
Abstract
The facial nerve is the most frequently damaged nerve in head and neck traumata. Repair of interrupted nerves is generally reinforced by fine microsurgical techniques; nevertheless, regaining all functions is the exception rather than the rule. The so-called “postparalytic syndrome,” which includes synkinesia and altered blink reflexes, follows nerve injury. The purpose of this study was to examine if nerve-gap repair using an autologous vein filled with skeletal muscle would improve axonal regeneration, reduce neuromuscular junction polyinnervation, and improve the recovery of whisking in rats with transected and sutured right buccal branches of the facial nerve. Vibrissal motor performance was studied with the use of a video motion analysis. Immunofluorescence was used to visualize and analyze target muscle reinnervation. The results taken together indicate a positive effect of muscle-vein-combined conduit (MVCC) on the improvement of the whisking function after reparation of the facial nerve in rats. The findings support the recent suggestion that a venal graft with implantation of a trophic source, such as autologous denervated skeletal muscle, may promote the monoinnervation degree and ameliorate coordinated function of the corresponding muscles.
Impact statement
This is the first attempt for facial nerve repair with muscle-vein-combined conduit. The findings indicate that a venal graft with implantation of a trophic source, such as autologous denervated skeletal muscle, may promote the monoinnervation degree and improve coordinated activity of the corresponding muscles.
Introduction
Facial nerve is the nerve damaged more often after injuries of the head or neck. Facial nerve lesions may occur during car accidents causing fractures of the temporal bone or face traumata. Most facial nerve injuries, however, occur after surgical operations and constitute a complication of parotidectomy, vestibular schwannoma surgery, or petrous bone surgery. After a peripheral nerve is damaged, there is consistently a local effort of the traumatized axons to regenerate, 1 a process that involves a wide spectrum of reactions. 2
Repair of interrupted nerves is usually reinforced by fine microsurgical techniques. Nevertheless, functional recovery is rarely fully succeeded and regaining of facial expressions and of function of all 42 facial muscles remains poor.3–5 The so-called “postparalytic syndrome,” which includes synkinesia and modified blink reflexes,6–8 follows nerve injury. The so-called “misdirected” reinnervation,9,10 the transmission of abnormally intensive stimuli from one axon to another, 11 and the alterations in synaptic input of the facial motoneurons12–15 are the main parameters responsible for the syndrome.
After the nerve is cut, both ends are pulled back and create a gap that is often surgically bridged by nerve grafts. 16 Blood vessels and striated muscles are the conduits that received the highest interest from study groups. A milestone for their use was the demonstration in the 1980s that vein grafts carrying injured nerves resulted in similar effects to autologous nerve grafting.17–21
Striated muscle autografts used to restore nerve damage were documented a long time ago, 22 but were investigated more thoroughly in the beginning of 1980s.23–25 The thought to employ muscular tissue for axonal regrowth was based on the resemblance between the basal lamina (BL) around muscles cells and the BL outside the endoneurial tubes. 23 Although the effectiveness of both autologous vein and muscle conduits has been separately demonstrated both in animal models and human patients,26–33 their positive outcome is usually related only to restoration of small nerve gaps. 34 To enable neural entubulation over longer distances, a conduit made out of venous pieces filled with striated muscular tissue has been previously described.35–37 Another rationale for this approach was that vein collapse can be impeded by the muscle content, while axon dispersion can be hindered by the vein wall. 37 Interestingly, the use of fresh muscle is especially effective when combined with vein conduits, whereas its effectiveness is limited when used with synthetic conduits.38,39
In our previous studies, we attempted to ameliorate axonal navigation using surgical reconstruction with an isogeneic aortic Y-tube and with the use of an aortic three-way guide. Both techniques reduced collateral branching of the axons at the lesion site; nevertheless, polyinnervation at the neuromuscular junction (NMJ) remained high and promoted poor vibrissal motor function (whisking).15,40,41
The purpose of this study was to investigate whether a nerve-gap repair using an autologous vein filled with skeletal muscle would improve axonal regeneration, reduce neuromuscular junction (NMJ-) polyinnervation, and improve the recovery of whisking. Although similar facial nerve reconstructions have been tested before by methods such as fascicular tubulization, 42 synthetic conduits, 43 or venous ensheathment, 44 this is the first attempt for facial nerve repair with muscle-vein-combined conduit (MVCC).
