
Abstract
There is a growing need for biomaterial scaffolds that support engineering of soft tissue substitutes featuring structure and mechanical properties similar to those of the native tissue. This work introduces a new biomaterial system that is based on centimeter-long collagen fibers extracted from Sarcophyton soft corals, wrapped around frames to create aligned fiber arrays. The collagen arrays displayed hyperelastic and viscoelastic mechanical properties that resembled those of collagenous-rich tissues. Cytotoxicity tests demonstrated that the collagen arrays were nontoxic to fibroblast cells. In addition, fibroblast cells seeded on the collagen arrays demonstrated spreading and increased growth for up to 40 days, and their orientation followed that of the aligned fibers. The possibility to combine the collagen cellular arrays with poly(ethylene glycol) diacrylate (PEG-DA) hydrogel, to create integrated biocomposites, was also demonstrated. This study showed that coral collagen fibers in combination with a hydrogel can support biological tissue-like growth, with predefined orientation over a long period of time in culture. As such, it is an attractive scaffold for the construction of various engineered tissues to match their native oriented morphology.
Impact statement
This study introduces a unique type of collagen fibers extracted from soft Sarcophyton corals, and demonstrates their biocompatibility and ability to serve as a scaffold that supports biological tissue-like growth with predefined orientation. The coral collagen fibers display hyperelastic and viscoelastic mechanical properties that resembled those of collagenous-rich tissues. These findings demonstrate the great potential of the coral collagen fibers to serve as an attractive scaffold for construction of various collagenous tissues according to their native anisotropic collagen structure and mechanical properties.
Introduction
Collagen is the main component of the extracellular matrix (ECM), and the most abundant protein in the human body, comprising ∼30% of the human proteins, 90% of which are type I collagen. It provides the main load-bearing structure in tissues such as tendons, ligaments, skin, corneas, blood vessels, intervertebral discs, and bones.1,2 Many of these collagenous tissues are characterized by a defined anisotropic orientation of their collagen fibers, which dictates tissue mechanical properties, specifically its hyperelasticity and viscoelasticy.3,4 For example, the outer layer of arteries, the adventitia, consists mainly of type I collagen oriented in two helical groups, and embedded with fibroblast cells aligned in the same directions.4–6 Tendons and ligaments consist of a close pack of collagen fibers oriented parallel to the long axial direction, allowing them to resist large uniaxial tensional forces. 7
Beyond securing mechanical and structural integrity, collagen fibers support the biological activity of cells, including cell morphology, adhesion, migration, and differentiation. 2 Collagen provides amino acid motifs to which cells bind by specific integrin membrane receptors, thus facilitating adhesion and spreading of cells. 8 Moreover, the alignment of collagen fibers can direct cellular organization and encourage the formation of anisotropic tissue-like constructs.9,10 Thus, many studies have utilized collagen-based scaffolds in attempts to engineer tissues mimicking native tissue structures and mechanical properties, which can ultimately improve tissue integration into the host tissue in vivo.11–17
The main types of collagen-based scaffolds currently used in tissue engineering applications include collagen hydrogels, 18 sponges, 19 electrospun fibers, 19 and extrude fibers. 20 To create these collagen scaffolds, a solution of collagen purified from different anatomical parts of animals (e.g., rat tails or bovine skin and tendons 21 ) is typically used. This purified solution is obtained by chemical processes that dissociate the collagen and destroy its native fibrillar structure. As a result, the commonly used collagen-based scaffolds do not preserve the structure and mechanical properties of the native collagen, and tend to be much softer compared with the native form.
Collagen fibers fabricated by extrusion and electrospinning methods20,22 are likely to most closely resemble the structure of native oriented collagen. Yet, they both still suffer from inferior mechanical properties with an elastic modulus of approximately 17–120 MPa for extruded fibers20,22 and approximately 0.3–5 MPa for electrospun fibers,11,23 while native collagenous tissues such as ligaments and tendons, typically exhibit an elastic modulus of a few hundreds of MPa.24–27 Only a handful of studies have attempted to directly extract and use the native collagen fibers as a scaffold to grow cells, including, for example, decellularized scaffolds derived from collagenous tissues.28,29
Recently, we have reported on a unique source for collagen fibers derived from Sarcophyton soft corals, and used these fibers to construct various biocomposites (i.e., collagen-reinforced hydrogels).30–35 The fibers can be manually pulled out of the coral polyp surface, enabling simple and direct extraction of fibers tens of centimeters long.31,33 The biochemical, molecular, and ultrastructural features of the collagen coral fibers have been well described and the amino acid content of these fibers showed close similarity in content to mammalian type I and II collagens. 31 Moreover, the collagen fiber's natural coiled structure and crosslinking are preserved, subsequently retaining their native great hyperelastic mechanical properties. To the best of our knowledge, this is the only system available to date that allows the production of centimeter-long collagen fibers while maintaining their full natural conformation.
Previous studies have achieved a considerable understanding of the mechanical properties of the coral fibers and their ability to form biocomposites that mimic, for example, the collagenous structure of the cornea or intervertebral disc.32–35 However, very little is known about the ability of the coral collagen fibers to support biological cell viability, growth, and adhesion required for tissue engineering applications.30,36 The current study evaluated the biocompatibility of coral-derived collagen fibers and their ability to serve as a scaffold that supports fibroblast growth over time. Cell coverage area, viability and cellular orientation, morphology, and dynamics were quantitatively evaluated. We demonstrate that the coral fibers, alone or in combination with a poly(ethylene glycol) diacrylate (PEG-DA) hydrogel, supported biological tissue-like growth with predefined orientation over a long period of time in culture (∼40 days). Therefore, this source of collagen can serve as an attractive collagen scaffold for the construction of various engineered tissues with native anisotropic morphology.
Materials and Methods
Extraction of coral collagen fibers and generation of aligned collagen arrays
Soft coral Sarcophyton sp. colonies were stored frozen (−20°C) until the day of the experiment. Before the experiment, the coral was defrosted for about 1 h in tap water, at room temperature. Collagen fibers were manually pulled out from the top surface of the coral, creating centimeter-long fibers, with a fiber bundle diameter of approximately 5–10 μm (Supplementary Movie S1). The fibers were wrapped around polylactic acid (PLA) frames (size of the frame was specific for the type of experiment, and will be indicated later for each case). The wrapped fibers and frames were thoroughly washed several times with double-distilled water and any coral remnants were gently removed using tweezers (no damage to the coral was seen by the naked eye).
Collagen fiber sterilization
The collagen arrays were sterilized by soaking them in 70% ethanol for about 1 h, then washing with a few milliliters of distilled water, and exposing to 254 nm ultraviolet light inside a biological hood for about 1 h (30 min each side).
Analysis of collagen fiber fraction
Fiber fraction was measured using bright-field microscopic images of the fiber arrays on a dark background, as previously described.30,32,34,35 Since the specimens were relatively thin (thickness of 0.5–0.7 mm), two-dimensional (2D) image analysis of fiber fractions was performed. The entire fiber array was divided into four window images. The images were processed into binary numerical arrays using a fixed threshold, to separate pixels into white foreground (fibers) and black background. Fiber fraction was then calculated as the average percent of fibers occupying the different analyzed areas. Image processing and calculations were performed using Matlab (Mathworks, Inc., Natick, MA).
