
Abstract
Adequate bone volume is required for osseointegrated implants to restore lost teeth and oral function. Several studies have demonstrated potential advantage of stem cells in regenerative medicine using osteoblasts. The periosteum is composed of osteoblasts, fibroblasts, and osteoprogenitor cells. It may be an alternative source for bone tissue engineering because of easy isolation and rapid proliferation in vivo and in vitro. Low-intensity pulsed ultrasound (LIPUS) has proved successful in recoveries from nonunions, delayed unions, and fracture of the bone in both animal experiments and clinical treatments. The study was to investigate the influence of LIPUS on the osteogenic differentiation in murine periosteum-derived cells (PDCs) and the underlying mechanism of LIPUS. PDCs were treated daily with LIPUS for 20 min up to 21 days with 3 MHz frequency, 30 mW/cm2 intensity, and pulse repetition frequency of 1 kHz. The effects of LIPUS on cell proliferation and viability were investigated. Osteogenic differentiation was analyzed by alkaline phosphatase (ALP)-positive cell staining, ALP activity assay, mineralized nodule formation, real-time reverse transcription-polymerase chain reaction, as well as western blotting. The results indicated that ultrasound stimulation did not significantly affect the proliferation of PDCs. But LIPUS significantly increased ALP activity on day 7 and markedly promoted formation of mineralized nodules on day 21. mRNA expression of ALP and osteocalcin was significantly upregulated by stimulation with LIPUS. LIPUS enhanced gene expression of both bone morphogenetic protein-2 (BMP-2) and osterix only in the presence of osteogenic medium. LIPUS stimulation did not affect Smad 1 and Smad 5 protein expression, but significantly upregulated protein levels of BMP-2 and phosphor-Smad 1/5/9 in PDCs. Thus, LIPUS stimulation increased early osteogenic differentiation in a normal medium and further enhanced expression of BMP-2 and subsequent osterix expression through the canonical Smad-signaling pathway in an osteogenic medium, leading to mineral apposition. Therefore, LIPUS might have potential to promote osteogenesis in PDCs.
Impact statement
There are few studies on periosteum-derived cells (PDCs) because conventional methods of their isolation are relatively difficult to procure abundant cells for cell culture and the total cell numbers are limited. In this study, a modified isolation technique of murine calvarial PDCs using gelatin is described. PDCs were initiated to emerge as early as day 3 and showed increased proliferation, which can be used for further studies.
Low-intensity pulsed ultrasound stimulation increased early osteogenic differentiation in a normal medium and further enhanced expression of bone morphogenic protein-2 and subsequent osterix expression through the canonical Smad-signaling pathway in an osteogenic medium, leading to mineral apposition.
Introduction
Mesenchymal stem cells possess the ability to differentiate into different lineages, including adipocytes, chondrocytes, osteoblasts, and nonmesoderm types.1–4 Traditionally, MSCs have been isolated from bone marrow; they can also be isolated from adipose tissue, fetal liver, umbilical cord blood, peripheral blood, and amniotic fluid.5,6 Recently periosteum-derived cells (PDCs) have become a highly attractive source in bone tissue engineering and regenerative medicine. They can be easily acquired by using a less-invasive method than bone marrow-derived mesenchymal stem or progenitor cells. 7
Ultrasound stimulation is a mechanical energy and can act as a biophysical signal that is conducted through the human body, resulting in complexed biochemical events inside the cells. Low-intensity pulsed ultrasound (LIPUS) has usually been considered as acoustic pressure waves that generated mechanical strains to elicit biochemical events. FDA approved LIPUS as a physiotherapeutic technique, to accelerate the fracture healing, delayed or nonunion of bone.8,9 The effectiveness of ultrasound stimulation has been shown through multiple in vivo studies.10,11
In addition, there are numerous reports on the stimulatory effects of LIPUS on different types of cells.12–14 These previous ultrasound and bone tissue studies have indicated that LIPUS stimulation increases the activity of osteoblasts and induces the osteogenic differentiation of stem and progenitor cells. Although experimental and clinical studies have demonstrated enhanced bone regeneration by LIPUS, according to our knowledge, there have been no reports on the influence of ultrasound treatment on the osteogenic differentiation of murine PDCs and its associated mechanisms. The study was to find out effects of LIPUS on the cell density as well as osteogenic differentiation of murine PDCs in vitro and the mechanisms of ultrasound on these cells.
Materials and Methods
Center for experimental animals of Tokyo Medical and Dental University approved the protocol design and research procedures of this study (A2017-343A).