Materials and Methods
Animals and experimental design
A total of 40 adult, female Wistar rats (175–200 g; strain HsdCpb:WU; Harlan Winkelmann, Borchen, Germany) were used. We used only female rats because testosterone has been shown to beneficially affect peripheral nerve regeneration. 45 In addition to that, doing this since 1990 provides us the possibility to compare data among different experimental sets. All rats were provided with tap water and were fed with common food (Sniff, Soest, Germany) ad libitum. A 12-h light cycle with lights on, followed a 12-h dark cycle, with lights off. The procedures were all approved by the animal care committee (Approval number 2133/2.27.201) and were performed according to the German Law for Animal Protection. The recommendations and regulations of the National Institute of Health about animal care were thoroughly followed.
The rats were separated into 4 groups, each of 10 rats. Group 1 consisted of intact (not surgically treated) rats. In animals from group 2, the right buccal branch of the facial nerve (BBFN) was transected and sutured (buccal-buccal anastomosis, BBA). In group 3, both ends of the transected BBFN were inserted into a fragment of an empty autologous external jugular vein, measuring 1 cm. In group 4, the vein conduit contained additionally a small piece of muscle, more specifically, the autologous pectoralis major, thus creating an MVCC.
All surgical processes were conducted with the help of a microscope, after the animals were intraperitoneally injected with Ketamin/Xylazin (100 mg Ketanest and 5 mg Rompun per kg body weight) as described in previous work.46,47
A postoperative interval of 4 weeks was chosen to evaluate vibrissal function of all animals. This was the time after surgery, where the behavioral recordings began to demonstrate a limited restitution of the rhythmical whisking in some animals. Thereafter, we documented the type of motor end plate reinnervation in the levator labii superioris (LLS) muscle, which is the biggest vibrissal muscle.
Surgery for BBA in the rats of group 2
All operations were performed by a trained microsurgeon (Prof. Dr. Nektarios Sinis), under an operating microscope.
