
Abstract
There is a constant need for improving embryo culture conditions in assisted reproduction. One possibility is to use mesenchymal stem/stromal cells derived from menstrual blood (mbMSCs), with an endometrial origin. In this study, we sought to analyze the expansion of mouse embryos in a direct coculture model with mbMSCs. Our results showed that after five passages, mbMSCs presented a spindle-shaped morphology, with surface markers that were comparable with the normal mesenchymal cell phenotype. mbMSCs could differentiate into adipogenic and osteogenic lineages and secrete angiopoetin-2 and hepatocyte growth factor. The coculture experiments employed 103 two-cell-stage embryos that were randomly divided into two groups: control (n = 50), embryos cultured in GV-Blast medium, and cocultured mbMSCs (n = 53), embryos cocultured with GV-Blast and mbMSCs. Typically, two to three embryos were placed in a well with 200 μL of culture medium and observed until developmental day 5. After 5 days, the cocultured group had more embryos in the blastocyst stage (69.8%) when compared with the control group (30%) (p < 0.001). It was also found that nearly 57% of blastocysts in the cocultured group reached the hatching stage, while only 13% achieved this stage in the control group (p < 0.001). Analyses of cultured mbMSCs and growth media, in the presence or absence of an embryo, were also performed. Immunofluorescence detected similar levels of collagen I and III and fibronectin in both mbMSCs and cocultured mbMSCs, and similar amounts of growth factors, VEGF, PDGF-AA, and PDGF-BB, were also observed in the conditioned medium, regardless of embryo presence. The present study describes, for the first time, an easy, noninvasive, and autologous method that could potentially increase blastocyst growth rates during assisted reproductive procedures (i.e., in vitro fertilization). It is proposed that this mbMSC coculture strategy enriches the embryonic microenvironment and promotes embryo development. This technique may complement or replace existing assisted reproduction methods and is directly relevant to the field of personalized medicine.
Impact statement
The study demonstrates a novel and potentially personalized assisted reproduction approach. The search for alternative and autologous methods provides assisted reproduction patients with a better chance of a successful pregnancy. In this study, mesenchymal cells derived from menstrual blood resembled the outside uterine surface and could potentially be employed for improving embryo outgrowth. Our protocol enriches the embryonic microenvironment and facilitates high-quality single-embryo transfer.
Introduction
Infertility, a complex disease, affects 50–80 million people worldwide (WHO) and has important implications for the couple's quality of life. 1 Women particularly may face psychosocial consequences that are disturbing for their health.1,2 Therefore, the use of assisted reproductive techniques, including in vitro fertilization (IVF), has been constantly refined. IVF is employed worldwide to facilitate pregnancy. This is a safe technique and allows the birth of thousands of healthy children every year.3–5 However, for several couples who need IVF, there is no clear cause of infertility and they have to repeat cycles without success or give up the idea of having their own child.6,7 With increasing use of personalized medicine, new tools are being developed to improve the chances of successful pregnancies.
The culture of IVF embryos has long been used as a protocol before uterine transfer. 8 For decades, investigators have been focusing on improving the conditions of embryo culture since they are crucial for its development. 9 It is well known that embryos that successfully reach the blastocyst stage have better pregnancy and implantation rates.10–13 In the female reproductive tract, the presence of a large repertoire of cytokines and growth factors causes them to act together to orchestrate the ideal development of the embryo. In this context, it is worth mentioning that stromal and/or epithelial cells derived from the endometrium play a crucial role in this process, involving proliferation of ICM blastomeres, 14 differentiation and proliferation of trophoblast cells,15,16 blastocyst survival, 17 implantation, 18 and embryo development as a whole.15,19
To mimic the in vivo complex microenvironment in vitro, the use of feeder layers, comprising animal cells,20,21 human cells,20,22–24 or biological/synthetic compounds,25–28 to stimulate blastocyst development in a coculture system has been showing efficient results, supporting and improving the cell growth rates.
In this scenario, we used human menstrual blood-derived mesenchymal stem/stromal cells (mbMSCs) as a feeder layer to improve the condition of embryo development in IVF cycles. Among the mesenchymal cell sources, menstrual blood was selected for two reasons: (i) it is derived from shedding of the endometrial lining, specifically, the functional layer, which contains highly proliferative cells used to prepare the uterus for implantation; and (ii) it can be collected from the same patient before IVF treatment, avoiding any possible immune reaction issue.
Since their discovery in 2007, mbMSCs have demonstrated outstanding advantages, such as an easy and noninvasive collection and isolation process, high proliferative capacity, and multilineage differentiation potential.22,29–31 In consonance with others, our group showed that mbMSCs, isolated from menstrual blood-derived mononuclear cells, expressed the surface markers of mesenchymal stem cells, such as CD73, CD90, and CD105, and are negative for hematopoietic cell markers, including CD34 and CD45.30–32 It was also demonstrated that mbMSCs were able to support human embryonic stem cells in an undifferentiated state. 22
In this study, we hypothesized that cross talk between the embryo and mbMSCs would improve embryo development. Our data showed (for the first time) that mbMSCs improved the embryo morphology characteristics and this procedure is compatible with clinical applications of personalized medicine.