Tensile test
Tensile tests (n = 7) were performed using a three-dimensional (3D) printed PLA frame with the following dimensions: total length of 35 mm, gauge length of 25 mm, width of 10 mm, and thickness of 0.7 mm. The two sides of the frame were cut in the center to enable free stretching (Fig. 1A). An Instron-5582 device, with a 100 N load cell and Bluehill 2 operating system, was used in displacement control mode. The bottom of the specimen was fixed, while its upper edge was displaced. Samples were preconditioned to 1.9 mm for two cycles and then stretched at a constant strain rate of 3 mm/min. To prevent slipping, the sample was gripped with a piece of sandpaper. Images of the fibers were taken by a high-resolution charge-coupled device (CCD) camera (Imager pro-SX 5M, and X12; LaVision), placed in front of the fiber array.

Tensile
Digital image correlation (DIC) was employed using a LaVision 2D StrainMaster system, which was used as an optical extensometer to extract the average strain values. Data analysis was performed using DaVis software, which uses the CCD images to calculate the displacement field. Fiber length, width, and thickness were measured at the beginning of each experiment. Throughout the experiment, the fibers were kept moist with Dulbecco's phosphate-buffered saline (DPBS; Biological Industries).
The true (logarithmic) strain,
The normalized nominal stress,
where |F| is the absolute force applied to the specimen obtained from the Instron device,
Calculation of elastic modulus:
where
where
The choice for the strain regions that define the Toe (0–2%) and Linear Modulus (5–8%) was based on the linearity of these regimes, in accordance with Holzapfel, and Fratzl and Weinkamer.37,38
Flexure test
Flexure tests (n = 19) were conducted as previously described.
34
A custom-made PLA frame (40 × 50 mm2, thickness of 0.7 mm) and a dedicated stand were designed to allow gripping of the fiber array in a horizontal position (Fig. 1B). A customized cylinder indenter was designed to press on the horizontal array from above. The Instron program (Blue Hill) was used for displacement and strain measurements. A load cell of 10 N was used and the loading rate was 6 mm/min. Preconditioning was performed by indenting the sample until 1% strain for 10 cycles, followed by 10–15 min of pause to allow stress release. Then, three cycles of loading and unloading until 4 mm deflection were conducted to analyze the hysteresis, according to the following calculation:
Confocal laser scanning microscopy
Images were taken using a confocal laser scanning microscope (LSM 880; Zeiss). The collagen fibers were observed in reflection mode, with a 633 nm excitation laser. Actin green fluorescent protein (GFP) 3T3 cells were imaged using an argon laser (wavelength 488 nm). The objectives used were: × 10 0.3 NA Apochromat air, × 20 0.8 NA Apochromat air, × 40 1.1 NA C-Apochromat water immersion objective lens (Carl Zeiss).
Scanning electron microscopy
The microstructure of the collagen fibers and the seeded cells was examined by scanning electron microscopy (SEM; JCM-6000; JEOL). The arrays were fixed in a 4% v/v paraformaldehyde solution for 20–30 min and washed three times in PBS for 5 min, while being agitated. Samples were then treated with 0.3% Triton X-100, for 10 min, at room temperature, washed three times with PBS and stored in PBS at 4°C. SEM preparation included dehydration by successive ethanol treatment and critical point drying. The samples were then mounted on aluminum stubs and sputter-coated (SC7620; Quorum) with gold.
Cell culture
NIH 3T3 fibroblasts (passages 36–42) stably expressing GFP-actin, were cultured in high-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% nonessential amino acids, penicillin–streptomycin,
Cell seeding on the coral collagen arrays
The sterilized collagen arrays were wrapped around PLA square frames with an area of 49 mm2, and thickness of 0.5 mm. The fiber-frame was placed in the center of a silicone mold (High Temperature Silicone Rubber, that was slightly larger than the frame dimensions; McMaster-Carr) adhered to the surface of a coverslip/culture dish. Then, 4 × 105 NIH 3T3 GFP-actin fibroblasts in 0.5 mL medium were seeded directly on the collagen array, followed by incubation in 37°C and 5% CO2, for 24 h. The cellular constructs were then transferred to a new 35-mm dish (MatTek, Ashland, MA) and 2 mL medium was added for further culturing in the incubator. The medium was replaced every 2–3 days.
Poly(ethylene glycol) diacrylate hydrogel
In some of the cellular experiments, PEG-DA hydrogels were added to the cellular collagen arrays. PEG-DA powder was synthesized and generously supplied by Prof. Dror Seliktar (Biomedical Engineering, Technion-Israel Institute of Technology). Briefly, the cellular collagen arrays were transferred to a new Petri dish and 55 μL 2% or 4% w/v PEG-DA hydrogel solution was gently added to the fibers and polymerized under an ultraviolet (UV) light (VL-6.L 6w- 365 nm power: 12 W; Vilber Lourmat) for 3 min, similar to the procedure reported in Almany and Seliktar. 39 After polymerization, the arrays were transferred back to the dish and incubated in 2 mL growth medium at 37°C and 5% CO2.
Cytotoxicity test
Cytotoxicity test was conducted using the indirect extract method, as described in the ISO 10993 standard parts 5 and 12 for the Biological Evaluation of Medical Devices.40,41 In short, the collagen fiber extract medium was prepared by immersing 6 cm2/mL sterilized coral collagen fibers in culture medium, and shaking for 24 h in the incubator (37°C, 5% CO2). NIH 3T3 cells (5 × 103–6 × 103 per well) were cultured in 96-well plates and incubated overnight. After 12 h, the medium was removed and replaced with 0.2 mL collagen fiber extract in three to six replicates. The cells were then incubated at 37°C and 5% CO2 for 24 h, after which, cytotoxicity was evaluated using alamarBlue (AB; Serotec, Kidlington,United Kingdom).
The AB procedure included replacing the original medium with 0.2 mL fresh medium containing 10% (v/v) AB and incubating the cells for 4 h. Subsequently, duplicates of 0.1 mL from each well were transferred into a 96-well plate for spectrophotometric absorbance analysis (Multiskan Go; Thermo Scientific) at 570 and 600 nm. The percentage difference of AB was calculated in comparison to the regular medium control, according to the manufacturer's protocol calculation:
where: O1 = 80586 = molar extinction coefficient (E) of oxidized AB at 570 nm, O2 = 117216 = E of oxidized AB at 600 nm (O1, O2 are known parameters provided by the manufacturer), A1 = absorbance of test wells at 570 nm, A2 = absorbance of test wells at 600 nm, P1 = absorbance of positive growth control well (regular medium) at 570 nm, and P2 = absorbance of positive growth control well (regular medium) at 600 nm.
Viability of cells cultured on the coral collagen arrays
To quantify the viability of cells cultured on the collagen arrays, an AB assay was used and the percentage reduction of AB was calculated according to the manufacturer's protocol:
where R1 = 155677 = E of reduced AB at 570 nm; R2 = 14652 = E of reduced AB at 600 nm (R1, and R2 are known parameters provided by the manufacturer), N1 = absorbance of negative control well (acellular collagen arrays) at 570 nm and N2 = absorbance of negative control well (acellular collagen arrays) at 600 nm.
Briefly, every 2 days the cellular collagen arrays were transferred to new six-well plates and 2 mL fresh medium containing 10% v/v of AB was added. After 8 h of incubation at 37°C and 5% CO2, 100 μL triplicates of the medium from each well were transferred to 96-well plates for spectrophotometric analysis. Subsequently, the medium containing AB in the six-well plates was replaced with 2 mL fresh medium (without AB), and reincubated under the same conditions. The 8-h incubation time was determined in a preliminary test in which cell growth was tested every hour after adding the AB; 8 h of incubation was found to be the optimum time at which the percentage of AB reduction was in the approximate linear phase.