In group 2, BBFN was uncovered, cut, and directly stitched with two sutures, type 11-0, epineurally and atraumatically (Ethicon, Braunschweig, Germany). In groups 2–4, the study emphasized on the precision reinnervation after transection by the BBFN; therefore, the aim was to remove possible extra innervation by the marginal mandibular branch. 48 This is the reason why the latter was transected, followed by proximal ligature, to prevent its regeneration when performing BBA (Fig. 1).
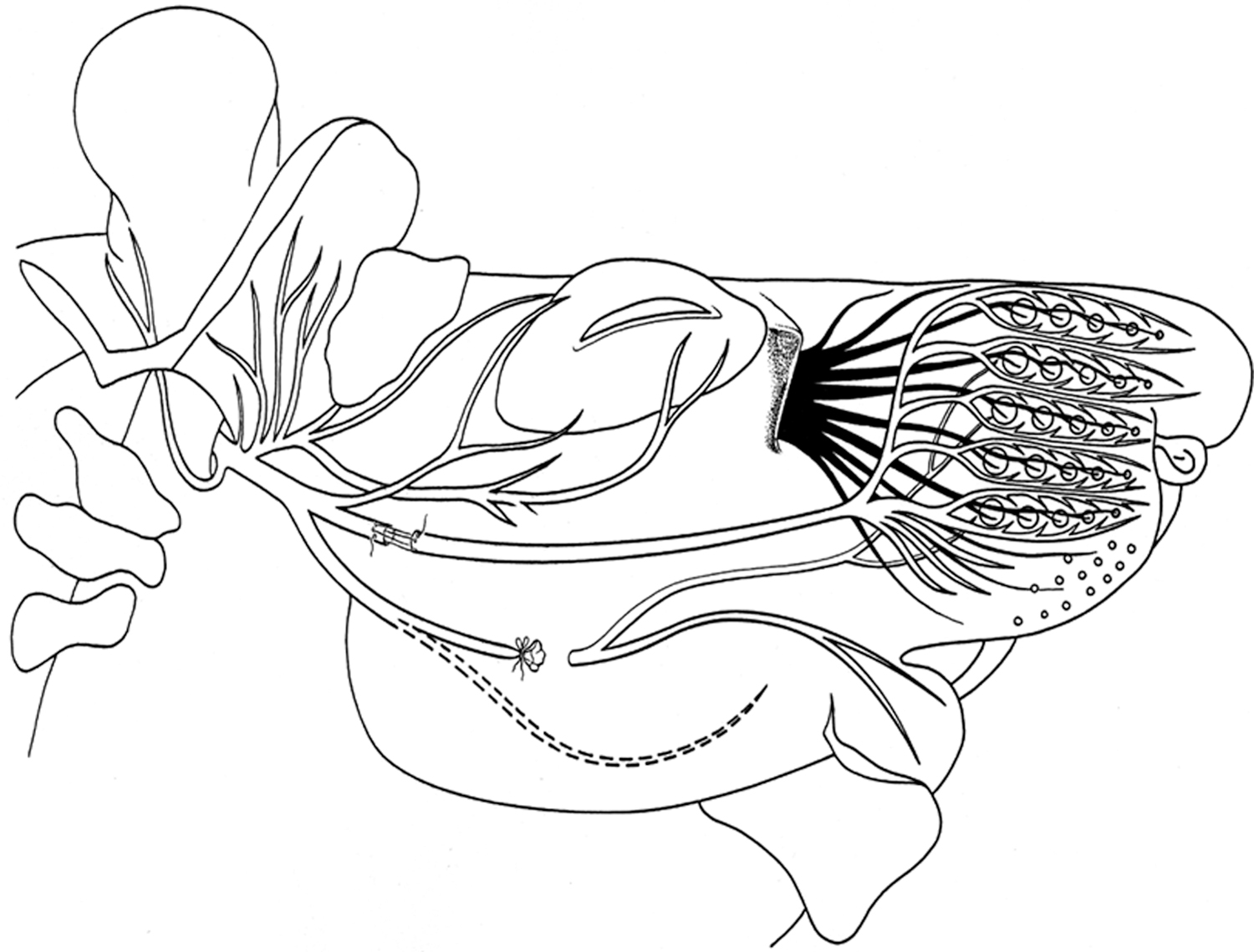
Schematic drawing illustrating the sites of transection and suture in the buccal branch (BBA) and of the transection and ligature of the marginal mandibular branch of the facial nerve. The cervical branch of the facial nerve is indicated by a dotted line. BBA, buccal-buccal anastomosis. Adapted from Dörfl 54 and Semba and Egger. 48
Surgery: entubulation of the transected BBFN
The right BBFN was transected and the two stumps, proximal and distal, were placed into an empty conduit that had been obtained from an autologous external jugular vein (Fig. 2). To avoid tension, the two nerve ends were attached to the vein wall with two epineural sutures, keeping a distance of about 5–8 mm. Previous studies demonstrate that cut axons placed in artificial nerve conduits in rats are capable to bridge gaps that measure up to 10 mm. 49 Phosphate-buffered saline 0.1 M at pH 7.4 filled the area between the two nerve stumps in group 3, while a piece of the ipsilateral m. pectoralis major was added in group 4 (Fig. 3).

Schematic drawing of the infratemporal portion of the rat facial nerve indicating the entubulation site of BBFN. BBFN, buccal branch of the facial nerve. Adopted from Angelov et al. 112
Schematic drawing illustrating the external jugular vein of a rat
Analysis of motor performance
Analysis of the movement of vibrissae was performed with a Video, a procedure that has been conducted many times in previous experimental studies and is very familiar to our research group.41,50–53
Under light anesthesia, on both sides of the face, all vibrissal hairs were cut off, apart from two big ones in row C. We employed a digital camera (Panasonic NV DX-110 EG) and animals were recorded for 3–5 min during active exploration.