Materials and Methods
Ethical approval for human samples and animal experiments
The use of human menstrual blood was approved by the Clementino Fraga Filho University Hospital's Ethics Committee (Protocol: 52627716.7.0000.5257). All donors provided signed informed consent in accordance with the principles of the Declaration of Helsinki.
The use of mouse embryos was carried out in accordance with the Guide for the Care and Use of Laboratory Animals [DHHS Publication No. (NIH) 85-23, revised in 1996, Office of Science and Health Reports, Bethesda, MD], and all animal protocols were approved by the Ethics Committee on the Use of Animals in Scientific Experimentation (Health Sciences Centre of the Federal University of Rio de Janeiro), under protocol number 108/16.
Isolation and culture of mesenchymal stem/stromal cells derived from menstrual blood
mbMSCs were isolated by direct plating of menstrual blood and maintained as previously described by us.31,33 Briefly, menstrual blood was collected from healthy, reproductive-age, and volunteer women (n = 8), using a proper collector pot containing 5 mL of 1 × phosphate-buffered saline (PBS, pH 7.2; LGC Biotecnologia), 100 U/mL penicillin, and 100 mg/mL streptomycin (Gibco). Samples were washed twice by centrifugation and resuspension in 5 mL of PBS. Cells were cultured in (4.5 g/L) high-glucose Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich), supplemented with 20% fetal bovine serum (FBS; Gibco), 2 mM
Immunophenotypic characterization of mbMSCs
The mbMSC protein profile was evaluated by flow cytometry. Cells were detached from the culture plate by 0.25% trypsin-EDTA (Sigma). The staining solution (PBS 0.5% FBS) and Human BD Fc Block (Cat. No. 564219; BD Biosciences) were used to avoid nonspecific antibody binding. A total of 3 × 105 mbMSCs were incubated for 20 min in the dark at 4°C with the following antibodies according to the manufacturer's instructions: CD14 PE (Cat. No. 39110; BD Biosciences), CD19 APC (Cat. No. 555415; BD Pharmingen), CD31 PE (Cat. No. 555446; BD Pharmingen), CD34 PE (Cat. No. 348057; BD Biosciences), CD45 FITC (Cat. No. 555482; BD Pharmingen), CD73 APC (cat 560847; BD Pharmingen), CD90 PE-Cy5 (Cat. No. 555597; BD Pharmingen), CD105 PE (Cat. No. 560839; BD Pharmingen), and HLA-DR PE-Cy5 (Cat. No. 551375; BD Pharmingen). Unstained cells were used as autofluorescence controls. The cells were acquired in ACCURI C6 (BD Biosciences) and analyzed using FlowJo, version 10, software.
mbMSC differentiation assays
For differentiation assay, cells (P5) were cultured for 21 days under specific medium. Adipogenic differentiation medium comprised DMEM-high glucose (4.5 g/L; Gibco) supplemented with 10% FBS (Gibco), antibiotics (100 IU/mL penicillin and 100 mg/mL streptomycin; Gibco), 10−7 M dexamethasone (Sigma-Aldrich), 2.07 μM insulin (Sigma-Aldrich), and 0.45 mM isobutyl-methylxanthine (Sigma-Aldrich). Oil Red O staining (0.2%; Sigma-Aldrich) was used to identify cytoplasmic lipid vacuoles. With regard to osteogenic differentiation, the medium comprised DMEM-high glucose (4.5 g/L) supplemented with 10% FBS, antibiotics (penicillin 100 IU/mL and streptomycin 100 mg/mL), 10−7 M of dexamethasone (Sigma-Aldrich), 0.5 μM of ascorbic acid (R&D Systems), and 10 mM of β-glycerophosphate (Sigma-Aldrich). Alizarin Red (1%; Sigma-Aldrich) staining was performed to identify calcium deposits in the extracellular matrix.
