
Abstract
The placenta acts as an interface between the fetus and the expecting mother. Various drugs and environmental pollutants can pass through the human placental barrier and may harm the developing fetus. Currently available in vitro placental barrier models are often inadequate, because they are lacking the functional trophoblast cells. Therefore, we developed and characterized a new human placental model using trophoblast stem cells (TSCs) derived from human induced pluripotent stem cells. Umbilical vein endothelial cells, fibroblast, and TSCs were cocultured using micromesh cell culture technique. These cells formed a tight three-layered structure. This coculture model induced progressive fusion of TSCs and formed a syncytialized epithelium that resembles the in vivo syncytiotrophoblast. Our model allowed the cultured trophoblasts to form microvilli and to reconstitute expression and physiological localization of membrane transport proteins, such as transporter for ATP-binding cassette subfamily B member 1, ATP-binding cassette subfamily C member 3, and glucose transporter-1. Drug permeability assays were performed using five compounds. The results from the permeability assays were comparable to the ones obtained with ex vivo placental models. In conclusion, we developed a novel coculture model mimicking human placenta that provides a useful tool for the studies on transfer of substances between the mother and fetus.
Impact statement
Compared with the currently available in vitro placental barrier models, a novel three-dimensional coculture placental barrier model presented in this study morphologically and functionally modeled the true placental barrier. The use of human trophoblast stem cells from human induced pluripotent stem cells substantially improved the current model. The use of micromesh sheet as a bioscaffold facilitated the formation of a good multilayer structure, which is closer to the physical appearance of the placenta observed in human.
Introduction
Placenta plays a key role in fetal growth and development1–3 (Fig. 1A). Placenta forms a barrier that regulates the exchange and accumulation of various endogenous and exogenous substances, such as gases, nutrients, and waste products, between the mother and fetus (Fig. 1B). Placenta is an essential barrier to protect the growing fetus from exposure to xenobiotics, however, it has been recognized that various drugs, environmental pollutants, and even certain nanoparticles could go through the placental barrier.4–6 Studies on drug transfer from the maternal to the fetal side and their potential teratogenic and toxic effects have increased in recent times. Nevertheless, appropriate development of drugs and their safety evaluation for pregnant woman and the fetus are still markedly underdeveloped. 4
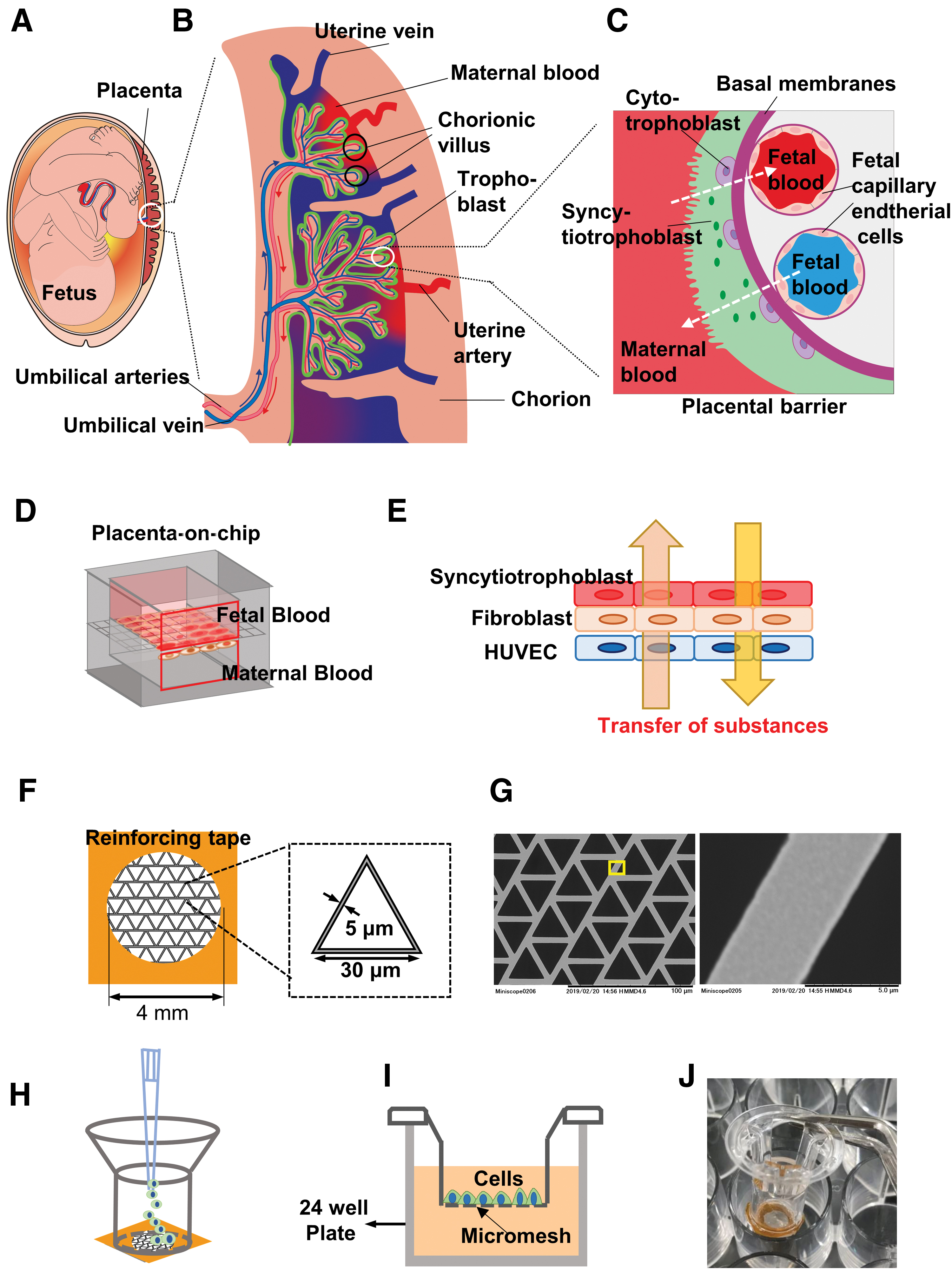
Design of 3D placental barrier model.