Isolation and culture of periosteum-derived cells
Isolation of murine calvarium was performed as previously described with modification (Fig. 1A). 15 Using pentobarbitone sodium euthanasia, skulls of ten 4-week-old SLC imprinting control region female mice (Sankyo Lab, Japan) were collected. The skulls were trimmed to 10 × 10 mm2 in size, and washing thoroughly with phosphate-buffered saline (PBS), the periosteum was peeled off the bone (Fig. 1B).

Isolation of mouse calvaria.
Isolation of PDCs was carried out as previously described with modification. 16 In brief, 600 mg of gelatin was immersed in 6 mL of high-glucose Dulbecco's modified Eagle's medium (Sigma-Aldrich, Japan) supplemented with 10% (v/v) fetal bovine serum (Sigma-Aldrich) and 1% (v/v) penicillin-streptomycin-amphotericin B suspension (Fujifilm Wako Pure Chemical Corporation, Japan) in 60 mm culture dishes (Fig. 2A).
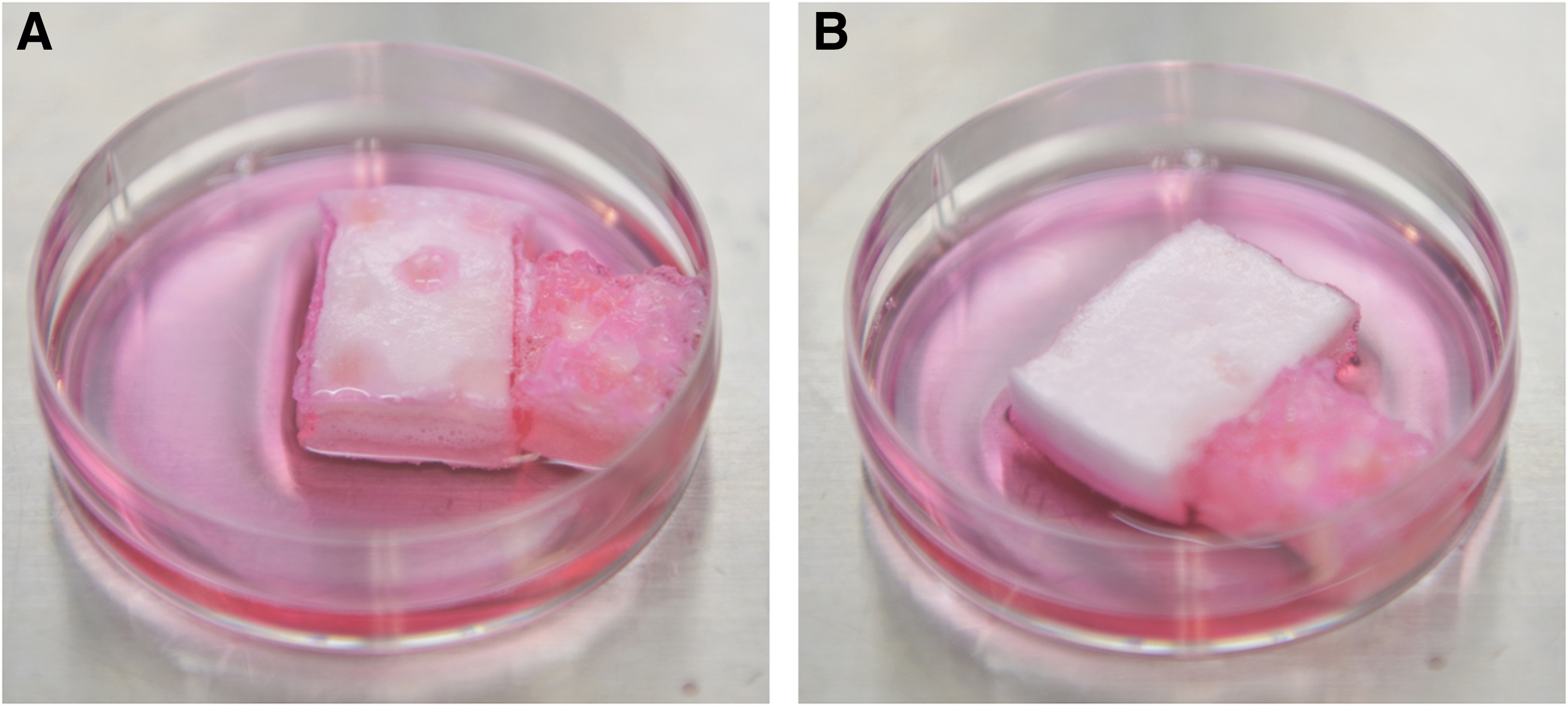
Periosteum-derived cell culture.