Cytokine quantification in mbMSC-conditioned medium by LEGENDPlex
Cytokine production in mbMSC-conditioned medium was analyzed by the LEGENDplex Human Growth Factor Panel kit (Cat. No. 740180; Biolegend) according to the manufacturer's instructions. This kit allows quantifying 13 cytokines (ANG-2, VEGF-A, EGF, EPO, b-FGF, G-CSF, GMCSF, HGF, M-CSF, PDGF-AA, PDGF-BB, SCF, and TGF-α) simultaneously in the same sample. The tubes for acquisition were prepared with 25 μL of sample, 25 μL of mixture of beads, 25 μL of detection reagent, and 25 μL of assay buffer solution. The same procedure was performed to obtain the standard curve. However, in this case, the sample was replaced by the standard solution and the assay buffer replaced by fresh culture. The tubes were kept on a shaking plate at 1000 rpm, for 2 h, at room temperature, protected from light. Then, 25 μL of streptavidin–phycoerythrin (PE) (substrate for enzyme conjugated with fluorochrome) was added in each tube and these were maintained again on a shaking plate at 1000 rpm, for 30 min, at room temperature, protected from light. Then, tubes were centrifuged at 1000 g for 5 min. The supernatant was then carefully removed and immediately 200 μL of wash buffer was added. This procedure was repeated two more times and then the samples were read in ACCURI C6. The data obtained were analyzed using Biolegend's LEGENDplex software.
Feeder layer procedure
One day before coculture, mbMSCs (∼80% confluence) were detached from the culture flasks by 0.25% trypsin-EDTA (Sigma-Aldrich). Then, 1.5 × 104 cells were resuspended in 200 μL of GV-Blast (Ingámed®) supplemented with 20% FBS, plated on 96-well plates (Corning), and maintained at 37°C in a 5% CO2 incubator.
Animals
A total of 16 males and 48 females of the FVB mouse strain, aged 4–5 weeks, weighing ∼20 g, were obtained from the Health Sciences Centre–Federal University of Rio de Janeiro/Brazil. Animals were housed under controlled temperature (23°C) with daily exposure to a 12-h light–12-h dark cycle with free access to water and standard rodent chow.
Superovulation protocol
For follicular recruitment, female mice of reproductive age (4–5 weeks) received (intraperitoneally) 5 IU of equine chorionic gonadotropin—Novormon®, Ourofino. After 46–48 h, 5 IU of human chorionic gonadotropin (hCG)—Vetecor®, Hertape Calier—was given intraperitoneally and females were placed with fertile males. Copula was confirmed the next morning by the presence of a vaginal plug.
Collection of embryos by uterine tube flushing
After 48 h of hCG administration, female mice were anesthetized with isoflurane inside a chamber to reach an unconscious state and euthanasia was performed by cervical displacement. A skin and peritoneal incision was made to visualize the abdominal cavity and expose and remove the ovaries and uterine tubes. 34 The dissected uterine tubes were placed in a 100-mm Petri dish containing ∼0.5 mL drops of modified human tubal fluid medium (Irvine Scientific) supplemented with 10% FBS. The flushing was done under a stereoscope with a 26G syringe. During this procedure, rupture and flushing of the tubal structures were performed and it was possible to visualize the two-cell embryos after decanting them to the bottom of the drop, which was recorded as E2.
Embryo culture on mbMSC feeder layer
In eight repetitions of the experiment, mouse two-cell embryos (n = 103) were randomly distributed into two groups: control group—embryos (n = 50) were cultured in GV-Blast medium supplemented with 20% FBS, and the coculture mbMSC group—embryos (n = 53) were grown over mbMSCs in GV-Blast medium supplemented with 20% FBS. Embryos in the two-cell stage (E2) were transferred to a 96-well polystyrene plate (Corning®) with or without mbMSCs (control group and cocultured mbMSC group) and it was considered culture day 0 (D0). The culture was maintained for 3 days (D3) at 37°C in a 5% CO2 incubator.
Development assay
At D0 and D3, cultured embryos were photographed under light microscopy using the EVOS XL Core Cell Imaging System (Thermo Fisher Scientific). Morphological parameters were analyzed based on the images captured.
Embryo development evaluation
At the end of D5, embryos were classified according to the stage of embryonic development. Embryos that showed evolution delay or stopped development were characterized as blocked embryos, while those presenting initial characteristics of the blastocyst stage, such as blastocele, inner cell mass, and trophectoderm, were classified as blastocysts. The blastocyst rate of each group was registered, as well as the percentage of the same in each experiment, according to the number of embryos collected in each procedure.
Blastocyst rates
Blastocysts were quantified on D5 and analyzed based on their morphological features observed on the captured photographs. In addition, they were classified according to their degree of development: the initial blastocyst—considered as having a blastocele occupying more than 50% of the volume of the embryo and thick zona pellucida; expanded blastocysts—those that presented a blastocele occupying 100% of the volume of the embryo, significant increase of the embryonic volume, and decrease of the thickness of the zona pellucida; hatching blastocysts—those that presented partial exit of the trophectoderm through the zona pellucida; and hatched blastocysts—those that were completely outside the delimitation of the zona pellucida. The degree of expansion and development of blastocysts on the fifth day of culture was used as a parameter of embryo quality.