Several in vivo, ex vivo, and in vitro placental barrier models have been developed to accurately measure the transfer of substances between the mother and fetus. Important insights about the placental barrier have been gained using in vitro cell culture models.7–9 Several cell types have been used for the research on the placental barrier in vitro, such as BeWo and Jeg-3 choriocarcinoma cells, isolated primary placenta cells, and the first trimester human trophoblast cell line ACH-3P. 10 Primary trophoblast cells are difficult to proliferate in vitro, and there are ethical issues about acquiring cells from human placental tissue and blastocysts. Choriocarcinoma-derived cell lines are easy to proliferate and share numerous characteristics of primary trophoblast cells, however, they differ in several aspects due to their malignant transformation. 11
In the first trimester, the placental barrier is composed of four different layers, namely the syncytiotrophoblast (STB), the mononuclear cytotrophoblast (CTB), the merged stroma of fetal capillary and the STB, and the fetal capillary endothelial cells (Fig. 1C). CTBs derived from the trophectodermal layer are trophoblast stem cells (TSCs), and they differentiate into STB. 12 Recently, human embryonic stem cells and human induced pluripotent stem cell (hiPSC)-derived trophoblast cell lines, have been immensely informative for placenta research.13,14 In our previous study, we established the TSCs derived from the hiPSCs. 15 These TSCs derived from hiPSCs were highly proliferative and were able to differentiate into STB-like cells.
Recently, Transwell technology has developed to become a popular alternative for drug testing and toxicology in vitro. The membrane used in the Transwell is polycarbonate or polyester membrane with 0.4–3.0 μm of pore size. The rate of pores is about 10%. Therefore, the membrane is almost like the usual two-dimensional (2D) tissue culture plate. The 2D cultures using tissue culture plate induce mechanical stimuli in the cells, which affects tissue-specific phenotypes.16,17 When evaluating the cell-specific functions, the cells cultured by cell–cell adhesion rather than cells cultured by cell–substrate adhesion are more desirable.
We originally developed a cell culture technique using a microstructured parylene-coated nickle mesh sheet as a bioscaffold. The apertures of these micromeshes are much larger than the size of single cells, hence the cells in these openings bind each other only by cell–cell adhesion.18–20 Additionally, micromesh cell culture has several features not found in other 2D culture methods. Hori and Kurosawa reported that micromesh-cultured hepatic cells have been shown to exhibit increased expression of hepatic marker genes and enhanced cell maturation. 18
Previous reports by Okeyo et al.19,20 have shown that limited culture on micromesh can induce hiPSCs to differentiate into trophoblast cysts. Since the cells are suspended in the culture medium in a cell sheet state maintained by cell–cell adhesion, nutrition and oxygen can be supplied directly from both sides of the cell sheet. 18 These features of micromesh might help to form a functional placental barrier model. Additionally, we recently found that upon forming the first cell sheet and by subsequently seeding different cells, a multilayer cell sheet using arbitrary cells can be formed. Since the placental barrier is composed of three different cell layers, micromesh as a scaffold might be appropriate for in vitro three-dimensional (3D) placental model.
In this study, we aimed to establish a multilayer 3D placental barrier model using micromesh culture techniques (Fig. 1D). Considering that not only the STB but also the microvascular endothelial cell layer strongly contributed to the permeability of the substance through the placenta; in our model, hiPSC-derived TSC and vascular endothelial cells were placed on both sides of the fibroblast layer (Fig. 1E). After a comprehensive characterization of the coculture model, transport studies were performed with FITC–dextran, and lipid- and water-soluble drugs. To our knowledge, this is the first placental coculture model using hiPSC-derived TSC to assess transport of drugs.
Methods
Cell culture
Human umbilical vein endothelial cells (HUVECs, C2519A; Lonza) were cultured in the EGM™-2 Endothelial Cell Growth Medium-2 BulletKit™ (Lonza). The maximum population doublings of C2519 HUVECs are guaranteed to further expand for 15 population doublings according to the instructions. In our culture condition, the number of population doublings every day of the C2519A HUVECs was 0.57. In this study, we performed the functional experiments on drug transport after HUVEC seeding for 10 days. HUVECs (passage 3–4) were used for seeding in all experiments. Furthermore, we tested the live cell level using calcein staining (Supplementary Fig. S1A) and cellular senescence levels using the Senescence-Associated Beta-Galactosidase (SA-β-gal) Detection Kit (Dojindo, old cells showed obvious SA-β-gal response) (Supplementary Fig. S1B) on 14-day single HUVEC sheet. HUVECs were healthy and functional in our experiment time point (Supplementary Fig. S1A, B).