Cell area analysis
The coverage area of cells on the coral collagen arrays was quantified based on the GFP fluorescence of the cells. A large 36-tile image (area of ∼40 mm2) was taken at the center of the cellular array, using confocal microscopy × 10 objective, every 1–2 days during the first week of culture and every 2–4 days during the following weeks. Each of the tiles was transferred to binary images using a fixed threshold, and the percentage of white pixels (representing the cells) of the total size of the tile image was calculated. The average cell coverage of each type of sample was normalized by the initial cell area percentage measured on the first day of the experiment.
Cell orientation analysis
Cell alignment relative to the fiber orientation was quantitatively analyzed by image processing. Fourteen to twenty-five volume stacks (of a size of ∼4.5 mm2 × 50 μm depth) were acquired with the confocal microscopy × 20 objective, every 1–2 days during the first week and every 2–4 days during the following weeks of culture.
Fiber and cell orientations were quantified by processing the fiber and the cell images separately using an OrientationJ plugin (ImageJ; NIH). 42 The output of the image processing is a color survey map of alignment and a histogram of orientation distribution based on the oriented pixels in the image (Fig. 2). The main orientation of the fibers was defined as a range of twice the standard deviation (2σ) around the mean fiber orientation. To quantify the percentage area coverage by cells oriented along the main direction of the fibers, the fraction of cells (pixel-wise) oriented in the fiber range interval of 2σ was calculated relative to the total pixels of the cells in the image.
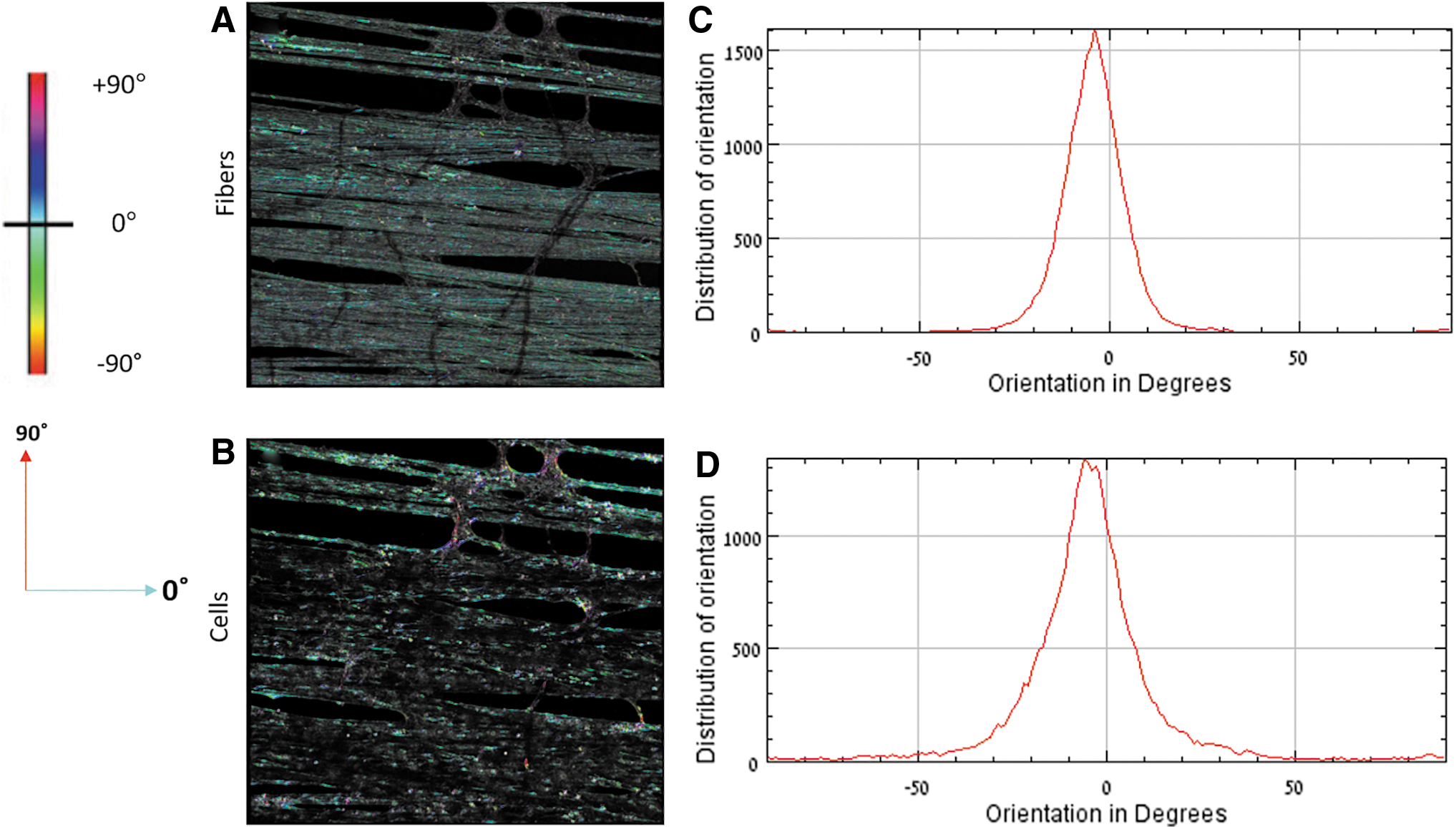
Measurement of cell and fiber orientation.
Statistical analysis
Cytotoxicity and cell area data were tested for normality using the Shapiro–Wilk test. Considering that most populations of the cytotoxicity test were found to be not normally distributed, the one-way analysis of variance on ranks (Kruskal–Wallis) test with Bonferroni post hoc analysis were used to explore statistical differences between the analyzed populations.
For the cell area data, the two-sample t-test and Mann–Whitney U test were used for the normal and not normal distributed populations, respectively. Among the normal-distributed populations, variance equality was tested, and the appropriate two-sample t-test was used (Welch t-test in the cases of unequal variance or two-sample t-test for the rest). A value of p < 0.05 was considered statistically significant. All statistical tests were implemented using the Statistical Package for the Social Sciences (SPSS) software.
Results
Fabrication of aligned coral collagen arrays
Aligned collagen fiber arrays were fabricated by manually pulling the collagen fibers from the coral top surface and wrapping them around thin rectangular frames (Fig. 3 and Supplementary Movie S1). The fibers were wrapped around the frames in a unidirectional orientation, as seen in Figure 3, to create aligned fiber arrays in which individual fiber diameter was about 5–10 μm, and overall fiber density was ∼80% (see Materials and Methods section).

Manual extraction of collagen fibers from the coral.
Mechanical properties of the coral collagen arrays
Mechanical properties of the aligned collagen fiber arrays were characterized by uniaxial tensile and flexure tests (see Materials and Methods section). The average true stress–strain behavior indicated a hyperelastic behavior (Fig. 4A), with an initial elastic modulus of 51.46 ± 24.07 MPa at small strains up to 2.5%, and a higher elastic modulus of 172.28 ± 17.86 MPa at strains 5–10%. Flexure tests indicated the presence of hysteresis, in which the loading phase and the unloading phase did not follow the same curve due to energy dissipation. The average hysteresis was found to be 19% ± 1.2% (Fig. 4B). Overall, the material stiffened with the deflection, similar to the observations of the tensile tests.