Detection of growth factors in conditioned medium of the cocultured mbMSC group
The analysis of growth factors, PDGF-AA, PDGF-BB, and VEGF, in the mbMSCs and cocultured mbMSC culture medium was performed with the MILLIPLEX-MAP Human Cytokine/Chemokine Magnetic Bead Panel—Immunology Multiplex Assay kit (HCYTOMAG-60K; Merck Millipore), following the manufacturer's recommendations. Briefly, the test was performed on a 96-well plate provided by the manufacturer, where quality controls and standards were tested in duplicate (50 μL in total) and samples were analyzed in triplicate (25 μL). The plate with samples and magnetic beads was read on the MAGPIX® System equipment (Merck Millipore). The analyses were performed using the Luminex xPONENT® for MAGPIX® software, v 4.2 (Luminex Corp.). For each reaction well, the MAGPIX Luminex® platform reports the median fluorescence intensity for each of the analytes in the samples. The levels of each analyte are then calculated by the software using the results of the standard curve.
mbMSC extracellular matrix characterization
The mbMSCs (no contact with embryos) and cocultured mbMSCs (mbMSCs used as a feeder layer to embryos) were fixed with 4% paraformaldehyde (Sigma-Aldrich) for 20 min at room temperature and then washed with PBS. Blocking of nonspecific sites with a solution containing PBS and 3% bovine serum albumin (BSA; Sigma-Aldrich) was done for 1 h and they were incubated with primary antibody for 18 h (fibronectin [1: 400, F3648; Sigma-Aldrich], collagen I [1:100, ab292; Abcam], and collagen III [1:200, ab7778; Abcam]). After this period, the following were performed: washes were performed with PBS, incubation with 1% BSA and the secondary antibody (1: 500, Alexa Fluor 488, A11034; Thermo Fisher Scientific) for 2 h, washing with PBS, and adding of DAPI (4′6′-diamidino-2-phenylindole; Sigma-Aldrich) for 5 min. After a last wash with PBS, the plate was analyzed by ImageXpress micro XLS automated fluorescence microscopy (Molecular Devices), ImageXpress® Micro XL (Software: Meta Xpress 6.0). For this purpose, the 10 × objective was selected and digital focus was used to acquire images, with lenses combined with DAPI excitation filters for the nucleus and FITC for type I and III collagen and fibronectin. Segmentation of images was done by positivity or negativity for marking with different dyes and antibodies. Total cell counts were performed by DAPI labeling and fluorescence intensity for the matrix proteins in each well of the culture plate with the Cell Scoring module. Subsequently, to avoid an erroneous analysis due to the number of cells, the fluorescence intensity of each well was normalized by the total number of cells.
Statistical analysis
Eight independent experiments were carried out. Multiple comparisons and chi-square test were used to evaluate blastocyst development. All the other results are expressed as mean ± standard error of the mean and evaluated using unpaired Student's t-test. A p-value of less than 0.05 was considered statistically significant.
Results
mbMSC feeder layer characterization
The standards of the International Society for Cell Therapy (ISCT) were used to characterize the mbMSCs as mesenchymal cells. 35 The mbMSCs composed an adherent cell culture, presenting a fibroblast-like shape (Fig. 1A) with the ability to differentiate into adipocytes (presence of lipid vacuoles in the cytoplasm) and osteocytes (deposits of calcium in the extracellular matrix) when cultivated under specific conditions, as shown in Figure 1E–H. The mbMSC immunophenotype was evaluated by flow cytometry and these cells expressed mesenchymal markers (CD73, CD90, and CD105) in more than 90% of the culture and were negative for hematopoietic (CD19, CD14, CD34, CD45, and HLA-DR) and endothelial (CD31) antigens, as shown in Figure 1B and C. These results are in accordance with our previous results 31 and collectively meet the criteria of the ISCT for MSC classification standards.