We reviewed captured video sequences and 1.5 s sequence fragments from every animal were chosen for examination of whisking biometrics. Selection criteria included the stable position of the animal's head, where whisking frequency and vibrissae protraction degree were clearly documented. The tip of the rat's nose and the inner angles of both eyes were defined as reference points. We evaluated the following: (1) protraction, (2) the whisking frequency, (3) the amplitude, (4) the angular velocity during protraction, and (5) the angular acceleration during protraction. The angle between the midsagittal plane and the hair shaft is a rostrally opening angle, showing the forward movement of the vibrissae, thus the protraction. It is measured in degrees. The cycles of protraction and the passive backward movement, which is acknowledged as retraction, correspond to the whisking frequency. The difference between maximal retraction and maximal protraction in degrees constitutes the amplitude. During protraction, the angular velocity and the angular acceleration are measured in degrees per second (Fig. 4).

The developed spatial model allows precise measurement of angles, angular velocity, and angular acceleration on the intact (left) and operated side (right) during protraction
Analysis of target muscle reinnervation
Four weeks after all surgical treatments, the rats were deeply anesthetized and transcardially perfused with 0.9% NaCl in distilled water for 60 s. They were then fixated with 4% formaldehyde in 0.1 M phosphate buffer at pH 7.4 for 20 min. The LLS muscle (Fig. 5a) of the face of both operated and unoperated side was dissected free, using a surgical microscope. Specimens were cryoprotected in 30% sucrose. Longitudinal sections (30 μm thick) were cut on a cryostat and were mounted on SUPERFROST/Plus slides (Carl Roth, Karlsruhe, Germany) to visualize intramuscular axons and motor end plates. For this purpose, every third longitudinal section through the muscle was stained with anti-neuronal class III β-tubulin (No. PRB-435P; Covance, Richmond, CA), diluted at 1:1000, and Alexa Fluor 488-conjugated α-bungarotoxin, diluted at 1:1000 (B-13422; Molecular Probes) (Guntinas-Lichius et al. 2005). Specificity controls were performed.

Sections were examined with a Zeiss Axioskop 50 epifluorescence microscope through the “rhodamine” filter (No. 15 of Carl Zeiss, excitation BP 546/12, beamsplitter FT 580, and emission LP 590) and through the “fluorescein” filter (No. 9 of Carl Zeiss, excitation BP 450–490, beamsplitter FT510, and emission LP 520). We documented the axonal branches with beta-tubulin staining, entering, or in some cases leaving the boundaries of individual end plates, which were demonstrated with acetylcholine receptor staining, alpha-bungarotoxin. Entries of branches from one axon were accounted as single events. The quality of end plate reinnervation was determined by counting the number of these axonal branches. The end plates were characterized as “monoinnervated” (one axon), “polyinnervated” (two or more axons), or denervated (no visible axonal associated with the receptor staining). All end plate countings were realized directly under the microscope (objective × 400) in a blind manner (Fig. 5b, c). We excluded from the count all end plates that appeared to be cut. The frequencies of mono innervated, poly innervated, and noninnervated end plates were expressed in percentage of the total population and group mean values were compared with the two-sided t test for independent groups. Photographic documentation was made on an LSM 510 confocal microscope (Zeiss).
All the counts were made on sections marked with codes, hiding the information about the surgical procedure used on each animal. All values are presented in the tables as mean ± standard deviation (SD) or as percentages of the total number of labeled motoneurons.
Statistical evaluation
A one-way analysis of variance was used, followed by a post hoc Bonferroni-Holm correction, to investigate the labeled neurons between the control and the experimental subgroups. A p-value of <0.05 was considered to indicate statistically significant differences. If significant effects were detected (p < 0.05), comparisons with the control group 2 were performed using the post hoc test of Dunnett at a significance level of 0.05. For analysis, Statistica 6.0 software (StatSoft, Tulsa, OK) was used.