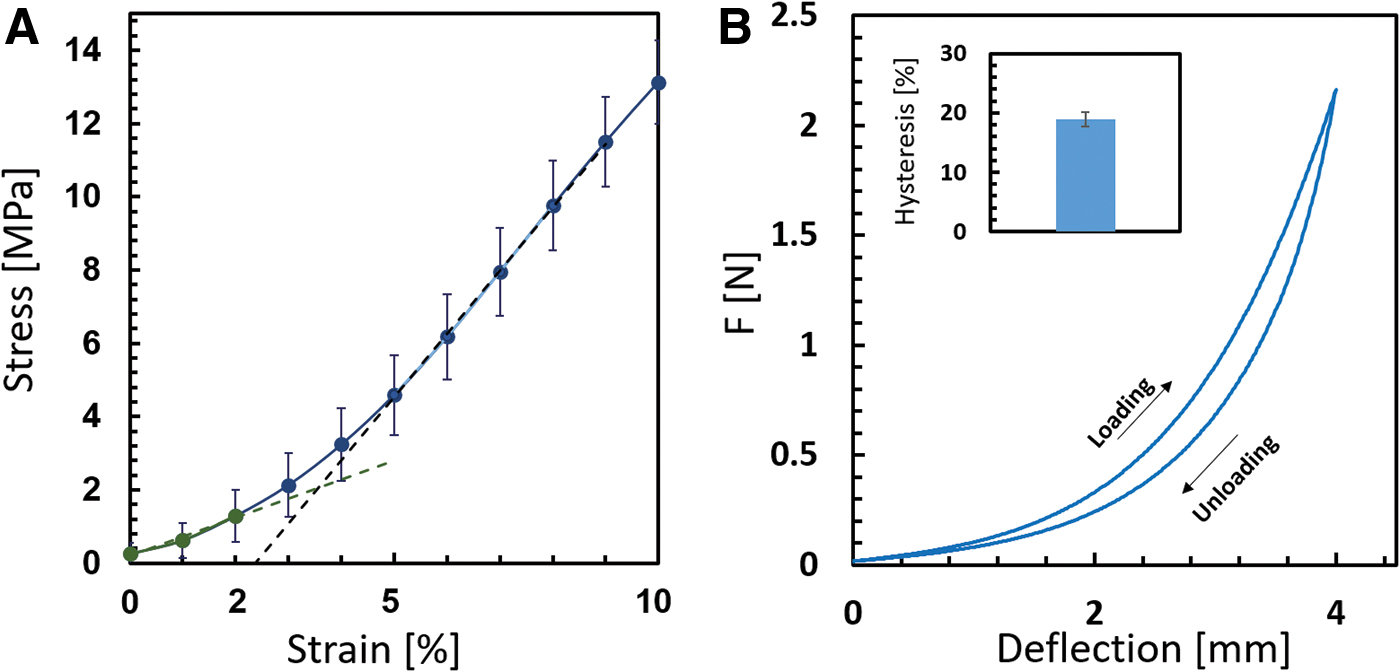
Mechanical characterization of the coral collagen arrays indicates hyperelasticity and viscoelasticity.
Cytotoxicity test of the collagen arrays indicates a nontoxic effect
To evaluate the biocompatibility of the collagen arrays, a cytotoxicity test, based on the ISO standard extract method (10993 standard part 5 and 12)41,42 was performed. Briefly (see Materials and Methods section for more details), an extract was prepared by immersing the collagen fibers in media for 24 h. The 3T3 fibroblast cells were then cultured in the presence of 30% and 100% media extract (Fig. 5), in regular medium (negative control) or in the presence of 70% methanol (positive cytotoxic control). The AB assay was used to evaluate cell viability before adding the extract, and 24 h afterward.
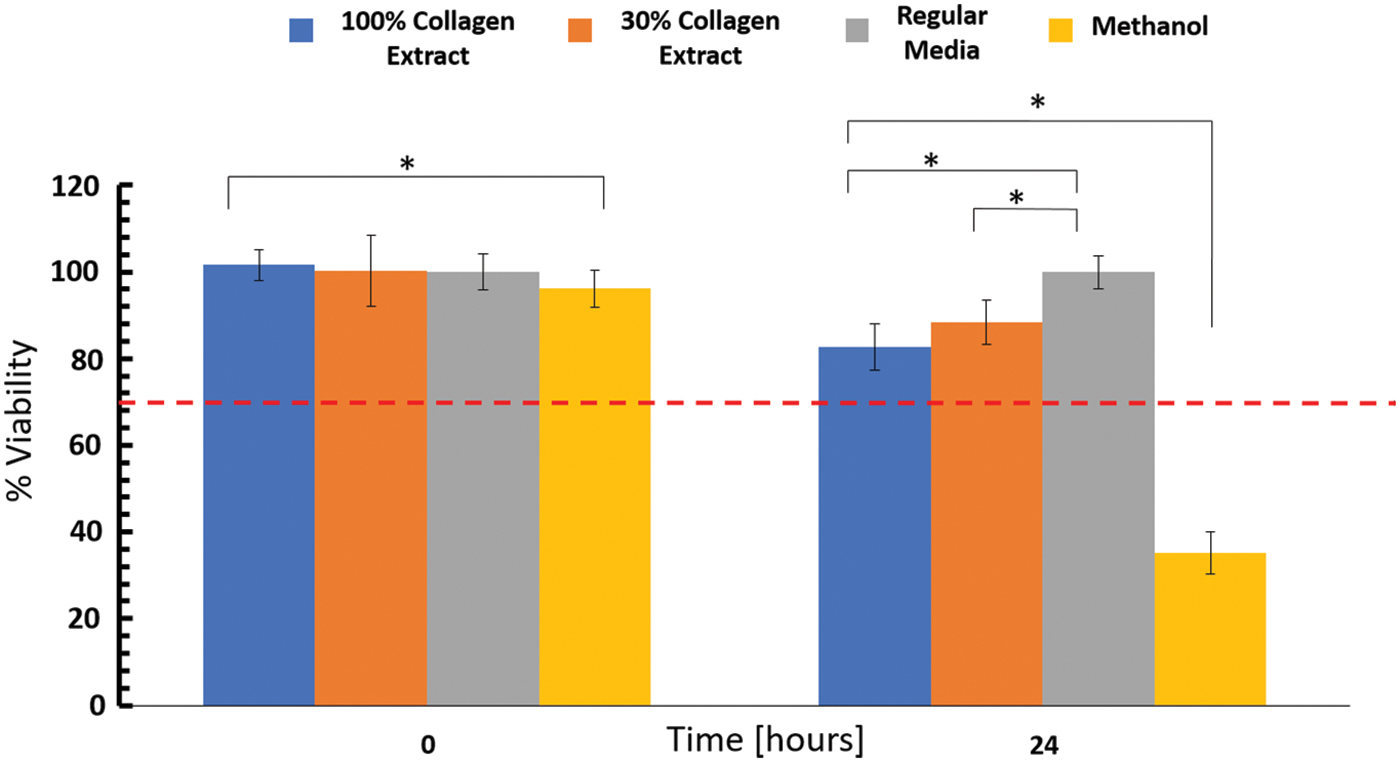
Cytotoxicity test using the extract method demonstrates a nontoxic response. Viability of 3T3 fibroblast cells was examined using the AB assay, after a 24-h exposure to 100% and 30% collagen fiber extract, and compared with the viability of untreated cells (regular medium). Cells incubated in methanol served as the positive cytotoxicity control. Significant differences are marked with an asterisk (p < 0.05). The number of repetitions included at least five wells for each condition. A line at 70% viability is shown to differentiate between viable versus toxic constructs. AB, alamarBlue. Color images are available online.
Before treatment (t = 0), all samples exhibited a similar level of viability, with minor fluctuations attributed to slight variations in the level of cell coverage in the well (Fig. 5). After 24 h of exposure to fiber extracts, the viability of cells exposed to 30% and 100% collagen extract reduced to 90% and 82.7%, respectively, compared with cells in regular media (with p < 0.05 in comparison to regular media). The control groups showed the expected response: viability of cells exposed to methanol reduced to ∼25%, whereas the viability of cells in regular media did not show any reduction.