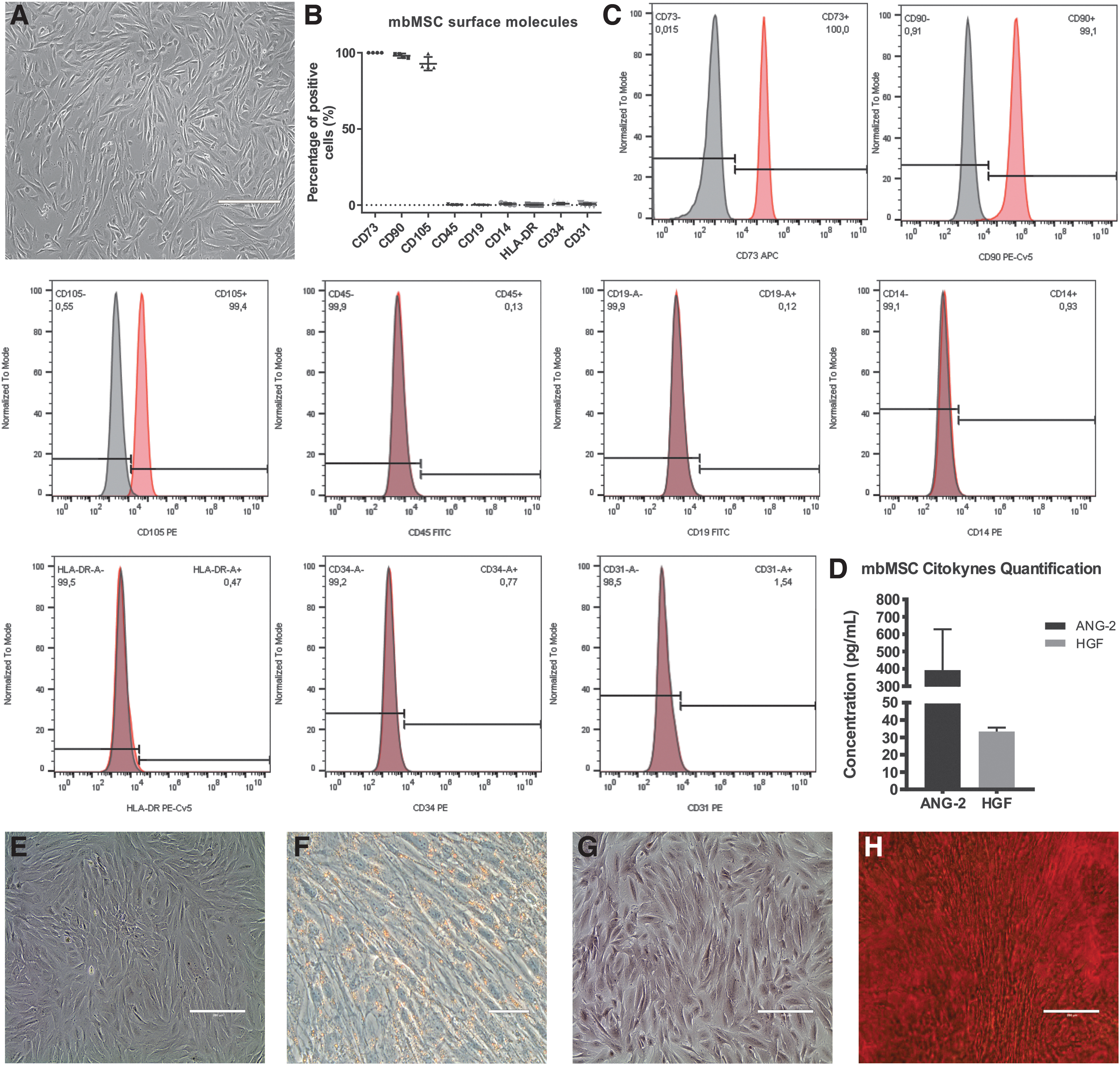
mbMSC characterization.
Cytokine quantification in mbMSC-conditioned medium by LEGENDPlex
To identify the growth factors secreted in the mbMSC-conditioned medium, a panel with 13 cytokines, which might be involved in embryo development, angiogenesis, and immunomodulation, was used to evaluate the levels of these associated biological factors. Typical angiogenic factors were observed in these cultures, including hepatocyte growth factor (HGF) and angiopoietin-2 (ANG-2), the latter in higher concentrations. However, the cytokines, VEGF-A, EGF, EPO, b-FGF, G-CSF, GMCSF, M-CSF, PDGF-AA, PDGF-BB, SCF, and TGF-α, were below the detection limit in the culture media (Fig. 1D).
mbMSC feeder layer morphology
To evaluate whether or not cell morphology could be modified after culture in a different medium, mbMSCs were plated and maintained with either DMEM or GV-Blast, supplemented with FBS, for 96 h. Figure 2A and B shows that mbMSCs grow as monolayers on 96-well culture plates and present fibroblast-like morphology, with thin and elongated adherent cells, in both conditions.