TSCs induced from hiPSC line (ChiPSC22; TaKaRa Bio),
15
were cultured in TSC basic medium.
21
Briefly, the cells were cultured in TSC basic medium (Dulbecco's modified Eagle's medium [DMEM]/F12 medium supplemented with 0.1 mM 2-mercaptoethanol, 0.2% fetal bovine serum [FBS], 1% penicillin/streptomycin, 0.3% bovine serum albumin [BSA], 1% ITS-X supplement, 1.5 μg/mL
Design and fabrication of the placental barrier
Fabrication of the placental barrier was carried out using the micromesh culture method. A commercially available nickel micromesh was used as bioscaffold instead of traditional porous membrane. Briefly, a nickel micromesh with triangular shape (Optnics Precision Co., Ltd., Ashikaga, Japan), having side length of 30 μm, thickness of the sheet at 5 μm, and the width of the mesh strand at 5 μm was obtained (Fig. 1F, G). The micromesh was cut into 0.6 × 0.6-cm sections and fixed on a silicon sheet (1 × 0.5 × 1 cm, length [l] × height [h] × width [w]) using a Kapton tape (Fig. 1F). The silicon sheet and Kapton tape had a 4-mm hole in the center to act as a frame reinforcing the ultrathin mesh sheets (Fig. 1F).
Our previous studies showed that hiPSCs proliferation was not ideal for direct contact with the untreated nickel mesh, and the old waste products produced by cells, including lactic acid dissolved the nickel metal (data not shown). To solve this, the mesh was coated with a 0.2 μm thickness of thin layer of parylene, which is widely used as a harmless biomaterial., 22 in a PDS 2010 system (Specialty Coating Systems). Parylene coating effectively prevents direct contact between cell and nickel mesh and do not influence the proliferation of human cells. Due to the parylene film's rough hydrophobic surface, the micromesh was then air plasma treated using a plasma cleaner (PDC-001-HP; Harrick Plasma) at a high radio frequency for 3 min. The reinforced mesh sheets were bonded to the transwell without the membrane at 55°C for 4 h (Fig. 1H). The well with mesh sheet (mesh-well from here on) (Fig. 1H) was sterilized using ultraviolet irradiation for at least 90 min.
Following sterilization, the mesh well was placed into the 24-well cell culture plate. For coculture of three cell lines, the protocol is shown in Figure 2A. One hundred microliters of HUVEC suspension (1 × 105 cells) was gently introduced in the mesh well (Fig. 1H). After incubation at 37°C for 4 h, 2.5 mL of HUVEC medium was added to the well (Fig. 1I, J). Culturing the cells for 3–4 days resulted in a monolayer sheet of HUVECs. Subsequently, for culture of fibroblast cells, the HUVEC medium was discarded and 100 μL of Tigs suspension (1 × 105 cells) was gently introduced on the HUVEC sheet. After incubation for 4 h, 2.5 mL of HUVEC medium was added to the well. A HUVEC-Tig sheet was constructed by incubating for 36 h. For TSC culture, 10 μL of 0.5 mg/mL iMatrix-511 silk (Nippi) was added to the TSC suspension. One hundred microliters of TSC suspension (1 × 105 cells) was gently introduced on the top of HUVEC-Tig sheet. After incubation for 4 h, 2.5 mL of HUVECs:TSCs (1:1) medium was added to the well. After 4 days of incubation, the three layers of cells characterized as HUVEC-Tig-TSC was constructed. For differentiation of TSCs into STBs, the medium was changed into HUVECs:STBs (1:1) medium and cultured for 2 days. We characterized the three layers of cells as HUVEC-Tig-STB sheet.

Establishment of multilayer cell culture placental barrier model.
Real-time quantitative polymerase chain reaction analysis
The experiments were performed using published methods. 15 The primers used for real-time quantitative polymerase chain reaction (qRT-PCR) are shown in Supplementary Table S1.
Immunostaining
The experiments were performed using published methods. 15 The antibodies used are shown in Supplementary Table S2. Nuclei were stained with propidium iodide (PI, 1:1000) or Hoechst 33342 (1:1000). At least three independent trials were performed for each experiment.
3D optical coherence tomography and visualization
Optical coherence tomography (OCT) and subsequent visualization was performed as previously described. 23 Minor changes were made to increase the optical resolution to 3 μm: the voxel size was 2 × 2 × 2 μm and the beam diameter was 10 μm. The average data acquisition window was 200 × 200 × 200 μm (h × w × l) correlating with 200 scans per axis in 5 min. For 3D images, original cross-sectional images captured using the OCT system were processed using the ImageJ plugin (National Institute of Health). Two independent trials were performed for the experiment.
Evaluation of barrier integrity
Placental barrier model was prepared as described above. Placental barrier integrity was measured with an EVOM2 epithelial voltohmmeter (World Precision Instruments Ltd.) containing 600 μL of medium on the basolateral side and 100 μL of medium on the apical side of transwell. Transendothelial electrical resistance (TEER) values were corrected for the surface area (Ω·cm2).