Taken together, coral collagen fiber extracts were found nontoxic to fibroblast cell culture over a 24-h exposure period, as a value higher than 70% in cell viability is considered to be nontoxic.41,42
Fibroblast cells cultured on the collagen arrays spread out and increased viability over time
To gain more insights into the ability of the coral collagen arrays to support cell growth, actin-GFP fibroblast cells were seeded on sterilized collagen arrays and cultured for up 40 days. Arrays with a fiber density of ∼80% were used, as they were found to better support cell spreading (Supplementary Fig. S1). In part of the experiments, to improve the integrity of the construct, a supporting PEG-DA hydrogel, was included. The PEG-DA hydrogel is a synthetic polymer with an amorphous nano pores structure that was shown to support 3D cell culture when combined with a biological material (e.g., fibrinogen). 43 It was included to coat the collagen fibers and the seeded cells, allowing to form a biocomposite stable construct.
Cells were seeded on collagen fiber arrays, or on collagen fibers combined with PEG-DA hydrogel 2% w/v or 4% w/v. These concentrations are associated with rigidities of ∼400 and 4000 Pa, respectively. 43 To not interfere with initial cell attachment to the collagen fibers, the PEG-DA hydrogel was added to the cellular construct only after observing an increase of about twofold in cell growth parameters (either in cell-GFP area, Fig. 6, or in cell viability, Fig. 7).
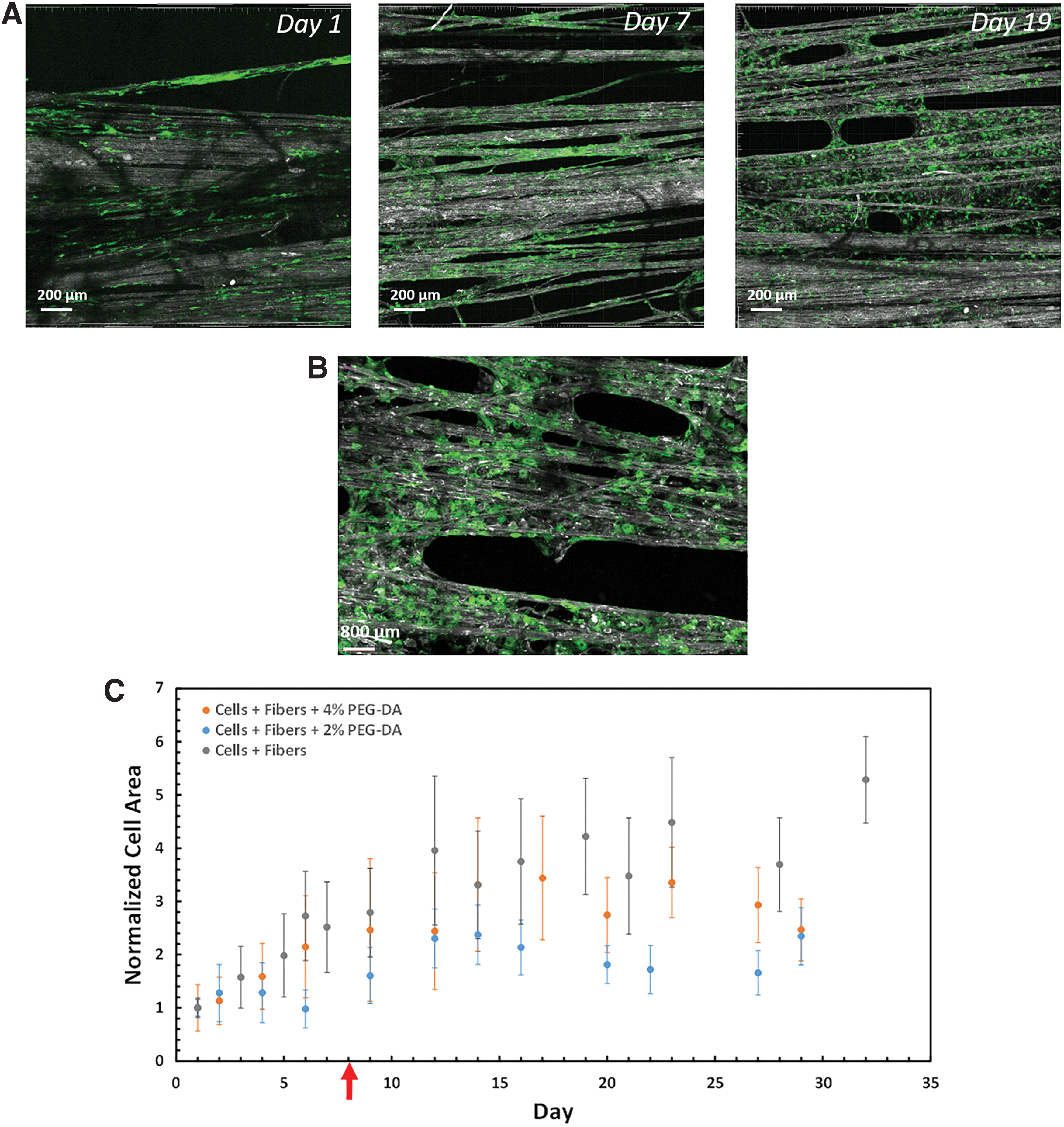
Cells spread out and increased the overall cellular area on the coral collagen arrays over time. Confocal microscopic images of actin-GFP 3T3 fibroblast cells seeded on the collagen arrays.

Viability of cells seeded on the coral collagen arrays increased over time. The 3T3 fibroblasts were seeded on coral collagen arrays, with or without PEG-DA hydrogels (2% or 4%) and monitored over time using the AB assay. The value of the control group of acellular collagen array was substracted from all data points. The PEG-DA hydrogels were added on day 14, as marked by an arrow. Values indicate mean ± SD of three analyzed array samples. PEG-DA, poly(ethylene glycol) diacrylate. Color images are available online.
In all tested arrays, the fibroblast cells attached, spread, and proliferated on the fibers, showing an overall increase in the GFP signal over time (Fig. 6A, B and Supplementary Fig. S2). Quantification of the cellular area, as measured by the total area occupied by the GFP signal, showed an increase during the first 2 weeks of culture, followed by a roughly stable cell coverage during the last 2 weeks (Fig. 6C). The difference between the experimental groups was not found significant at all time points. Therefore, the precise influence of PEG-DA hydrogel on cell area coverage cannot be fully determined. SEM images confirmed the high degree of cell spreading and cell area coverage on the fibers after 30 days in culture (Supplementary Fig. S3).
Quantification of cell viability on the tested arrays using the AB assay, showed that the coral collagen fibers alone, or in combination with the hydrogels, supported cell viability over time, as manifested by an overall increase in the reduction of AB from ∼17% to 40–45% (Fig. 7). The growing time persisted for ∼3 weeks, followed by a roughly stable phase.
By the end of the experiment, the cellular collagen–hydrogel biocomposite was disconnected from the supporting frames and maintained its structural integrity (Supplementary Fig. S4). The possibility to disconnect the construct from the frame strengthens its future potential to be transferred and transplanted in the host tissue in vivo.
Cells aligned along the fibers while presenting various morphologies
The morphology of cells on the unidirectional coral collagen fibers was characterized using confocal microscopy (live assessment through the actin-GFP signal) and SEM (fixed samples on day 30). In general, the cells presented different morphologies, depending on their local fiber environment, as shown in Figure 8. Cells were observed as highly slender and elongated when attached to the aligned bundle of fibers (Fig. 8A). Cells interacting with a single isolated fiber, often wrapped around it, taking on an elongated shape (Fig. 8B). When encountering a dense, relatively flat assembly of fibers that resembled a 2D flat situation, the cells adopted a wider morphology, with apparent stress fibers (Fig. 8C). Cells also filled gaps between fibers, in which cases, they were connected to two separate fiber bundles while assuming a wide morphology with evident actin stress fibers (Fig. 8D).