Periosteum were placed on the gelatin and flipped to ensure that the periosteum was sandwiched in between the culture dish and gelatin (Fig. 2B). The cells were incubated in a 37°C incubator with CO2 5% and air 95%. After 2–3 days, cells started to come out from the calvarial periosteum (Fig. 3A). The medium was changed fresh every 3 days. When PDCs showed 70% confluence, they are passaged using 0.5 w/v% trypsin EDTA solution without including phenol red (10 × ) (Fujifilm Wako Pure Chemical Corporation).
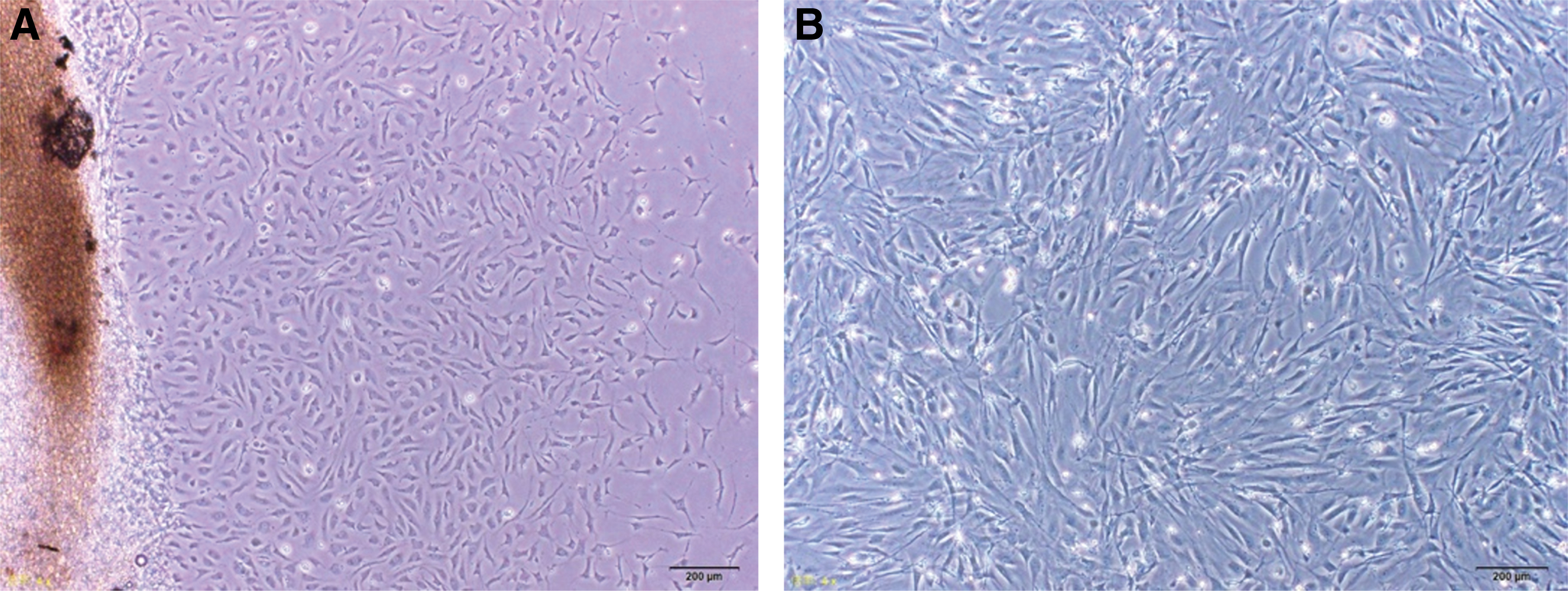
PDCs.
PDCs at passage 3 were used in this study (Fig. 3B). PDCs were seeded with cell density of 3 × 105/well in 6-well culture plates or 5 × 103/well in 96-well culture plates. The cells were cultured in DMEM or osteogenic medium. Fifty microgram per milliliter of
Ultrasound stimulation
After 72 h of incubation, the cells were exposed to LIPUS. In brief, OSTEOTRON D2 (Ito Co., Ltd, Japan) was used (Fig. 4). A 35 mm diameter transducer surface was circular, and the ultrasound intensity was nearly constant throughout its cross-sectional area. The beam nonuniformity ratio of 3 MHz ultrasound probe is 2.7% ± 30% (IEC standards—International Electrotechnical Commission). On the transducers, the culture plates were placed with coupling gel in between them. The cells were exposed to ultrasound irradiation as per the following: 3 MHz frequency, 30 mW/cm2 intensity, pulse repetition frequency of 1.0 kHz, 1:4 pulse duty cycle (2 ms “on” and 8 ms “off”), and spatial average-temporal average. Cells are treated with LIPUS for 20 min daily. Controls were prepared in the same way without switching on the machine.