In vitro drug transport experiments
To measure the flux of drugs (Table 1) across the placental layers, following measurement of TEER, cell culture inserts were transferred to 24-well plates containing 2.5 mL HBSS solution (Thermo Fisher) in the basal compartment. The inserts were then transferred to a new basal compartment containing 600 μL HBSS with 4.5% BSA solution. In the apical chamber, culture medium was replaced by 100 μL HBSS with 4.5% BSA solution containing 1mM atenolol and acyclovir, and 0.1 mM caffeine, metoprolol, and propranolol and incubated for 30 min in 5% CO2 at 37°C. The concentrations of the test molecules in samples from the apical and basal compartments were determined by liquid chromatography–mass spectrometry (LCMS-8040; Shimadzu, Kyoto, Japan). Chromatographic separation was carried out on a C18 column (50 × 2.1 mm, 3.5 μm; Waters). A sample volume of 5 μL was injected into the LCMS system. Two solvents were used for gradient elution: (A) 0.1% formic acid in water and (B) acetonitrile. For the analysis of atenolol and acyclovir, the gradient program was: 0–2 min, 30% B; 2–2.5 min, 30–70% B; 2.5–4.5 min, 70% B; 4.5–5.0 min, 70–30% B; 5.0–6.0 min, 30% B. For the analysis of caffeine, metoprolol, and propranolol, the gradient program was: 0–0.5 min, 40% B; 0.5–1 min, 40–95% B; 1.0–2.5 min, 95% B; 2.5–3.0 min, 95–40% B; 3.0–4.0 min, 40% B. The flow rate was 0.2 mL/min. The ions were used for identification and quantification of the test molecules as follows: atenolol (parent ion m/z, 267.35; product ion m/z, 145.10) and acyclovir (parent ion m/z, 226.20; product ion m/z, 152.10), caffeine (parent ion m/z, 195.20; product ion m/z, 138.10), metoprolol (parent ion m/z, 268.35; product ion m/z, 116.10), and propranolol (parent ion m/z, 260.35; product ion m/z, 116.15). The permeability coefficient (Papp) was calculated from the following equation:
List of the Drugs Selected for Transport Study Across Placental Barrier Model
Apparent permeability factors (Papp, 30 min) across the HUVEC-Tig-STB coculture model. Values are the mean ± SEMs (n = 4).
HUVEC, human umbilical vein endothelial cell; SEM, standard error of the mean; STB, syncytiotrophoblast.
where dQ/dt is the rate of permeation of the compounds across the cells; C0 is the initial concentration of the test compound in the apical compartment at time zero; A is the area of the cell layers.
Statistical analyses
Data are presented as mean ± standard errors of the means. Statistical analyses were conducted with SPSS Statistics version 22 (IBM, Armonk, NY). Comparisons were made among groups by using Student's t-tests or a one-factor analysis of variance followed by Tukey's post hoc analysis. p < 0.05 was considered as statistically significant.
Results
Establishment of a human placental microarchitecture
In this study, a micromesh cell culture technique was used to develop a coculture model representing the human placental barrier (Fig. 2B). The VE-cadherin immunostaining results showed that HUVECs formed a flat cell monolayer mimicking the fetal side (Fig. 2C). Tigs layers, cocultured upon the HUVEC monolayer, acted as the basal layer that separates fetus and maternal side (Fig. 2B, D). TSCs mimicking the maternal side were grown on the apical side of the mesh.
To identify a suitable medium for optimal growth of the cocultures, cells were cultured on tissue culture plates in the recommended media for TSCs or HUVECs as well as in a 1:1 mixture of both media for up to 5 days (Supplementary Fig. S2). HUVECs could grow into a confluent monolayer in a 1:1 mixture of EGM2 and TSC basic medium (Supplementary Fig. S2). Although TSCs were found to be proliferating at a slower rate in the 1:1 mixture of the two media compared with the TSC basic medium alone, at day 4 the TSCs formed a confluent layer (Supplementary Fig. S2). Thus, the 1:1 mixture of media was used for the coculture system. The Tigs also grew quickly in the 1:1 mixture of media (data not shown).
Formation of a tight barrier was monitored by TEER measurements. TEER values of mesh wells without cells was 33.3 ± 1.57 Ω·cm2 (Fig. 2E). For HUVEC monocultures or the coculture of HUVECs and Tigs, TEER values were not significantly increased compared with mesh wells without cells (Fig. 2E). At day 8, TEER values of cocultures with TSCs has considerably increased and reached 174 Ω·cm2 at day 2 of TSC differentiation into STB (Fig. 2E).
Characterization of a human placental microarchitecture
The microengineered human placental model, constructed by the co-culture of HUVECs, Tigs, and TSCs on the mesh, was further characterized by hematoxylin and eosin (HE) staining, OCT nondestructive observation, and immunostaining. The cells were distinguished based on their specific morphologies (Fig. 3A). As HUVEC monolayer is very thin, it was difficult to observe them in a cross-section HE staining image (Fig. 3A). Round shaped TSCs and elongated Tigs were observed in the HE staining image. They were not mixed and maintained independent layers. Tight connection of TSC layers existed upon the multilayers of Tigs. In OCT observation, the signals of Tigs were weaker than TSCs, which showed that Tigs were not tightly connected (Fig. 3B and Supplementary Video S1). The thin HUVEC monolayer was observed in OCT images (Fig. 3B).

Characterization of multilayer structure of cell culture placental barrier model.