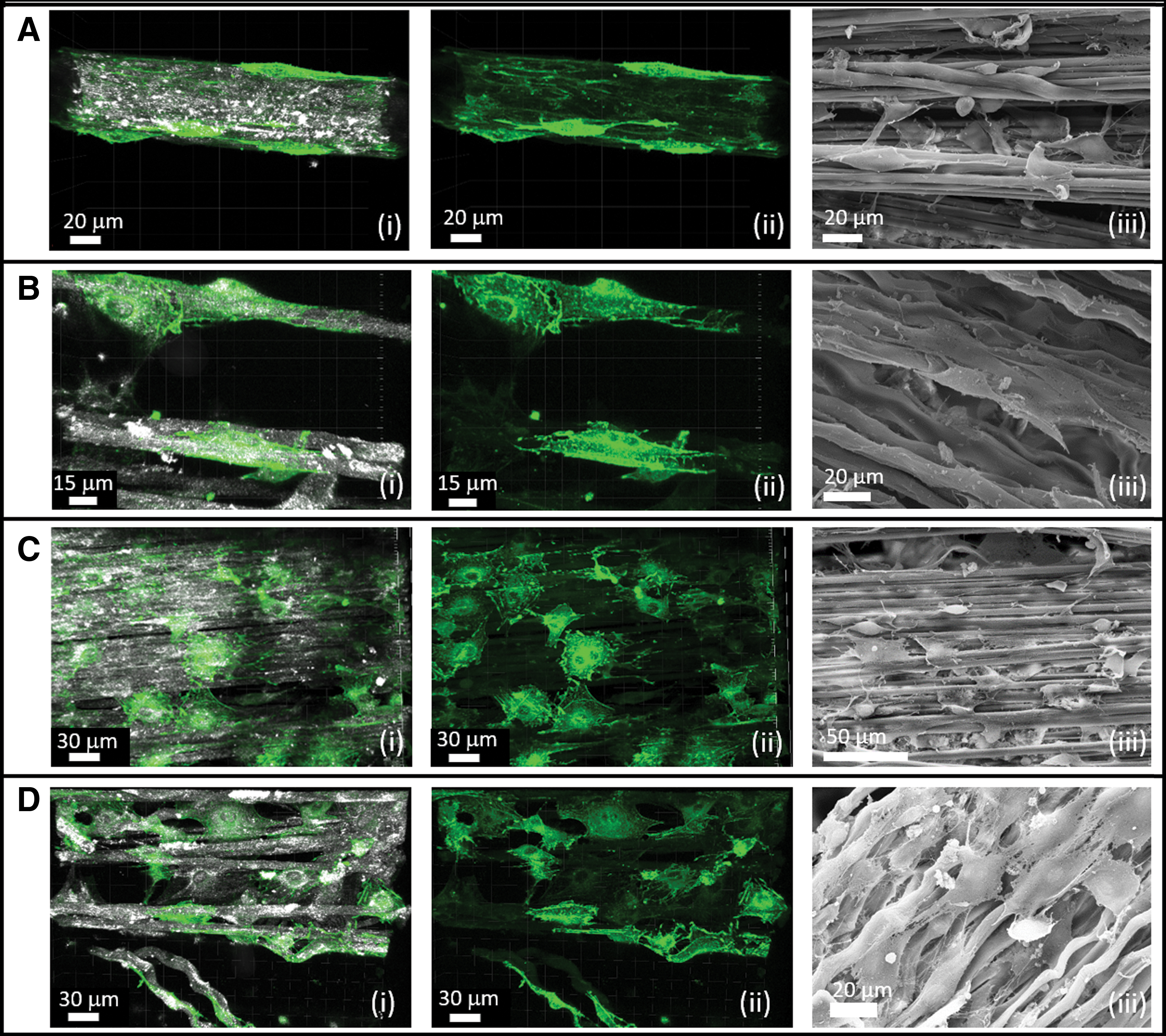
Cells exhibited different morphologies on the aligned collagen arrays. Actin-GFP cells (green) were seeded on the collagen arrays (gray, reflection) and their morphology was monitored on day 32. Several morphologies were detected:
SEM images showed similar cellular morphologies (Fig. 8D.iii). The structure of the collagen fibers and specifically its wavy structure is evident in the SEM images (e.g., Fig. 8D.iii and Supplementary Fig. S3), resembling the wavy structure of native collagen. Moreover, areas of fibers that were occupied with cells were observed to be covered with entanglement of small fibers, presumably these could be new collagen fibers secreted by the cells, whereas fibers without cells were observed to be smoother (Supplementary Fig. S5).
Cell orientation was examined to determine if the aligned fibers directed cell alignment, an important aspect for the construction of anisotropic tissues. Overall, the entire structure of the newly forming tissue had a preferred orientation, as can be seen in the images taken at the beginning (Fig. 9A) and at the end (Fig. 9B) of the experiment. Quantitative image analysis was conducted in which both the orientation of the fibers and the orientation of the cells were independently measured and then correlated with each other (Materials and Methods section). More than 75% of the cellular area was oriented along the main direction of the fibers, on all types of tested samples (with and without the hydrogel) and for the entire duration of the experiment, up to 30 days (Fig. 9C).
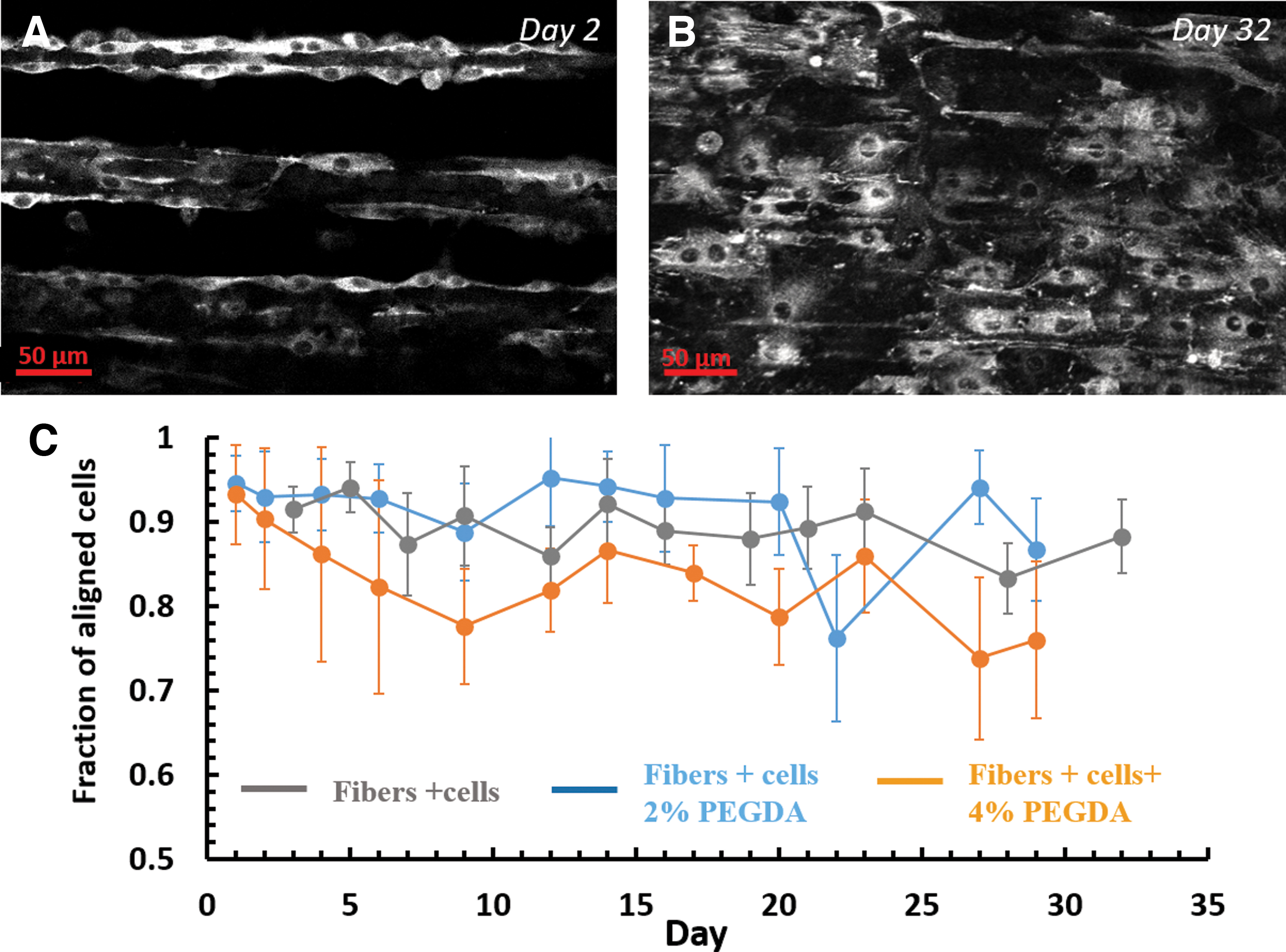
Cells aligned along the main direction of the fibers. Actin-GFP 3T3 fibroblasts were cultured on the collagen arrays and their alignment relative the direction of the fiber arrays, was analyzed.
