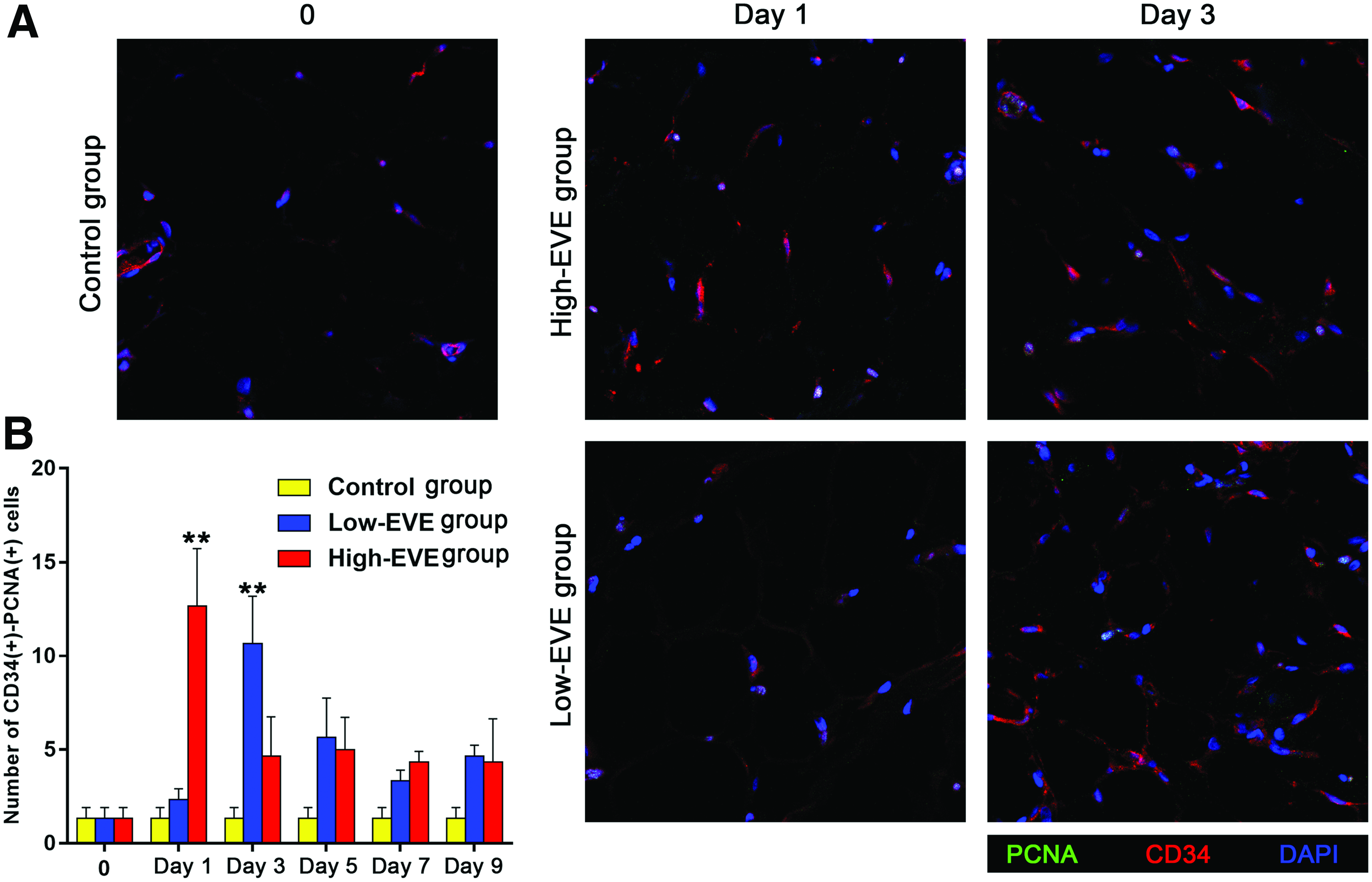
Immunofluorescence analysis of CD34 and PCNA expression in fat flaps.