We further evaluated the formation of cell–cell junctions to assess structural integrity of the barrier. We focused on the visualization of VE-cadherin expression in the fetal vein endothelial cells and E-cadherin in the trophoblast cells. Immunofluorescence imaging of the multilayer tissue cultured for 8 days clearly showed a network of continuous and well-defined junctional complexes throughout the trophoblast and endothelial layers (Fig. 3C, D). Expression of the junctional proteins was found to be uniform across the entire cell layer on the mesh. The three of cells lines were not mixed and formed separate layer of cells.
We then investigated the stability of the coculture model for extended periods of time. At day 12 of triculture, TEER value was not significantly decreased. Due to the growth of fibroblast, HUVEC sheet of coculture became a little rough based on the VE-cadherin staining images at days 10 and 12 (Fig. 3E, F). As the differentiation progresses, E-cadherin became discontinuous (Fig. 4A). Thus, at day 12, STB layer of coculture was evaluated based on immunostaining of GATA-binding protein 3 (GATA3), which is a nonspecific trophoblast factor coexpressed in the nuclear of TSC and STB. GATA3 staining results showed that TSC sheet of coculture model was maintained up to day 12 (Fig. 3F). These results showed that the coculture model was stable and might be widely used for in vitro placental toxicity evaluation.

Syncytialization of the mesh-cultured TSCs.
Syncytialization of the mesh cultured trophoblast stem cells
As the pregnancy progresses, TSC differentiate and fuse to form a multinucleated STB covering the chorinic villi. Based on our previous studies, forskolin-supplemented media induce syncytialization of TSCs. Concurrent to the fusion of TSCs was a loss of epithelial cell–cell junctions. The localization of adherens junction (E-cadherin) and tight junction (tight junction protein 1 [TJP1]) was analyzed by immunostaining. We found that E-cadherin and TJP1 were clearly expressed at the interface of mononuclear TSCs (Fig. 4A). Microscopic inspection of the TSC treated with forskolin showed significant downregulation of E-cadherin and TJP1 throughout the epithelial layer (Fig. 4A). Interestingly, this reduced expression of junctional proteins did not compromise the structural integrity of the barrier. On the contrary, syncytialization in our model led to improved TEER values, indicative of enhanced barrier function (Fig. 2E).
To convincingly investigate syncytialization of the mesh-cultured TSCs, including changes of morphology, TEER, and correct marker expression, the mono TSC cultures on mesh were prepared. The single undifferentiated TSC layer on mesh incubated with forskolin for 2 days began to undergo cell–cell fusion as illustrated by nuclear aggregation (Fig. 4B). The production of a placental hormone, human chorionic gonadotropin (hCG), is an important functional consequence of trophoblast syncytialization. The production of hCG was examined by immunostaining, and the results revealed the presence of hCG in the STB sheet on mesh, providing evidence for TSC differentiation into functional STB (Fig. 4C).
Single STB layer on the mesh could not be maintained for more than 3 days (Fig. 4D). The TEER value of the STB layer at day 2 significantly increased compared with the single undifferentiated TSC layer (Fig. 4D). However, the TEER value of the STB layer at day 3 significantly decreased compared with those of day 2 (Fig. 4D). At day 3, with the progression of syncytialization, the cells detached from the mesh (Supplementary Fig. S3A). Similar results were observed with the cell fusion on the tissue culture plate (Supplementary Fig. S3B). TSCs were known to form 3D multinucleated STB cysts in the floating culture 15 (Supplementary Fig. S3C). Compared with the floating culture, the cell–mesh connection was stronger; however, this was still not enough to maintain their STB status for a long time. This problem was improved in the coculture model. The TEER value of cocultured sheet was not significantly decreased up to day 12 compared with that of day 10, however, TEER value significantly decreased at day 15 with the progression of syncytialization (Fig. 2E). Unlike single STB sheets, at day 4 of differentiation (day 12), the STB was still observed to be continuous upon the Tigs layers based on GATA3 immunostaining (Fig. 3F). However, TEER value was the highest at day 2 of differentiation (day 10), thus cocultured sheet at day 10 were used for the following experiment (Fig. 2E). The effective differentiation of TSCs into STB need forskolin treatment for 6 days and suspension culture condition (Supplementary Fig. S3C). After forskolin treatment for 2 days in mesh well, not all the cells differentiated into STB and expressed hCG. However, because the STB and part of undifferentiated TSC were in coexistence throughout pregnancy, the coculture cell model treated with forskolin for 2 days can be considered acceptable and reasonable.
We then analyzed the expression of tight junction genes and key trophoblast lineage-specific genes in mesh-cultured single TSC and STB sheet using qRT-PCR analysis. The expression levels of TJP1 and occludin (OCLN) significantly decreased in STBs compared with TSCs (Fig. 4E). Expression levels of TSC marker, tumor protein p63 (TP63), significantly decreased in STBs compared with TSCs (Fig. 4E) The expression levels of STB-specific markers, chorionic gonadotropin subunit beta 3 (CGB3), significantly increased in STBs compared with those in TSCs (Fig. 4E). GATA3 expression level was significantly increased in STBs compared with TSCs (Fig. 4E). Expression levels of STB-specific markers, chorionic somatomammotropin hormone 1 (CSH1), did not show a significant change (Fig. 4E). Taken together, these results demonstrate that mesh culture enables both morphological and functional differentiation of TSCs to reconstitute the syncytium of the placental barrier.