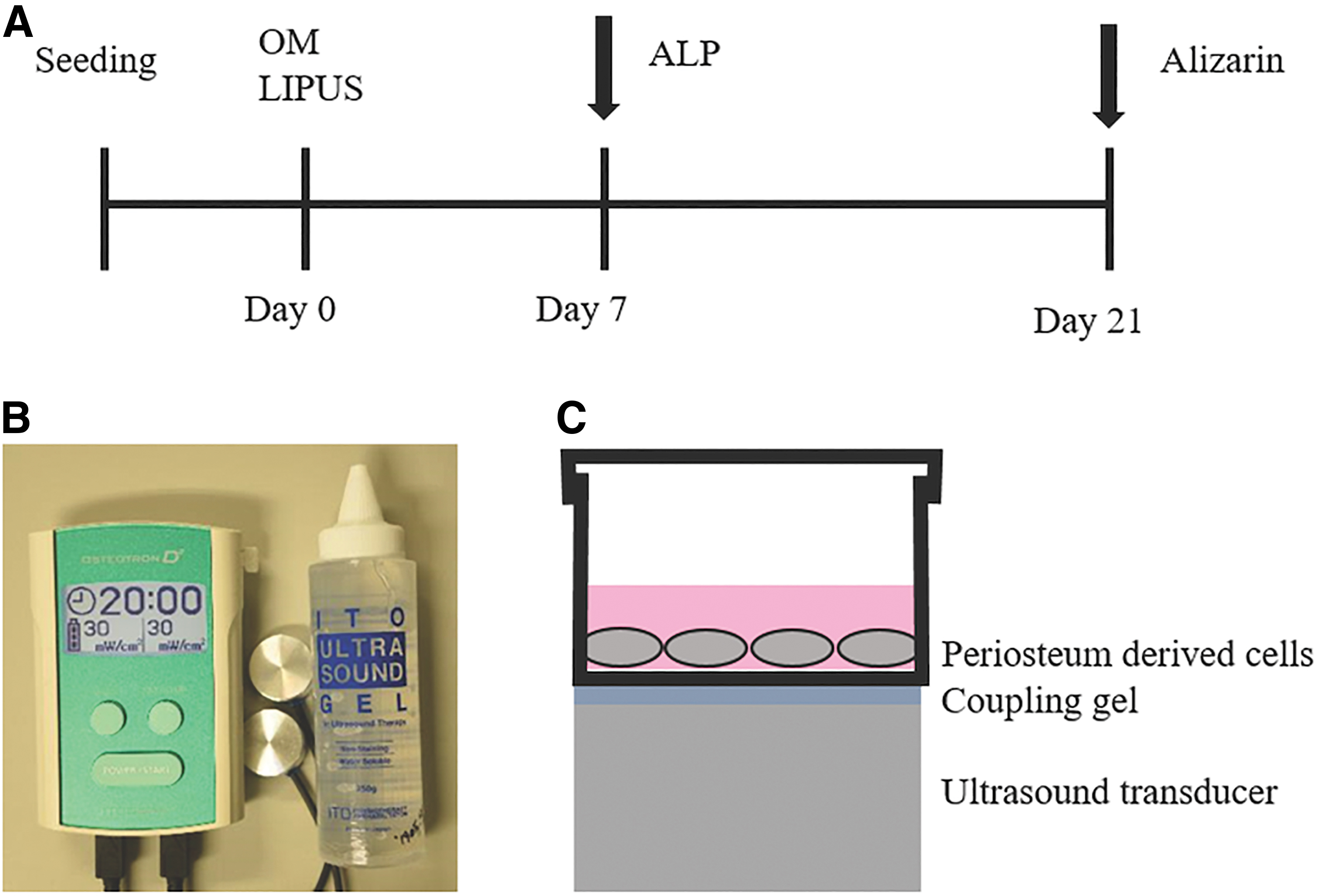
LIPUS application.
Cell proliferation/cytotoxicity assay
Cell counting kit-8 assay (CCK-8; Dojindo, Tokyo, Japan) was used to analyze cell proliferation or cytotoxicity as mentioned in the manufacturers' protocol. In brief, PDCs were seeded as above with LIPUS or without LIPUS treatment for up to 14 days (n = 6). The culture medium was replaced every 2 days. On days 0, 1, 3, 5, 7, 10, and 14, 10 μL CCK-8 solution was added and incubated for 1 h at 37°C and then measured the absorbance at 450 nm with a microplate reader (ARVO MX; Perkin Elmer, Japan). The cell density was calculated from the absorbance values using a standard curve.