Rats were transplanted with CiI-labeled ASCs at day 1 and 3 respectively (Fig. 5A). Many more CiI+ cells were observed in the High-EVE group than in the Low-EVE group on day 1 (Fig. 5B). However, many more CiI+ cells were observed in the Low-EVE group than in the High-EVE group on day 3 (Fig. 5B). Quantitative analysis revealed that the number of CiI+ cells in the High-EVE group was significantly higher than in the Low-EVE group on day 1, but was significantly higher in the Low-EVE group than in the High-EVE group on day 3 (Fig. 5C). However, the number of CiI+ cells was lower in the Low-EVE group on day 3 than in the High-EVE group on day 1 (Fig. 5C).

Labeling and tracing of ASCs.
Expression of cell migration-associated proteins
Expression of cell migration-associated proteins is shown in Figure 6A. Western blotting revealed that CDC42 expression in the High-EVE group was significantly higher than that in the Low-EVE group on day 1, but was lower than that in the Low-EVE group on day 5 and much lower than that in the Low-EVE group on day 9 (Fig. 6B). CDC42 expression was high in the Low-EVE group on day 5–9. RhoA expression decreased on day 3 in the High-EVE group and on day 5 in the Low-EVE group (Fig. 6B). Expression of RhoA peaked on day 9 in the Low-EVE group and then decreased. Dynamin expression was higher in the High-EVE group than in the Low-EVE group at all time points (Fig. 6B). Expression of cell migration-associated proteins was delayed in the Low-EVE group compared with the High-EVE group.

Western blot analysis of the cell migration-associated proteins and adipogenesis-associated proteins.
Expression of adipogenesis-associated proteins
Protein expression of PPAR-γ, C/EBP, and AP2 was lower in the High-EVE group than in the Low-EVE group on day 9 (Fig. 6C). This indicates that differentiation of ASCs was lower in the High-EVE group than in the Low-EVE group.
FE simulation of tissue stiffness
An FE mesh of adipose tissue exposed to an EVE device is shown in Figure 7A. On the left is the original stiffness of the adipose tissue relative to its microscopic condition, while on the right is the stiffness change of the adipose tissue in EVE device. Adipose tissue stiffness increased in proportion to the amount of external force applied (Fig. 7B). The stiffness was increased in both groups when under external force. For the High-EVE group, the stiffness of adipose tissue increased more because of the higher external force. While for the Low-EVE group, the stiffness of adipose tissue increased less because of the lower external force. However, when the external force was removed, stiffness was restored to normal in both groups (Fig. 7C).

FE analysis.
Discussion
In our previous study, we found that daily application of the EVE device for 12 weeks significantly increased the adipose tissue volume. 10 It suggested that mechanical forces can increase cell migration in the early stages of adipose tissue regeneration and induce adipogenic differentiation in the late stages. However, the underlying mechanism is still unclear. In this study, we investigated how external force regulates cell migration and differentiation. Our results showed that application of external force using an EVE device upregulated expression of cell migration-related proteins and promoted migration of ASCs; however, excessive external force decreased adipogenic differentiation and increased fibrosis in adipose tissue. All these processes may be related to changes in adipose tissue stiffness (Fig. 8).

Potential mechanism by which changes in adipose tissue stiffness upon application of external force using an EVE device regulate the migration and differentiation of ASCs. Color images are available online.
Stiffness of adipose tissue plays a key role in cellular behavior such as proliferation, migration, and stem cell differentiation through mechanical transduction pathways.18,23,24 Cell migration, which is central to many biological processes including wound healing, tissue regeneration, and cancer progression, is sensitive to environmental stiffness.15,25,26 Durieux found that stiffness gradients control cell contraction by regulating myosin and that cells migrate at a constant rate toward areas of high stiffness. 27 Engler and coworkers reported that ASCs are more sensitive to stiffness than mesenchymal stem cells by culturing these cells on polyacrylamide gels with different stiffnesses.23,24 Moreover, ASCs differentiate into neurons, adipocytes, myoblasts, and osteoblasts upon culture on substrates with stiffnesses of 0–1, 2–4, 8–10, and >30 kPa, respectively.22,23 Therefore, ASCs differentiate into adipocytes in areas where stiffness is optimal.