Results
Application of MVCC improved whisking function
The movement of the large caudal whiskers is regulated by two kinds of muscles. One type is responsible for the movement of the whole mystacial pad, while the other for every individual hair follicle. The second type of musculature is absent in the rostrally situated follicles. 54 The musculature responsible for protraction creates a sling surrounding the rostral part of each hair follicle and its function is controlled by branches of the facial nerve. During contraction, each follicle at its root is pulled downwards, thus shifting the hair upwards/forwards. Retraction of the hair on the other hand mainly depends on the elasticity of the deep connective tissue and occurs passively.54,55
In the control group, the whiskers were upright, oriented anteriorly, and during exploration, they moved back and forth with a frequency around 6 Hz. The highest protraction, which corresponds to the rostrally open angle between the vibrissal shaft and the median sagittal plane, was 70°. The amplitude of whisking, which corresponds to the difference between maximal retraction and maximal protraction, measured 50°. These movements were performed at a sagittal angular velocity of about 500°/s and a sagittal angular acceleration of ∼20.000°/s 2 .
The biometric analysis of vibrissal movements on operated rats demonstrated that MVCC ameliorated the amplitude of whisking to 22.67° ± 4.93°, which was higher than 15.67° ± 2.25° after BBA and higher than 15.21° ± 4.82° after bridging with an empty vein (mean ± SD, n = 10 rats per group). In all operated rats, all the corresponding values remained lower than the values measured in the intact controls (60–70°) (Fig. 6).
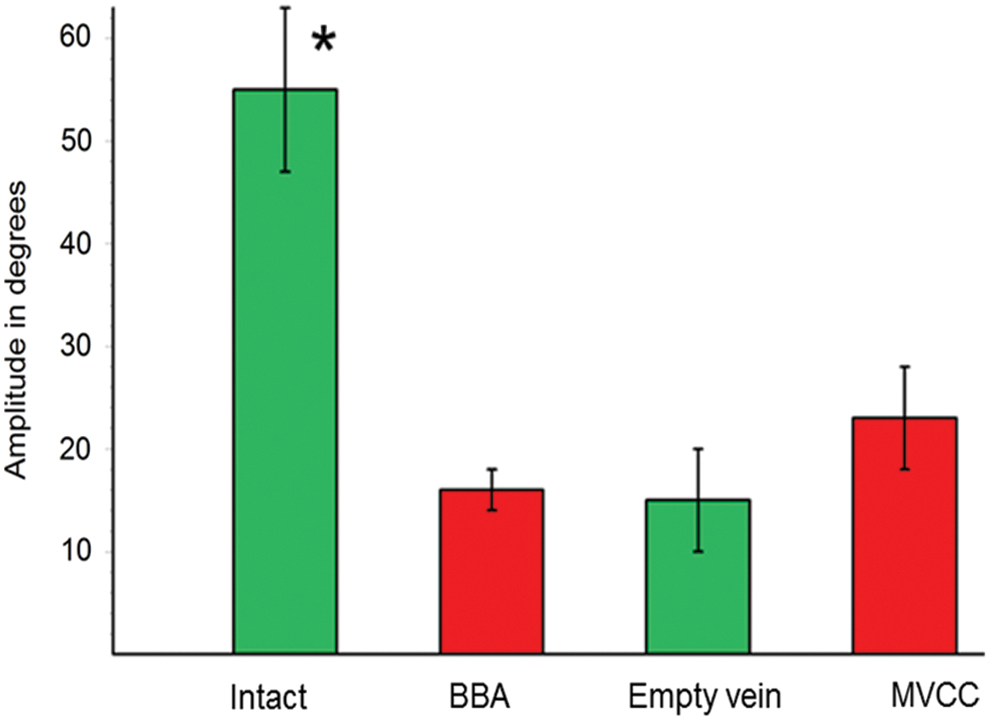
Graphic representation of the results from our biometric analysis of vibrissae movements. Application of MVCC promoted a clear trend for increase in the amplitude (22.67° ± 4.93°), which was larger than that after BBA (15.67° ± 2.25°) and that after bridging with an empty vein (15.21° ± 4.82°; mean ± SD, n = 10 rats per group). Anyway all these values remained significantly lower than those measured in the intact controls (*). MVCC, muscle-vein-combined conduit; SD, standard deviation. Color images are available online.