Alkaline phosphatase activity assay
On day 7, the cells were lysed with 500 μL solution of 0.1% Triton solution (Triton™ X-100; Sigma-Aldrich) following the instructions of manufacturer. The cells were freezed for 15 min at −70°C and melted at 4°C for another 15 min. The procedure was repeated two times. The solution was transferred into 1.5 mL microtube. The microtubes were put onto the ice and sonicated for 5 min to disrupt the cell membranes by using ultrasonic machine (Branson, Japan). The microtubes were then centrifuged at 15,000 rpm for 15 min at 4°C. Then sample solution was used to analyze the alkaline phosphatase (ALP) activity and measure protein content.
Quantitative ALP activities were analyzed by assaying 20 μL of sample using LabAssay™ ALP (Fujifilm Wako Pure Chemical Corporation). In brief, using 96-well plate, samples were mixed with 100 μL of p-nitrophenylphosphate solution. Then, the sample was incubated for 15 min at 37°C. The ALP reaction was stopped by adding of 80 μL of NaOH stop solution, and ALP activity was analyzed using a microplate reader (ARVO MX; Perkin Elmer) measuring the absorbance at a wavelength of 405 nm. Average measurement of blank standard replicates was substrate from all other standard and sample replicates. Standard curve was prepared by plotting optical density of standard versus its concentration in nmol/μL. ALP activity (units/μL) was calculated using the manufacturer's formula.
To determine protein concentrations, in each well of 96-well plate, 20 μL of sample was added. Then, 200 μL of bicinchoninic acid (BCA) working reagent was added (Pierce™ Rapid Gold BCA Protein Assay Kit; ThermoFisher SCIENTIFIC, Japan). The samples were incubated for 5 min at room temperature, quantified using a microplate reader (ARVO MX; Perkin Elmer) measuring the absorbance at a wavelength of 480 nm.
Average measurement of blank standard replicates was substrate from all other standard and sample replicates. Standard curve was prepared by plotting optical density of each bovine serum albumin standard versus its concentration in μg/μL. Protein concentration was determined by using standard curve. ALP activity was normalized with total protein concentration (units/μg protein/min). Data are expressed as means ± standard deviations.
Alkaline phosphatase-positive cell staining
ALP-positive cells were stained using the ALP Stain Kit (Fujifilm Wako Pure Chemical Corporation). After washing with PBS for two times and fixing with 3.7% formalin for 10 min, cells were then washed again with PBS and incubated in 1 mL ALP staining solution for 45 min at 37°C, and then, washed with PBS. Images of ALP-positive (blue) cells were captured using a BZ-X700 microscope (Keyence, Japan).
Alizarin red S staining
At day 21, after fixation, the culture plates were placed for 1 h in 70% ethanol, and subsequently stained for 10 min using 2% Alizarin Red S solution. Then, the cells were washed with PBS, and calcium nodule formation was captured under phase-contrast microscopy using a BZ-X700 microscope (Keyence). Quantification of staining was analyzed as previously. 18 In each well, 10% (v/v) acetic acid (800 μL) was added and then shaken at room temperature for 30 min. The staining was then collected and transferred to 1.5 mL microtest tubes. First vortexed for 30 s, then heated for 10 min at 85°C, and cooled by placing on the ice for 5 min. Then, centrifuged for 15 min using 20,000 g and then 200 μL of 10% (v/v) ammonium hydroxide was added. Of the sample, 100 μL was added into each well of 96-well plate. Alizarin Red S quantification was analyzed using a microplate reader (ARVO MX; Perkin Elmer) measuring the absorbance at a wavelength of 405 nm.
Real-time reverse transcription-polymerase chain reaction
PDCs at passage 3 were seeded in six-well plates and cultured in DMEM or the osteogenic medium for up to 14 days with or without daily LIPUS stimulation. Total RNA was extracted from each sample on days 7 and 14 using the TRIzol reagent (Invitrogen, Japan), and cDNA synthesis was carried out using the SuperScript™ IV VILO™ Master Mix with ezDNase™ Enzyme (Invitrogen). mRNA expression was analyzed using RT-PCR in ABI Prism 7300 Sequence Detection System (Applied Biosystems, Japan) with PowerUp™ SYBR Green Master Mix in the following program: 95°C for 5 min; cycles of 35 for 15 s at 95°C, and for 1 min at 60°C. The specific primers sequences are described in Table 1. The gene of interest expression was normalized to that of the GADPH and then relative expression was analyzed to the control group using 2−ΔΔCT method.
Real-Time Polymerase Primers
ALP, alkaline phosphatase; BMP-2, bone morphogenetic protein-2; GADPH, glyceraldehyde 3-phosphate dehydrogenase; OCN, osteocalcin; Osx, Osteoblast-specific Transcription Factor Osterix; PCR, polymerase chain reaction; Runx2, Runt-related transcription factor 2; Smad, Caenorhabditis elegans SMA (“small” worm phenotype) and Drosophila MAD (“Mothers Against Decapentaplegic”).