FE analysis is a technique for reconstructing and evaluating stress, strain, and deformation in structures. 28 This methodology creates a representation of a structure that is readily resolved by mathematical analysis as a series of subdivisions, that is, it reduces a complex geometry into a finite number of elements with simple geometries that have the same properties as the original model.28,29 All FEs are described by differential equations and are solved by mathematical models. This approach has been widely used in medical research to study the human musculoskeletal system, to delineate skin and fat, and to mimic the breast shape in different positions.30–32 Therefore, we used FE analysis to simulate the force exerted by the EVE device and its effect on stiffness. Adipose tissue stiffness increased upon exposure to external force, decreased when this force was removed, and increased again when this force was reapplied.
This study demonstrates that when tissue stiffness increased upon application of external force, the number of ASCs increased, especially in the early stage. Expression of the cell migration-related proteins CDC42, RhoA, and dynamin also increased. CDC42 localizes to the leading edge of cells, where it triggers microtubule polarization and directs cell migration. 33 Moreover, CDC42 activates glycogen synthase kinase 3 α/β, which regulates CLASPS, APC, and stabilizing microtubule-associated proteins at plus ends, and thereby improves the capping, growth, and stability of microtubule.34,35 Microtubule depolymerization increases RhoA activity and cell contractility. 36 Microtubule dynamic changes of forming focal adhesions at the proximal end may increase cell contractility locally, thereby increasing focal adhesion assembly.34,37 RhoA-mediated contraction may also enhance retraction of the cell rear. 35 Moreover, dynamic microtubules repeatedly target mature focal adhesions, which dismantle at the cell rear, by interacting with plus-end tracking proteins. 38 Dynamin can localize focal contacts probably by promoting internalization of integrin complexes and play a role in focal adhesion disassembly in migrating cells. 39 In addition, dynamin interacts with microtubules, suggesting that microtubules can deliver dynamin to focal adhesions to trigger integrin-associated endosome internalization.34,40 Expression of CDC42, RhoA, and Dynamin, which are important for the early, middle, and late phases of cell migration, respectively, increased upon application of external force. Expression of cell migration-related proteins was delayed in the Low-EVE group compared with the High-EVE group. Some studies suggested that stiffness has a greater influence on cell migration than proteins. 19 External force increases tissue stiffness, which upregulates expression of CDC42, RhoA, and Dynamin, leading to migration of ASCs.
Although the number of ASCs increased in the High-EVE group, the long-term effect on adipose tissue was unsatisfactory. The lack of adipose tissue growth, hardness of adipose tissue, and high degree of fibrosis indicate that the increase in the number of ASCs was not conducive to adipogenesis. Sequential expression of PPAR-γ, C/EBP, and AP2 plays a significant role in terminal differentiation of preadipocytes into triglyceride-laden adipocytes.23,41 Expression of these three proteins was significantly lower in the High-EVE group than in the Low-EVE group on day 9. This suggests that excessive external force decreased adipogenesis and increased fibrosis in adipose tissue, which may be due to high tissue stiffness.20,21 The optimum stiffness for adipogenesis is ∼2–4 kPa. ASCs differentiate into muscle or bone when stiffness exceeds 4 kPa.22–24 High stiffness may explain why adipose tissue was harder and displayed a higher degree of fibrosis in the High-EVE group than in the Low-EVE group.
Therefore, tissue stiffness upon application of an EVE device is very important for long-term adipose tissue regeneration. When such a device is clinically used, external force should be increased to promote migration of ASCs and subsequently decreased to promote adipogenesis. The size of the external force can be changed according to the tissue stiffness required at various time points to promote long-term adipose tissue regeneration.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Nature Science Foundation of China (81772101, 81871573, 81801933, 81601702).