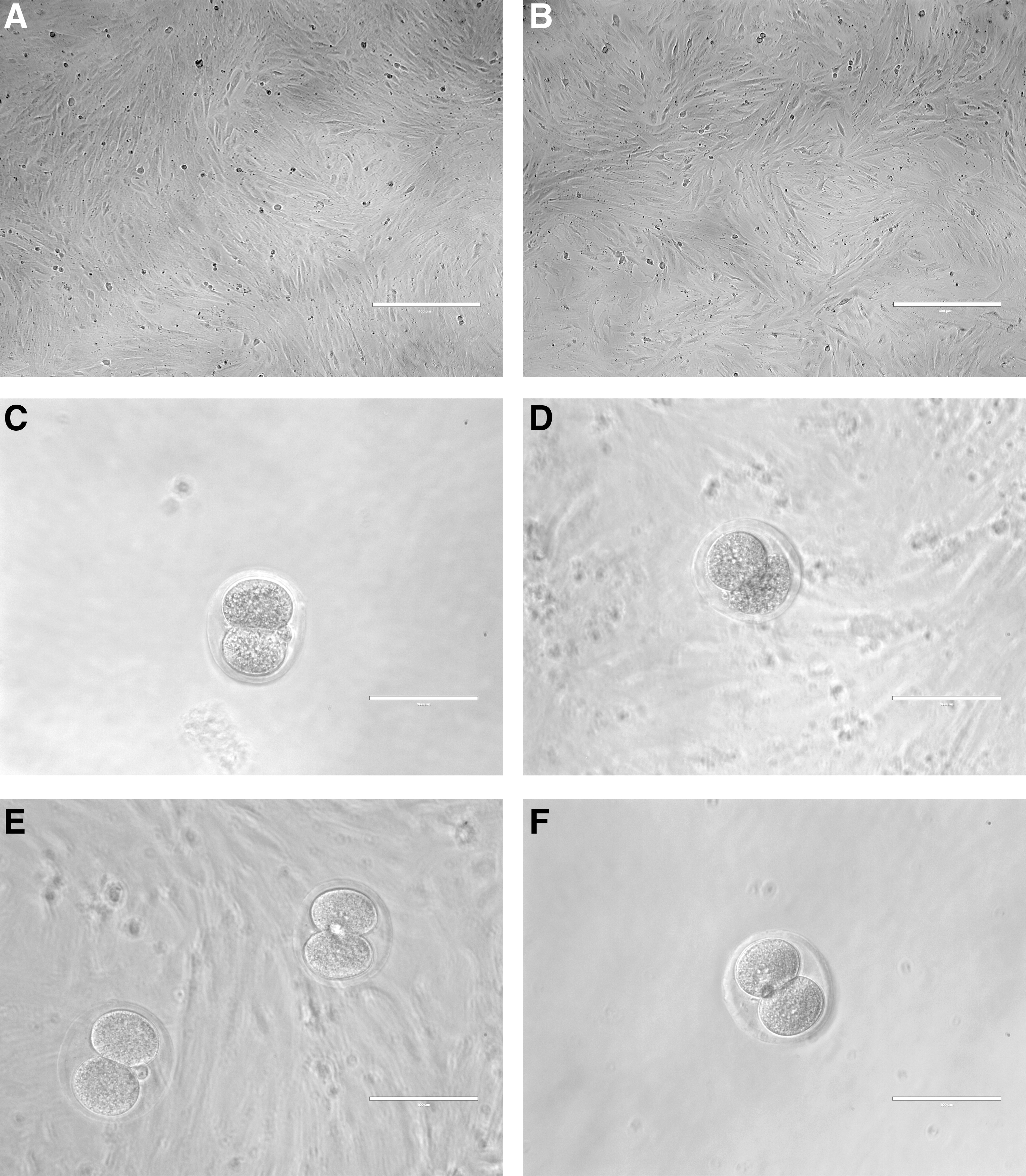
Feeder layer and embryo morphology.
Embryo collection
After the superovulation in mice, 103 embryos were collected in the two-cell stage (E2), as shown in Figure 2C. All embryos that did not present complete cell morphology were discarded (Fig. 2D). Figure 2E and F illustrates embryos in E2 transferred either to the mbMSC feeder layer culture (Fig. 2E) or GV-Blast medium control well (Fig. 2F).
In vitro development
On the fifth day of development, embryo morphology was evaluated and classified as blastocyst or blocked embryos. The number of blastocysts was higher in embryos cocultured over mbMSCs (69.8%) compared with GV-Blast (30%). Therefore, the percentage of blocked embryos was higher in the GV-Blast condition (70%) when compared with the cocultured group (30.2%), as shown in Figure 3A and Table 1. According to the representative image in Figure 3B, the blastocyst classification includes embryos that show a well-defined space of a blastocele and clear distinction between the inner cell mass and the trophectoderm. In contrast, the classification of blocked embryos includes embryos that remain in the early stage of cleavage or present signals of cellular degeneration.

Embryo development evaluation.
Number and Percentage of Blastocysts and Blocked Embryos in Each Condition on the Fifth Day of Development
p < 0.001.
mbMSCs, menstrual blood-derived mesenchymal stem/stromal cells.
Blastocyst development evaluation
On the fifth day of development, blastocysts were classified into four different stages of development. Considering that the hatching process, a natural stage of embryonic development, consists of the movement of contraction and expansion of the blastocyst with the objective of leaving the pellucida zone, the partially hatched blastocysts are considered less developed than those that spent substantial energy to complete the total exit from the pellucida zone and are now ready for the next stage of the embryo's metabolism and implantation in the uterus. Figure 4A and Table 2 show that the majority of embryos in the control group were at the initial blastocyst stage (60.0%), whereas the cocultured group with mbMSCs presented mostly blastocysts in the hatching stage (54.1%) and was also the only group that showed an embryo in the hatched stage (2.7%). Representative images of initial, expanded, hatching, and hatched blastocysts are illustrated in Figure 4B–E, respectively.