Expression of transporters in the triple coculture model
We examined representative placenta transporters in trophoblast epithelial cells in our triple coculture model using immunostaining and qRT-PCR assay. Transporter for ATP-binding cassette subfamily B member 1 (ABCB1), ATP-binding cassette subfamily C member 3 (ABCC3), and glucose transporter-1 (GLUT1) were visualized in both HUVEC-Tig-TSC and HUVEC-Tig-STB sheets by immunostaining (Fig. 5A, C, E). When the TSCs were incubated with forskolin, the location of these transporters transferred from cell membrane of mononuclear TSC to the membrane of multinuclear STB.
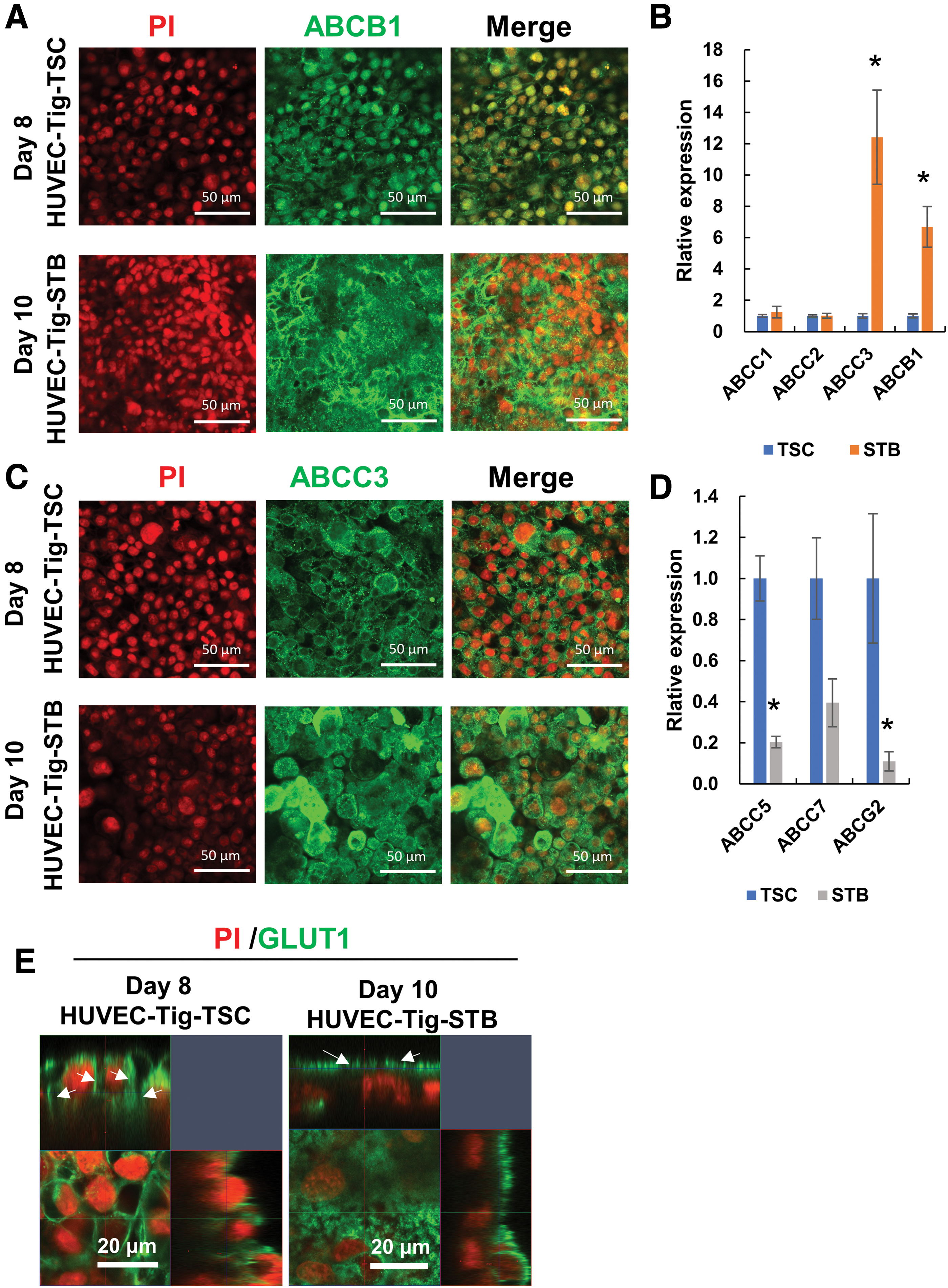
Transporter expression in multilayer cell culture placental barrier model.
Furthermore, to examine the expression of these transporters in the trophoblast epithelial cells, mesh-cultured single TSC and STB sheet were used for qRT-PCR. The expression levels of ABCC3 and ABCB1 significantly increased in STBs compared with TSCs (Fig. 5B). Expression levels of ABCC1, ABCC2, and ABCC7 did not show a significant change (Fig. 5B, D). However, the expression levels of ATP-binding cassette subfamily G member 2 (ABCG2) and ABCC5 significantly decreased in STBs compared with TSCs (Fig. 5D).