Western blot analysis
At day 7, after washing with PBS, the cells were then exposed to a cell lysis buffer using NUPAGE LDS sample buffer (4 × ) (Invitrogen). After determination of protein concentrations using Nanodrop, equal amounts of total protein (40 μg each sample) were dissolved in 20 μL running buffer, were heated for 10 min at 70°C, and electrophoresis was performed using NUPAGE Bis-Tris gels 4–12% (Invitrogen) for 30 min. Then, transferred to polyvinylidene difluoride membranes using iBlot system, followed by blocking for 30 min before incubation with primary antibodies against Smad1(D59D7) XP® Rabbit mAb (No. 6499), Smad5(D4G2) Rabbit mAb (No. 12534), Phospho-Smad1(Ser463/465)/Smad5 (Ser463/465)/Smad9 (Ser465/467) (D5B10) Rabbit mAb (No. 13820; Cell Signaling Technology, Japan), GADPH Rabbit mAb (EPR16891), and bone morphogenetic protein-2 (BMP-2) Rabbit pAb (ab14933) (Abcam, Japan) for overnight at 4°C, incubated for 1 h with anti-rabbit IgG secondary antibodies. The proteins were then detected using the Chromogenic Western Blot Immunodetection Kit (WesternBreeze; Invitrogen). The relative intensities of the developed bands were photographed, calibrated, and analyzed using image analyzing software ImageJ 1.8.0_112(NIH) and expressed as ratios.
Statistical analysis
Statistical analysis was performed using statistics software SPSS version 22.0 for Windows (Chicago, IL). All data are presented as means ± standard error of the mean. Data were analyzed using one-way analysis of variance followed by the Tukey's multiple comparison test. Data were considered statistically significant if p value was <0.05.
Results
Cell proliferation
The CCK-8 assay was performed up to day 14 on PDCs cultured with or without ultrasound stimulation or osteogenic medium (OM). Exposure to ultrasound stimulation with or without OM at 30 mW/cm2 did not produce significant impact on cell proliferation compared with the control cells (Fig. 5).

LIPUS stimulation on cell proliferation. Murine PDCs were cultured with or without transient LIPUS stimulation and the cell numbers were determined on days 0, 1, 3, 5, 7, 10, and 14 of culture (n = 6). The data are shown as the mean ± SD (p > 0.05). SD, standard deviation.
ALP activity
At day 7, ALP activity of L+O+ and L–O+ was significantly higher than that of L–O– (p < 0.001) (Fig. 6). Furthermore, the L+O+ showed significantly higher ALP activity than the L–O+ (p < 0.01) and the L+O– showed significantly higher ALP activity than the L–O– (p < 0.01). Thus, LIPUS significantly increased the ALP activity and promoted osteogenic differentiation, and LIPUS stimulation alone in absence of OM could enhance the osteogenic activity.

LIPUS stimulation on ALP activity. Murine PDCs were exposed daily to LIPUS stimulation for 20 min, and the activity of ALP was determined on day 7 of culture. The data are represented as mean ± SD (n = 6). (*p < 0.05).
ALP-positive cells and mineralized nodule formation
The results of ALP staining were in accordance with the results of the ALP activity assay (Fig. 7A). In alizarin red staining, L+O+ group showed the highest intensity than that of all other groups. In addition, the combination of LIPUS stimulation and OM significantly enhanced calcium nodule formation (Fig. 7B). In quantitative analysis of alizarin red S staining, the L+O+ group showed significant increase in intensity compared with that in other groups (Fig. 8).


Quantification of Alizarin red staining (n = 3; *p < 0.05).
mRNA expression
The L–O+ showed an increase in ALP gene expression significantly higher than that in the L+O– (p < 0.05), but no significant difference was observed between the L+O+ and L–O+. Both L+O+ and L+O– showed significantly higher ALP levels than that in the L–O– (p < 0.01) (Fig. 9A). For Osx gene expression, the L+O+ and L–O+ showed significant differences compared with the L–O– (control). However, no significant difference was observed between the L+O+ and L–O+ (Fig. 9B). BMP-2 gene expression was significantly higher in the L+O+ than in the L+O– and L–O– (p < 0.05) (Fig. 9C). OCN expression levels were the highest in the L+O+, which was significantly different with that in the L–O+ and L–O– (p < 0.001). OCN expression was also significantly increased in the L+O+ when compared to that in the L+O– (p < 0.01) (Fig. 9D). These results are consistent with the alizarin red staining result. LIPUS alone did not increase Osx and BMP-2 gene expression significantly. Smad 1 and Smad 5 expression did not have significant differences between all groups (Fig. 9E, F).