Classification of Blastocyst Stage in Each Condition on the Fiftieth Day of Development
p < 0.001.
Embryo's influence on mbMSC-secreted growth factors
To evaluate whether or not the embryos could modulate the growth factors secreted by mbMSCs, conditioned media from mbMSCs with no contact with embryos and cocultured mbMSCs were collected for multiplex analysis. Secretion of PDGF-AA, PDGF-BB, and VEGF showed no difference between mbMSC-conditioned media cultured with or without embryos, as shown in Figure 5.

mbMSC growth factor secretion in the presence or absence of mouse embryos.
Extracellular matrix protein detection in mbMSCs
To evaluate the impact of embryo's presence on extracellular matrix protein expression, mbMSCs cultured with or without embryos were submitted to immunofluorescence assay for collagen I and II and fibronectin. The presence and distribution of collagen I and II and fibronectin within individual cells showed that there was no difference between mbMSCs cultured with or without embryos (Fig. 6). The presence of these extracellular matrix proteins might improve embryo outcome.
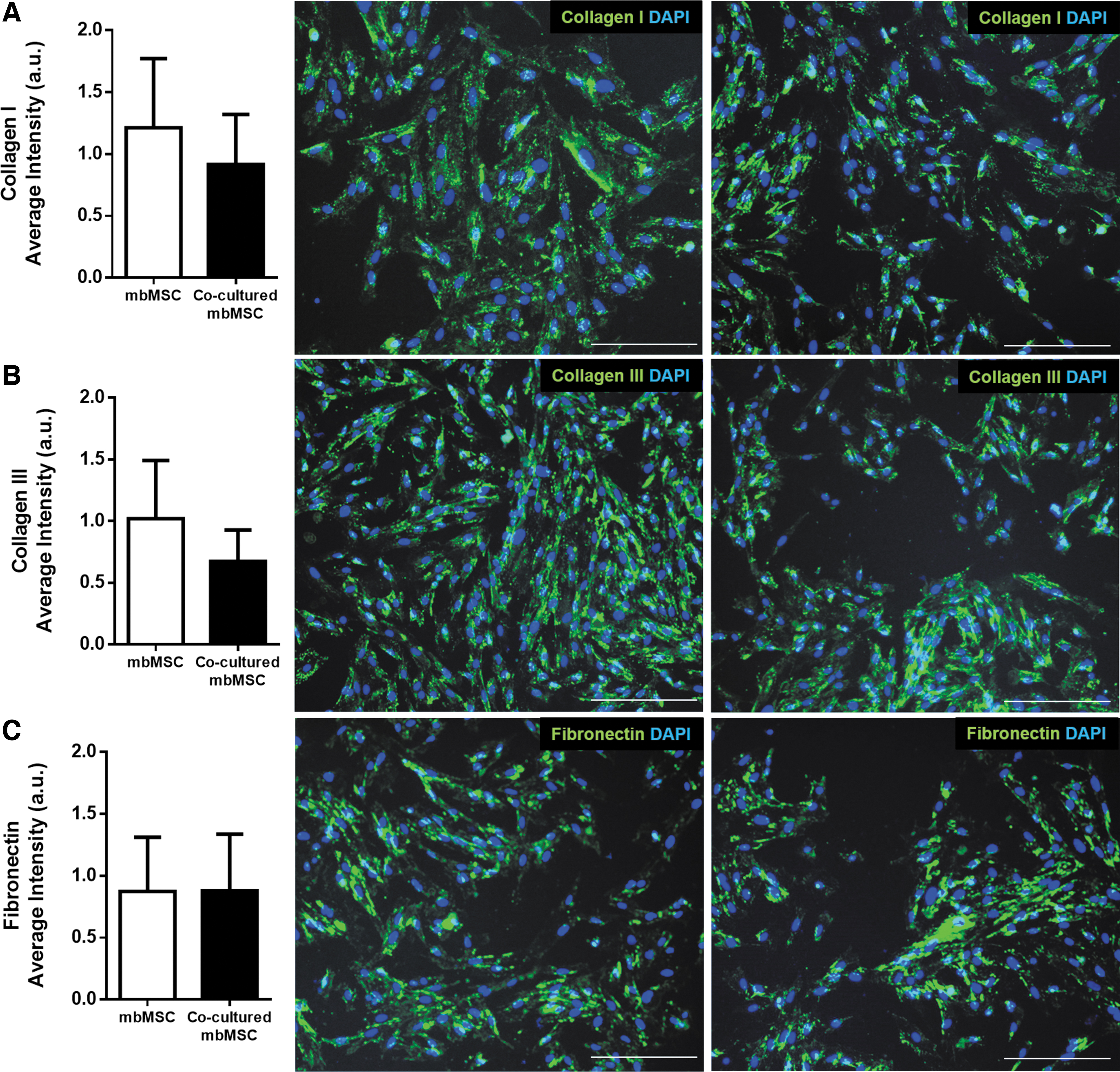
Extracellular matrix protein analysis. Quantification and representative image of the immunofluorescence of
Discussion
The literature review shows that a woman has an increased chance of getting pregnant when a blastocyst is transferred, compared with the transfer of cleavage-stage embryos. 12 It is known that an extended embryo culture up to fifty days needs special conditions that satisfy all requirements of the embryo. In this study, we tested the use of mbMSCs as embryo feeders in a way so as to increase blastocyst quality and quantity. First, we found that coculture affects the quality of the embryos. Cocultured embryos showed early compaction at the morula stage and an increase of visible blastocyst hatching compared with the control group.