Drug permeability of the microengineered placental barrier model
The triple coculture placental model was further characterized by testing drug permeability. For these assays, five compounds with known permeability properties, including passive transport were selected (Table 1). All molecules which enter the fetal system by lipid-mediated free diffusion, such as caffeine, metoprolol, and propranolol, displayed a high Papp (4.5–24.5 × 10−6 cm/s) when measured on our model (Table 1). In contrast, hydrophilic small molecules crossing the cell membrane by passive diffusion, such as atenolol and acyclovir, displayed a low Papp (0.8–0.9 × 10−6 cm/s) (Table 1). We measured the paracellular permeability using FITC–dextran. Our results revealed that the FITC–dextran had very low Papp (0.25 × 10−6 cm/s) (Table 1).
Discussion
One of the major challenges in toxicological research for pregnant woman, including evaluation of drug safety, is to find an appropriate placental barrier model. Any in vitro model to serve as a permeability screen should exhibit restrictive paracellular pathway, a physiologically realistic cell architecture, and functional expression of transporter mechanisms. 24 In the present study, we have constructed and characterized a novel placental barrier model consisting of the triple coculture of TSCs, fibroblast, and HUVECs, with the aim to produce an in vitro tool for research on placental barrier physiology, pathology, and pharmacology.
In the placenta, the fetal circulation is separated from maternal blood by two cell layers, the fetal capillary endothelium and placental trophoblast. In the first trimester, the placental barrier is composed of four different layers, namely STB and CTB covering the villi, the chorionic stroma, and the fetal capillary endothelium. 25 To mimic the placental structure, most published coculture placental barrier models just used two types of trophoblast cell lines and vascular endothelial cell lines. Aengenheister et al., have reported a placental barrier model using BeWo cells and placental venous endothelial cells on a low-absorbing 3 μm porous membrane. 4 More recently, several studies have engineered microfluidics-based placental barrier model, in which the coculture of BeWo/HUVEC or JEG-3/HUVEC were used.9,26,27 During pregnancy, CTBs continuously proliferate to fuse into STB, and at term 44% of CTBs retain to cover the villi. However, in these previous models, choriocarcinoma cell monolayer was used instead of actual CTB and STB bilayer. Our immunostaining results for TJP1 and E-cadherin proteins showed that the STB and part of undifferentiated TSC were in coexistence. Thus, our model was closer to the real human placental barrier structure.
Moreover, the previously reported models did not incorporate the basement membrane part of epithelial and endothelial tissues, which is the extracellular matrix comprising of proteins like laminin. 28 In this study, the fibroblasts were used to support and separate trophoblast and HUVECs. Fibroblasts are known to be producers of laminin, 28 and thus might mimic a physiological basement membrane. Fibroblasts actually help to establish three layers without affecting the electric resistance. This is probably because fibroblasts supported cellular adhesion and function without disrupting cell to cell communication, including the formation of gap junctions. 28 Future studies evaluating the location of basement membrane proteins and gap junctions, and extracellular matrix composition are important for understanding the role of fibroblast. When the TSCs and HUVECs were cultured on the mesh without fibroblasts, HUVEC layer became discontinuous and even fell off from mesh (data not shown). We also cultured TSCs and HUVECs on the tissue culture plate, we found that TSC or HUVECs formed independent colony, but not a two-layered structure (Supplementary Fig. S4A, B). It is reported that different fibroblast lineages and culture condition led to quantitative and qualitative alterations in extracellular matrix composition. 29 Although Tig-1-20 cell line is easily obtained, its rationality is the next consideration. For instance, placental-derived fibroblast line might be a beneficial choice for the in vitro placental barrier model.
Previous in vitro models of the trophoblast barrier required the use of cancer cell lines to make them confluent. These cell line models provide an incomplete model of the placental barrier, since they lack functional STB. In humans, TSCs terminally differentiated syncytium forms the continuous outer lining of the chorionic villi. Different from other physiological barriers in the body that use intercellular tight junction as central regulator in exchange of substance, syncytium acts as a key regulator of substance exchange between the maternal and fetal circulations. However, most in vitro cell models did not use STB as barrier to evaluate the drug transport.4,9,10 For example, Rothbauer et al., have reported that the high seeding density of BeWo cells displayed an increase in TEER from 124 Ω·cm2 at day 1 to 157 Ω·cm2 at 7 days using 12-transwell insert. 10 The TEER value was not significantly increased in their studies, since they have only considered tight junctions with sharp cell-to-cell boundaries of BeWo cells, but not their acquisition of STB-like phenotype. In our models, trophoblast part used hiPSC-induced TSCs that showed the capacity to differentiate into STBs. Forskolin-treated STB showed the induction of hCG expression along with lack of TJP1 and E-cadherin expression by immunostaining. Furthermore, differentiation of TSCs enabled the significant increase of TEER from 96 Ω·cm2 at day 8 to 174 Ω·cm2 at day 10. Our results demonstrate that our models morphologically and functionally mimic syncytium of the human placental barrier, compared with the widely used choriocarcinoma-derived cell line model. In this study, HUVECs-Tig-STB showed tight barriers with net TEER values 140 Ω·cm2 (mesh area = 0.1256 cm2). A human primary trophoblast monolayers with net values around 300–400 Ω·cm2 (Transwell area = 1.12 cm2). 30 Because of the difference of culture area and substrate, it was difficult to compare our model with the primary trophoblast monolayer model.