The osteogenic mRNA expression by RT-PCR (*p < 0.05; **p < 0.01; ***p < 0.001, ****p < 0.0001). BMP-2, bone morphogenetic protein-2; OCN, osteocalcin; Osx, Osterix; RT-PCR, reverse transcription-polymerase chain reaction; Smad, Caenorhabditis elegans SMA (“small” worm phenotype) and Drosophila MAD (“Mothers Against Decapentaplegic”).
Change in bone morphogenetic protein-2 and phosphor-Smad 1/5/9
The results of western blot analysis showed that LIPUS stimulation induced a detectable increase in BMP-2 and phosphor-Smad 1/5/9 relative to the control group when the cells were treated with 30 mW/cm2 20 min daily. Total Smad 1 and Smad 5 protein level was not found to exhibit any considerable change (Figs. 10 and 11). The protein level of BMP-2, Smad 1, and Smad 5 showed a similar trend with its mRNA level.

Representative images of western blotting. GADPH, glyceraldehyde 3-phosphate dehydrogenase. Color images are available online.

The protein levels of Smad 1, Smad 5, BMP-2 and Phospho-Smad 1/5/9 were quantified by normalization to GADPH. Data are presented as mean ± SD (n = 3); (*p < 0.05).
Discussion
PDCs are potential autologous cell source for regenerative medicine and bone tissue engineering, mainly due to its high regenerative potential and easy access. 19 LIPUS has been used as an effective and safe method in bone regeneration, as it can transmit mechanical energy into the body in the form of acoustic wave, producing complexed intracellular events.20,21
LIPUS applications for fracture healing and the treatment of nonunion were already approved by the U.S. FDA. Previously, 1 kHz pulse repetition frequency was found to significantly increase calcium accretion levels, 22 and 30 mW/cm2 for 20 min daily resulted in increased osteogenic differentiation and mineralization in rat bone marrow stromal cells. 23
A previous study demonstrated that the osteoinductive response in vitro and the acceleration of bone regeneration in vivo induced by LIPUS were through its nonthermal effects.24,25 Integrins promoted formation of transmission channels due to focal adhesions in the process of signal conversion. 26 LIPUS can also contribute to the healing of bone fractures, chondrogenesis, angiogenesis, endochondral ossification, intramembranous ossification, and bone remodeling. 27 However, the use of ultrasound stimulators in these treatments is still controversial because of adverse effects, and the mechanisms involved in the fracture healing process need to be further studied.
In the present study, LIPUS did not significantly increase cell density with or without OM for up to 14 days, indicating that use of LIPUS caused no harm to cells and serve as being safe for clinical use. A significant increase in ALP activity in PDCs was observed after 7 days of LIPUS stimulation using 30 mW/cm2 for 20 min daily. These findings were similar to results of previous report in that LIPUS treatment on osteoblasts increased ALP activity after day 5. 28 L+O+ group showed more significant ALP activity than L–O+ group (p < 0.05) and L+O– showed more significant ALP activity than L–O– (p < 0.05). These results are in accordance with ALP staining and LIPUS treatment stimulated the PDCs differentiation into osteogenic lineage by increasing ALP activity. Mineralized nodule formation in L+O+ was significantly increased than in the other groups. This result indicated that LIPUS might have synergistic action with OM, enhance mineralization, and increase calcium nodule formation in the presence of OM. It was demonstrated that bone healing was a result of an increase in intracellular Ca2+ concentration that occurred within seconds after LIPUS stimulation 29 ; 3 MHz daily LIPUS up to 14 days could enhance bone formation. 30
To confirm LIPUS-induced osteogenic differentiation of PDCs, expression of osteogenic marker genes such as ALP, Osx, and OCN was evaluated by real-time reverse transcription-polymerase chain reaction (RT-PCR). L+O+ and L+O– groups showed elevated ALP gene expression significantly more than L–O– (p < 0.01). Osx was less expressed in L+O+ than that in L–O+, although there was no significant difference. L+O+ and L–O+ elevated Osx gene expression significantly higher than L–O– (p < 0.01 and p < 0.0001 respectively). For OCN gene expression, L+O+ showed the highest value and was significantly different than that in the L–O+ (p < 0.05). Osteocalcin plays a role in bone mineralization, and is considered a late-stage marker of osteogenic differentiation and bone maturation process, which is in accordance with our findings that mineralized nodule formation upon LIPUS stimulation increased in PDCs. This indicates that LIPUS and OM synergistically increase osteogenic differentiation.