MVCC improved the quality of target reinnervation
The quality of target muscle reinnervation was evaluated in m. LLS, which is an extrinsic vibrissal muscle. It is innervated the same way as the intrinsic vibrissae muscles, by the “six longitudinal branches” of the BBFN (the short common trunk of the fused ramus buccolabialis superior and ramus buccolabialis inferior). 54
Examinations of the longitudinal sections revealed that MVCC promoted a monoinnervation degree of 74.05% ± 3.91%, whereas after BBA and bridging with an empty vein, the corresponding values were 64.51% ± 15.71% and 67.74 % ± 5.92 %, respectively (Fig. 7).
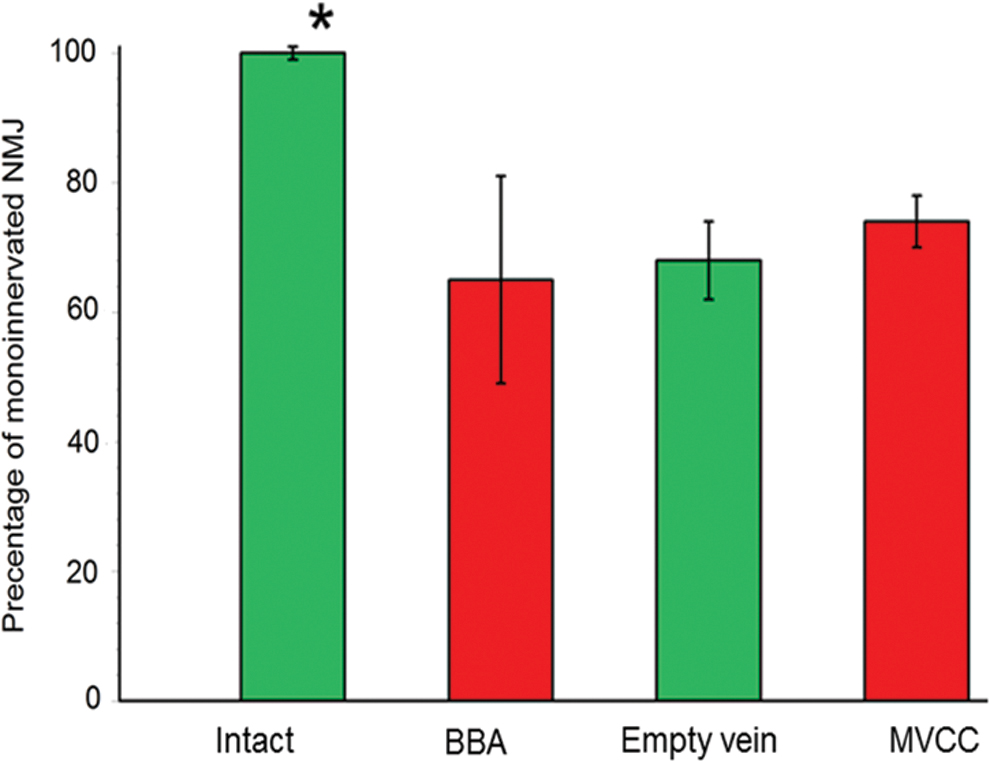
Graphic representation of the results from the analysis of quality of target reinnervation. Application of MVCC promoted an increased monoinnervation degree (74.05% ± 3.91%), whereas after BBA and bridging with an empty vein, these values were 64.51/% ± 15.71% and 67.74 ± 5.92, respectively. All these values remained significantly lower than those determined in the intact controls (*). Color images are available online.