Cells exhibited dynamic activities on the coral collagen fibers
Time-lapse video confocal microscopy was used to examine the dynamics of cells cultured on the coral collagen fibers. Images were acquired every 15 min for 8–16 h, at different stages from seeding (Supplementary Movies S2, S3, S4). Overall, most cells remained stationary, with their main body intact, while their protrusions dynamically explored their surroundings.
Occasionally, cells migrated along the direction of the fiber (Fig. 10A), in the small gaps between the fibers, or moved between different fibers by extending a long protrusion that reached the new fiber and then propelled the main cell body forward (Fig. 10B and Supplementary Movie S2). Cell migration along the fibers seemed to be more common than migration in other directions. Cell division was also observed (Fig. 10C and Supplementary Movie S3), in which the daughter cells remained aligned and migrated along the fiber after division. Cells were also observed to fill gaps between fibers by first extending a thin protrusion to a nearby fiber, and then spreading in the gap (Fig. 10D and Supplementary Movie S4).
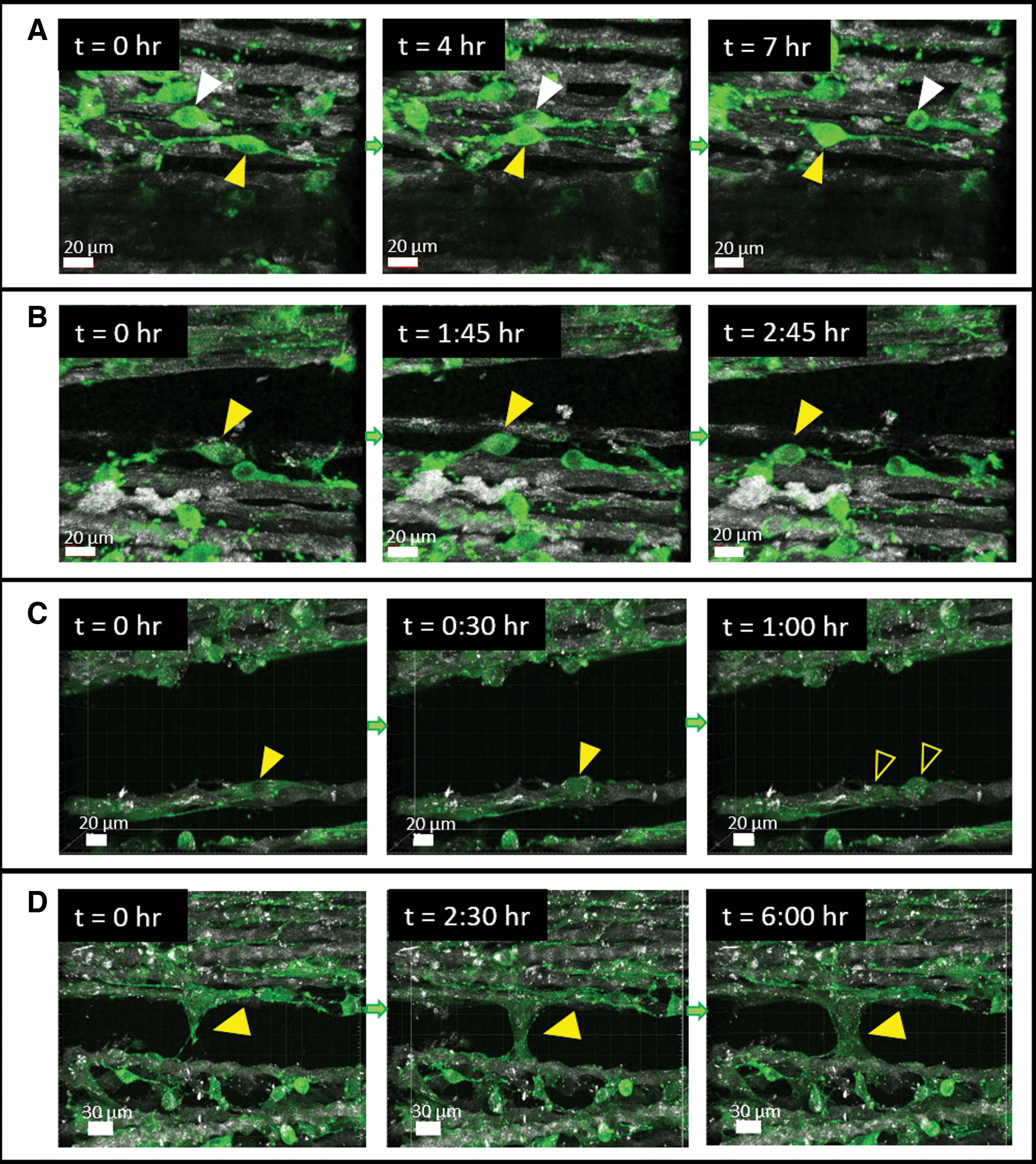
Cell migration and division on the coral collagen arrays. Actin-GFP fibroblast cells were seeded on the coral collagen arrays, and recorded in real-time using the confocal microscopy, over several hours.
Discussion
This study characterized the in vitro biocompatibility of collagen fibers extracted from Sarcophyton soft corals and their ability to serve as a scaffold that supports cell spreading, viability and growth. The extracted collagen fibers were wrapped around dedicated frames to create aligned, unidirectional arrays of collagen fibers, to encourage the formation of an aligned tissue-like construct.