To further confirm LIPUS stimulation on osteogenic differentiation of PDCs, expression of BMP-2, Smad-1, and Smad-5 was evaluated by real-time PCR and western blot analysis. LIPUS treatment upregulated significantly in BMP-2 gene and protein expression in PDCs. L+O+ showed an increase in BMP-2 expression significantly than L+O– and L–O– (p < 0.05, respectively), but no significant difference was found between groups in gene expression of Smad-1 and Smad-5. Phosphorylated-Smad 1/5/9 protein expression was significantly increased in L+O+ group than in L+O– and L–O– groups (p < 0.05, respectively). These findings indicated that LIPUS stimulation enhanced BMP-2 expression.
The previous study suggested the mechanism of LIPUS in enhancing osteogenic differentiation and bone fracture repair. 31 However, until now, the complete mechanism by which LIPUS promotes BMP-2 expression has been unclear. Therefore, here, we suggest a possible mechanism of BMP-2 enhancement by LIPUS stimulation based on previous studies.
The mechanism by which LIPUS enhanced osteogenic differentiation and increased intracellular calcium and mineralized nodule formation was suggested as follows: 200 ms pulses produce nanovibrations 32 as well as acoustic pressure 33 to cells. Vinculin acts as a mechanosensor of the mechanical stimuli generated by LIPUS at cell-extracellular matrix (ECM) adhesion. Integrin-mediated focal adhesion kinase activation leads to actin rearrangements and cell migration. 34 Integrin-dependent cellular attachment and detachment with ECM results in cell migration that later induces intracellular reorganization with the production of reactive oxygen species (ROS).35–37 The accumulation of ROS causes oxidative damage and cell apoptosis. To control ROS, prevent the cell damage, and regain redox homeostasis, a series of mechanisms lead to the production of heat shock protein (HSP). HSP70 protein promoted cell viability and decreased cell apoptosis and can thereby protect against damage of the cell by ROS.38–40 LIPUS stimulation upregulates HSP70, 41 which in turn upregulates the cyclooxygenase-2 (COX-2) expression, leading to the production of the prostaglandin E2 (PGE2). 42 COX-2 played a major role in PGE2 production in osteoblasts. 43 PGE2 produced by COX-2 results in increased BMP-2 expression through the EP4 receptor.44,45 LIPUS leads to the expression of BMP-2.46,47 BMP-2 vastly increased osteocalcin release from MSCs. 48 BMP-2 significantly increased the calcium concentration by the influx from extracellular calcium and the release of calcium from endoplasmic reticulum.49,50 BMPs have been implicated in differentiation of osteoblasts and formation of the bone. 51 There has been reported that BMP-2 upregulated Osx expression.52,53 Notably, BMP-2 treatment increased Osx expression in Runx2 knockout mice mesenchymal cells.54,55
In conclusion, our findings demonstrated that LIPUS stimulation increased ALP activity in PDCs and promoted mineralized nodule formation and upregulated Osx, OCN, and BMP-2 expression together with OM. These findings suggest that LIPUS treatment enhances the osteogenic differentiation and mineralized nodules formation in PDCs. Therefore, we concluded that LIPUS-mediated mechanism of osteogenic differentiation may be achieved via upregulation of BMP-2 expression and through activation of the BMP-Smad canonical pathway in PDCs (Fig. 12). Our results might provide a significant evidence for clinical usage of LIPUS in bone regeneration. Since these in vitro findings cannot be associated to the in vivo scenario directly, further studies will be necessary for better understanding of the mechanism of LIPUS for its safe and efficacious use.

The BMP-Smad canonical pathway is shown. LIPUS increase BMP-2 expression. BMP-2 ligand binds and activates BMP receptor complex leads to R-Smads 1, 5, and 9 phosphorylation. R-Smads are released from the cell membrane and binds to Co-Smad proteins. The complex was imported into the nucleus and activated osteogenic gene Osx transcription. BMPRI, bone morphogenetic protein receptor type-1, BMPRII, bone morphogenetic protein receptor type-2; Co-Smad, common-mediator Smad; P, phosphorylated; R-Smad, receptor-regulated SMAD. Color images are available online.
Footnotes
Acknowledgments
Moreover, the authors also greatly appreciate the kind help of Professor Dr. Mari Yuasa (Tokyo Medical and Dental University, Chemical Biology Screening Center, Tokyo, Japan) for providing the necessary materials. The authors also express their gratitude to Dr. Michiko Suzuki, Dr. Quan Sun, and Dr. Kaori Yokota (Oral Implantology and Regenerative Dental Medicine Department, Tokyo Medical and Dental University, Tokyo, Japan) for technical advice.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was kindly supported by a research grant provided by the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.