Discussion
The results taken together indicate a positive effect of muscle-vein-combined conduit (MVCC) on the improvement of whisking, following reconstruction of the facial nerve in rats. To our knowledge, this is the first study of such a conduit on the facial nerve. Our findings support the recent suggestion that a venal graft with an implanted trophic source, such as autologous denervated skeletal muscle, may promote the monoinnervation degree and ameliorate the coordinated function of the corresponding muscle fibers.
Misdirected target reinnervation: intramuscular sprouting
It has been established that there are three ways for the post-transectional “misdirected” or “aberrant” reinnervation of muscles to take place. 10 First, the regenerated axons may simply be misrouted along “false” endoneural tubes toward incorrect muscles because of false axon guidance.56–59 The second way is caused by the plethora of branches arising from the cut and misguided axons.60–62 In the course of embryonic development, there is an accurate target-directed pathfinding of single motor axons. 63 During regeneration, however, many branches, rather than one only, arise from the cut axon,8,64–66 and finally, a muscle fiber may be reinnervated by a number of axonal branches coming from separate, individual motoneurons. 67 This is a condition known as “polyneuronal innervation.”68,69 Finally, misdirection can occur within the target, with intramuscular or terminal sprouting of axons. 70 Although thought to be temporary, 71 this abnormal innervation may continue for long time61,72 with harmful consequences on muscle performance.
Sprouting of terminal axons is an impressive occurrence that takes place after neuronal injury and aims to regeneration. Motor units of remaining neurons increase their size aiming to reinnervate as many muscles as possible, thus compensating for reduced functional capacity.73–75 Both collateral branching and terminal axonal sprouting are naturally and automatically realized following segmentation of a peripheral nerve, so it is crucial to understand their influence on the recovery of function. A muscle fiber is regulated by at least two motoneurons, which are often functionally different and may operate asynchronously.73–79 In our previous studies, results show that reduced collateral branching does not promote a decrease of terminal sprouting or an amelioration of the coordinated muscular function, so we suggest that sprouting within the muscle is critical.15,40,41 In this way, presumptions of reports are supported, which demonstrate that enhanced neuromuscular activity ameliorates the quality of target reinnervation by restraining terminal axonal sprouting and by bridge formation of perisynaptic Schwann cell processes.75,80–83
The use of tubes to improve reinnervation/MVCCs
Following nerve transection at the injury site, the majority of neurons react with branching, meaning that each parental axon divides into as many as 25 daughter axons or branches.64,84 Axonal branching is initiated within 3 h after injury85,86 and is considered a pursuit for local navigating cues required for axonal piloting. It is established that if the sectioned nerve ends are placed very close to each other, a large part of the traumatized axons will be entrapped at the lesion area. Overall, guided axonal growth is compromised, mainly because the proceeding Wallerian degeneration is performed slower than axonal elongation. Therefore, the newly developed branches will start growing backwards or will alternatively create a disorganized, disarranged, confused tangled terminal aggregation, called neuroma.87,88 This is the reason why we attached the cut nerve stumps to each other using a tube, enabling an undisturbed axonal regrowth and avoiding a facial-facial anastomosis.