Other authors have shown different systems of coculture to improve the embryo environment.20,21,36 In this study, we chose to test mbMSCs because menstrual blood obtention does not face ethical and technical problems since it does not require donor hospitalization and invasive procedures and brings no discomfort to the donor. In addition, mbMSCs do not have teratogenic potential and have a capacity for rapid expansion in culture. 37 Another advantage is the reduced risk of rejection of mbMSCs, which belong to the patient, enhancing the chances of autologous transplant success and still avoiding the use of immunosuppressive drugs. At present, the therapeutic potential of mbMSCs has already been demonstrated in several types of diseases in preclinical research, including Duchenne muscular dystrophy, 38 myocardial infarction,39,40 liver disease,41,42 stroke,43,44 diabetes, 45 and ovarian failure.46,47 These positive results are crucial for future clinical applications of mbMSCs in tissue repair and regenerative medicine. Moreover, mbMSCs have already been reprogrammed to a pluripotent state, originating induced pluripotent stem cells, which are extensively used in personalized medicine studies since these cells are able to differentiate into cells of the three germ layers and have the donor's genetic information.48,49
Additionally, we demonstrated that mbMSCs secrete the cytokines, Ang-2 and HGF, under baseline conditions, in consonance with data from studies by Meng et al. 29 and Liu et al. 50 Both cytokines participate in development and implantation of the embryo. Ang-2 seems to act complementarily to the VEGF system and contributes to the later stages of angiogenesis. HGF stimulates trophoblast cell outgrowth during implantation and influences placental development in mice. 51 Moreover, we observed the secretion of angiogenic factors such as VEGF, PDGF-AA, and PDGF-BB in the conditioned medium of both groups analyzed—mbMSCs only and cocultured mbMSCs. VEGF is involved in embryo–endometrium interactions by regulating increased endometrial vascular permeability and endothelial cell proliferation in implantation sites in rats. 52 VEGF establishes a receptive endometrium to support blastocyst implantation and trophoblast invasion, while PDGF-A is likely to be involved in the regulation of cell growth and differentiation during embryo development as PDGF-A and its receptor (PDGFR-a) have been found to be expressed in mouse eight-cell, morula, and blastocyst stages. 53 It is not yet known whether the mechanisms underlying the effects of these growth factors are related, but it could be speculated that both growth factors influence the metabolic activity of inner cell mass and/or trophectoderm cells, leading to an improved rate of cell division and blastocyst development.
We also looked into the matrix proteins such as collagen I and III and fibronectin (important proteins in the nidation process54,55) in mbMSCs and cocultured mbMSCs. Our data showed that regardless of the presence of the embryo, these extracellular matrix proteins do not alter their expression, which may support embryo development, explaining the higher rate of hatching in the presence of mbMSCs.
Finally, we provide evidence, for the first time, of an easy, noninvasive, and autologous method to increase blastocyst rates during IVF treatment. We are aware that the usage of mbMSCs on a daily basis in IVF clinics is still difficult. However, in individual cases, where embryos are unable to develop in vitro, the use of the patient's own cells could be an important tool for personalized reproductive treatment.
Footnotes
Acknowledgments
The authors want to thank Fernando Marques Guimarães e Livia Guapyassú Siqueira for the assistance during the execution of some protocols. The authors would also like to thank the technicians from the National Center of Structural Biology and Bioimaging of Federal University of Rio de Janeiro for the immunofluorescence microscope and technical support.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Center of Science and Technology for Regenerative Medicine/REGENERA grant no. 465656/2014-5, National Research Council/CNPq grant no. 401816/2016-8, Rio de Janeiro Research State Foundation/FAPERJ grant no. E-26/202.963/2017, Brazilian Council for Scientific and Technological Development (CNPq) grant no. 465656;2014-5, and the Coordination for the Improvement of Higher Level Personnel (CAPES) for scholarship support.