The tensile and flexure tests used to examine the mechanical properties of the collagen arrays have been previously used to characterize the coral collagen fibers but in the context of collagen-reinforced biocomposites.30,32–35 The mechanical properties of the coral collagen arrays, without addition of a hydrogel has yet to be examined. The tensile tests measured an elastic modulus of ∼50 MPa in the toe region (up to ∼2.5% strain) and ∼170 MPa in the linear region (5–10% strain), was within the lower range of native collagenous tissues such as ligaments (approximately 5–450 MPa)25,26 and tendons (approximately 80–1735 MPa).24,25 It should be noted, however, that ranges reported in the literature vary widely, due to the natural variability between different ligaments and tendons (location in the body and the load they bear), intersubject variability, age, and other factors.
Possible ways to improve the strength of the coral collagen arrays can include the following: (1) incorporating a stiffer hydrogel, such as a high concentration of alginate (stiffness of up to ∼1 MPa for 3%w/v Alginate hydrogel 34 ), or a higher concentration of PEG-DA, (2) adding more coral fibers along different orientations (e.g., horizontal) and increasing their volume fractions, 32 and (3) inducing crosslinking of the collagen fibers by UV irradiation44–46 or chemical crosslinkers, such as dehydrothermal treatment, N-hydroxysuccinimide (NHS), or 1-ethyl-3-(3-dimethyl aminopropyl) carbodiimide (EDS)-based crosslinkers.47,48
Importantly, the elastic modulus of the coral collagen fibers is considerably higher than that of other commonly used collagen-based scaffolds, such as collagen hydrogels (0.001–0.3 MPa),49–53 collagen sponges (∼0.021 MPa), 54 electrospun collagen fibers (0.3–5 MPa),11,23 and extruded collagen fibers (17–120 MPa).20,22 The currently available collagen-based scaffolds (e.g., hydrogels, sponges) are generally much softer than the native collagen due to the techniques used for scaffold manufacturing. In contrast, the coral collagen fibers used in this study maintained their original native structure, thereby preserving their mechanical properties.
Moreover, the coral collagen fibers exhibited hysteresis of ∼20%, indicating that the coral fibers featured viscoelastic mechanical behavior, which is in accordance with the viscoelastic behavior reported for various mammalian tendons and ligaments.55–59 Specifically, various types of tendons were found to have a percentage of hysteresis ranging between 3.6% and 45%. 58
For cellular experiments, the coral collagen fibers were sterilized with 70% ethanol and 1 h exposure to UV light (254 nm). This sterilization protocol was selected based on previous studies. 60 We note that the effect of UV irradiation on the collagen fibers was not examined in this study. However, based on previous studies, UV exposure could induce crosslinking of collagen and results in higher stiffness and lower degradability of collagen,44–46,61 without an observed effect on the attachment and proliferation of seeded cells. 46
Coral fibers proved noncytotoxic, with over 80% of fibroblast cells remaining viable after a 24-h exposure to fiber extracts. These findings are in accordance with previous studies that used the extract test to determine the cytotoxicity of other types of biomaterials and concluded a nontoxic response when the viability remained above 70%.62–64 The minor decrease in cell viability can be attributed to the unsatisfactory purification of the coral collagen fibers. Sarcophyton corals, like many other corals, are known to have a mechanism of toxic protection providing chemical defense against their natural predators. 65 Therefore, unsatisfactory cleaning of the fibers can result in the presence of minimal levels of toxic agents, for example, neurotoxin peptides and proteins, or cytolysins that can harm the cell membrane and cause cytolysis.65,66
Another possibility for the decrease in cell viability is that during the extraction process, proteins from the medium adhered to the collagen fibers, and thereby depleted from the medium that was later added to the fibroblast culture. This could have resulted in reduced viability of the cells exposed to extract medium as compared with cells cultured in regular medium. One possible way to test the possibility of depleted proteins is to measure the protein content (e.g., by gel electrophoresis) in the medium before and after 24 h incubation with the fibers.
The coral collagen fibers were shown to support cellular growth for up to 40 days in culture, indicated by the growing level in the cellular area (GFP signal) and AB viability, the spread morphology of the cells, and their dynamic activity. Demonstrating that cells migrated between fibers and that cells divided provide great indicators for the strength of this material for cell culture applications. Previous studies by Benayahu et al.30,36 are the only ones so far that have examined the biological interaction of the coral collagen fibers and seeded cells. However, these studies focused on a system of the coral fibers reinforced with an alginate hydrogel, and provided mainly a qualitative description of spreading and alignment of the seeded cells. In contrast, the present study provides a more extensive and quantitative evaluation of cellular interaction with the coral fibers, with or without the addition of a different hydrogel PEG-DA (2% or 4%).
Specifically, about 2.5-fold increase in cell viability (AB assay) was observed after 40 days in culture (Fig. 7). The growing time persisted for ∼3 weeks, followed by a roughly stable state. The deceleration in the growth rate toward the end of the experiment, may have been due to limited space available in the scaffold for additional cell growth, or more limited diffusion of nutrients or AB in the dense cellular construct. Previous studies that used the AB assay to estimate cell viability in other scaffold types,67–69 also typically observed increased cell viability over the first few weeks in culture, followed by a more steady or even reduced cell viability at later time points.
Quantification of cell alignment demonstrated that a high fraction of cells (>75% cellular area) aligned along the main orientation of the collagen fibers, indicating the great potential of the fibers to support construction of highly aligned tissues, such as tendons, ligaments, blood vessels, skeletal muscle, or nerve tissues. Previous studies have attempted to direct the orientation of other collagen-based scaffolds to direct cells and tissue alignment, for example by directed electrospinning,11,70,71 extruded,20,22 magnetic, 72 or mechanical manipulations. 73
The main advantage of our system is that it is relatively simple and does not involve the sophisticated equipment required for most existing methods. Although the entire cellular construct exhibited an aligned conformation, various cell morphologies were evident, depending on the localized fiber environment, with areas of highly polarized slender cells, and areas occupied by broader cells.
This work demonstrated the ability of the coral collagen fibers to serve as effective and biocompatible tissue engineering scaffold. Future works should focus on the ability of the coral collagen fibers to support the formation of specific engineered tissues, such can tendons, ligaments, skeletal muscles, blood vessels, neurons, or valves. In the case of tissues that are naturally exposed to external mechanical stimulations, such as muscles or tendons, the application of external stretching of the coral collagen arrays inside a bioreactor should be considered.
Additionally, previous studies of cancer cell migration in vivo, for example by Friedl and colleagues,74,75 have described that cancer cells tend to collectively move in open spaces defined by boundaries of collagen fibers or blood vessels. We thus propose for future studies to use the coral collagen arrays as an in vitro model to examine cancer cell migration in gaps between the collagen fibers. Control over the fiber gaps size may be engineered.
Moreover, while this study focused on unidirectional fiber arrays, the fibers can be wrapped in various orientations, to create a complex architecture, as was previously shown by Sharabi et al.32,34,35 Complex fibrillar structures may be advantageous for construction of tissues that consist of various fiber layers each having different orientation, such as the skin or blood vessels. Moreover, evaluation of the degradation rate of the collagen fibers, and the secretion of new ECM by cells, will help to characterize the long-term behavior of the tissue. Also, we assessed the mechanical properties of the construct without seeded cells. Future studies should investigate the properties of the construct with cells, as they may differ from those of acellular constructs. Finally, the biocompatibility of the coral collagen fibers presented in this study should be assessed in future animal models.
Footnotes
Acknowledgment
The authors thank Vered Holdengreber from the Electron Microscopy Unit in Life Science Faculty, Tel-Aviv University, for her assistance with the scanning electron microscope.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Elizabeth and Nicholas Slezak Super Center for Cardiac Research and Medical Engineering under Grant 2018 to the last two authors. The third author acknowledges the support of the Nathan Cummings Chair in Mechanics.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.