The preferred method used to bridge big peripheral nerve defects is still the autograft, which, however, has serious restrictions, such as limited nerve availability and donor morbidity, presenting mainly with neuroma formation, scarring, and persistent impairment of function. 89 This is why alternatives are investigated and tested. Arterial grafts 90 and especially venal grafts91–95 have been selected based on specific characteristics/abilities: they are able to assist neuronal regrowth at the transected nerve end, they offer a duct where neurotropic and neurotrophic agents that are produced by the traumatized nerve ends can diffuse, and they restrict penetration of fibrous tissue. 94
Every axon along with its myelinating Schwann cell is surrounded by a BL canal, consisting of collagen, laminin, heparan sulfate proteoglycans, and fibronectin. 96 These BL tubes are disrupted by nerve injuries, they then degenerate, and finally they are phagocyted. The notion for utilizing muscle tissue for axonal restoration was based on the resemblance of the muscular BL to the endoneurial tubes. 23 Confocal imaging studies of such conduits demonstrated that the muscle-vein grafts, even from the early days after surgery, were extensively colonized by Schwann cells migrating from cut nerve ends, actively increasing rapidly in number.97–99 In addition to that, 14 days after surgery within the graft, axon regeneration was easily detected. It seems that for sufficient regeneration, it is absolutely necessary for Schwann cells to be very closely situated to the BL. 100 Another rationale for the incorporation of muscle in the vein was that it averts breakdown of the vessel, while the vein itself impedes neuronal spreading. 37 In vitro and in vivo studies have shown that fresh skeletal muscle fibers can dismiss important gliotrophic factors aiming to promote axonal reconstruction and Schwann cell activity. 89
Such combined ducts have been applied in the clinical practice to repair gaps measuring up to 6 cm (mean 2.5), both for sensory and mixed nerves.35,101 In 21 studied cases, both sensory and mixed nerve gaps were bridged, and in 85%, favorable results were reported with a follow-up of at least 14 months. These outcomes appear to surpass those described when different conduits are used, either artificial or biological. 35 Moreover, the muscle-vein conduits are free of cost and can be produced after examining and documenting the gap extent, according to the specific demands, after taking into account the type of nerve and the extend of the damage.
In a following study, Battiston et al. 102 studied 47 treated patients with very restricted indications. A total of 13 patients appeared with sensory nerve damage, 9 with motor nerve, and 25 with mixed injuries. All patients received treatment in emergency, and in all cases, the gaps were too extended and the nerve graft was not sufficient for surgical treatment. There was a satisfying or very satisfying recovery in 52% of the patients with mixed nerve lesions, while 13% exhibited an unsatisfactory recovery. A limited improvement was observed in 35% of the participants in the study with combined nerves injuries; either just the motor or only the sensory nerve fibers exhibited a satisfying improvement. In the cases where nerve lesions were solely motor or sensory, 90% of patients had good recovery results. 102 Overall, favorable results were achieved in a group of 13 patients with crush traumata of sensory and mixed nerves of the upper limb in emergency. In all these patients, a muscle-vein-combined graft was applied for nerve reconstruction. 103
Taking everything into account, Battiston et al. 104 suggest that reconstruction of a traumatized nerve in primary care using a muscle-vein-combined graft is reasonable and promising. Moreover, if the nerve repair fails in this stage, it is still feasible to attempt a secondary reconstruction with a positive outcome. 105
The role of trophic factors in axonal regrowth, future targets
Under normal conditions, the target tissue generates the majority of neurotrophic agents, which are transported retrogradely along the axons to access the motoneurons. Following axotomy, the traumatized axons pick up the trophic factors to maintain the neurons trophically satisfied for as long as the target-derived source is not fully functional.106–110
Neural regeneration constitutes a complicated procedure, which requires the participation of many agents, elements, and signaling cues for it to be effective. The exact extracellular proteins and/or neurotrophic factors that play a key role on the restoration of axons are not fully specified; however, in neurological diseases, the most frequently used and suggested therapies include the administration of neurotrophic factors, since it is known that they enhance motoneuron cell survival during embryonic and early postnatal development. For each type of nerve (motor, sensory, or both) there is a subset of neurotrophic factors recommended. For axonal regeneration, the neurotrophic factors commonly used are NGF, GDNF, brain-derived neurotrophic factor (BDNF), neurotrophin-3,4/5 (NT-3,4/5), and ciliary neurotrophic factor (CNTF). At the time, there are various methods applied for neurotrophic factor transportation, such as matrices, microspheres, hydrogels, or combined muscle conduits. 111
Conclusion
Peripheral nerve regeneration along the distal nerve stump is a fruitless process, unless the surviving axons succeed to reinnervate the original targets. Increasing evidence suggests that investigation should mostly be aiming to find ways to minimize polyinnervation at the NMJ, since recent studies that succeeded to do so, also achieved improvement in function. 41 Our findings support the recent suggestion that a venal graft with implantation of a trophic source, such as autologous denervated skeletal muscle, may promote the monoinnervation degree and ameliorate coordinated function of the corresponding muscles.
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.
Funding Information
No funding was received for this project.