Another vital structural feature of the placental barrier is microvilli, which increases the overall surface area available for placental transport and enables highly efficient exchange of substance. 26 The effect of fluid mechanical forces have recently been shown to trigger microvilli formation in BeWo cells.26,31 In contrast, BeWo cells grown in Transwell for the same period do not easily express microvilli. 26 Nishiguchi et al., reported primary trophoblast cultured in Transwell-developed microvilli. 32 The apical surface of the STBs cultured in our coculture model developed microvilli-like feature without the use of microfluidic stimulation (Fig. 4A). Microvilli-like feature was also observed on the cell surface of STBs cultured on the dish (Supplementary Fig. S3D). Although we used the scanning electron microscopy method to observe these microvilli-like part, it was still difficult to confirm 3D structure of them, which might be due to complication of the coculture cell structure.
In the placenta, ABCA-G transporters are believed to play major roles in the active transport of various endogenous metabolites and in moving various xenobiotics and drugs out of the placental trophoblast. 33 Previous report showed that the expression level of ABCG2, ABCC1, 2, 3, 5, 7, and ABCB1 of the villous STB is higher compared with CTB in human placenta. 34 In our studies, the result for ABCC3 and ABCB1 was consistent with the previous report, but not ABCG2, ABCC5, and 7. This is probably because there is not enough villous STB formation in our model.
Transmembrane transport through the placental barrier is related to the physiochemical properties of compounds. In our study, five compounds were chosen as model compounds, because they are small molecules with major differences in physiochemical properties. All compounds are classified as freely diffusing across the placental barrier. In the ex vivo perfusion system or BeWo monolayer model, the compounds were transported in the following rank order: caffeine ≥ hydrophobic substance > hydrophilic substance, in terms of the apparent permeability coefficient.35,36 In our coculture system, the rank order of the compounds were as follows: caffeine > metoprolol ≥ propranolol > atenolol ≥ acyclovir. These results showed an excellent correlation with the ex vivo results. Moreover, caffeine displayed a high Papp (3.2 × 10−4 cm/s) in BeWo monolayer 36 compared with our model system (24.5 × 10−6 cm/s). The transfer rate using ex vivo the perfusion system was much faster compared with the BeWo monolayer. 36 Metoprolol, propranolol, atenolol, and acyclovir also displayed similar tendency. Although the transfer rate is not the same, our results from the in vitro studies showed a good correlation with the ex vivo results. Our model can serve as a valuable tool, necessary to predict the fetal exposure to potentially harmful substances that may affect fetal development. Metoprolol, propranolol, atenolol, and acyclovir as drugs are widely used, however, these drugs have been restricted due to concerns about maternal or fetal safety. 37 Furthermore, these compounds were commonly selected to evaluate the other human barrier, such as blood–brain barrier model and gastrointestinal tract models.38,39 Therefore, we first selected these common drugs to confirm function of our placental barrier model. Future studies evaluating more compounds, which across the barrier by facilitated diffusion or active transport, are important to test the availability of our model as a drug screen tool. Furthermore, to keep the maternal or fetal safety, the safe dose of compounds, such as caffeine concentration (1.03 mmol/day), are suggested. 27 Since larger dose might induce higher transfer concentration, the dose–response of compounds are needed to consider.
Our model integrated functional TSC derived from human iPSCs, which was an advantage over the widely used choriocarcinoma-derived cell lines. 40 Previous reports by Okeyo et al. have shown that the hiPSCs (clone TIG1–4F hiPSC from lung fibroblasts, provided by Dr. Takashi Tada of Kyoto University, Kyoto, Japan) can differentiate into TSC-like cells.19,20 In this study, we used the TSCs derived from the hiPSC line ChiPSC22, which is more ordinary and commercially available than the above hiPSC. Furthermore, we have been tried to obtain TSCs using the other common hiPSC lines 201B7 and 253G1, however, these hiPSC lines did not effectively differentiate into trophectoderm.
In summary, we established and characterized an in vitro reconstituted placental barrier model using hiPSC-derived TSCs. The use of micromesh technique enabled us to develop a layered microstructure that was closer to the realistic physiological structure. The models may provide a useful platform for in vitro investigation of the structural and functional organization of the placental barrier. The models provide new opportunities to create cell-based pharmaceutical screening assays for evaluating the placental transfer of pathogens, drugs, chemicals, and environmental toxins. Our current model, however, still leaves room for improvement to realize its full potential. The system used in this study was static, and to address this limitation, the design of microfluidics-based approaches will be more effective for improved physiological and biological relevance.
Footnotes
Acknowledgments
The authors thank Dr. Fumiyoshi Yamashita (Kyoto University) and Dr. Masahiro Tsuda (Kyoto University) for LCMS measurement of samples, the technical staff (SCREEN Holdings Co., Ltd.) for obtaining OCT images, and Dr. Kennedy Omondi Okeyo (Kyoto University), Dr. Masao Washizu (Tokyo University), Dr. Takeshi Hori (RIKEN Institute), Ms. Hiromi Ito (RIKEN Institute), and Mr. Nobutaka Tani (RIKEN Institute) for valuable suggestions.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by grants from the Japan Society for the Promotion of Science (JSPS; KAKENHI grant No. 18K12071) and the “Compass to Healthy Life” Research Complex Program of the Japan Science and Technology Agency (JST), and the Japan Agency for Medical Research and Development (AMED, grant number: JP20bm0404030).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.