
Abstract
Craniofacial reconstruction of critical bone defects typically requires a bone graft. As graft availability may be restricted by disease or comorbidities, tissue engineering approaches are actively sought. The pericranium could provide new bone graft material. During development and repair, bone transitions through a chondrogenic phase. However, with tissue engineering, pluripotent cells can differentiate directly into bone cells. Does ability to recapitulate bone formation in vitro affect osteogenesis and vascularization of pericranium grafts? To answer this, we obtained tissue from nine patients with preplanned craniotomy surgery and studied three-dimensional osteogenesis and angiogenesis of pericranium-derived spheroids. First, we established growth and differentiation conditions on Matrigel. For each spheroid sample, we investigated (i) continuous osteogenic differentiation (COD) and (ii) osteogenic differentiation preceded by chondrogenesis (CD → OD). The effect of vascular endothelial growth factor (VEGF) was compared to VEGF supplemented with fibroblast growth factor, interleukin (IL)-1, IL-6, platelet-derived growth factor, and tumor necrosis factor-α, a growth factor mix (GFM) with possible synergistic effects. In this limited sample, we observed no age- or sex-related differences in cell expansion. Similarly, no statistically significant differences in osteogenic or angiogenic scores between COD or CD → OD spheroids were noted with regular media. In COD, however, VEGF statistically significantly increased angiogenesis compared to control media (p = 0.007). Also, in COD, both VEGF and VEGF + GFM increased osteogenesis (p = 0.047 and p = 0.038, respectively). By contrast, in CD → OD, neither VEGF nor VEGF + GFM yielded statistically significant angiogenesis or osteogenesis scores compared to control media. To understand these results, we characterized spheroid protein expression by nanoliquid chromatography coupled to tandem mass spectrometry. Nine angiogenic proteins were either uniquely expressed or upregulated in COD compared to CD → OD: (i) endothelial markers JUP, PTGIS, PTGS2, and TYMP, (ii) tissue remodeling factors CHI3L1 and MMP14, and (iii) metabolic pathways modulators ANGPTL4, ITGA5, and WNT5A. ANGPTL4, ITGA5, PTGIS, PTGS2, and WNT5A define a conserved angiogenic network and were >2-fold increased in VEGF compared to VEGF + GFM. Finally, we examined bone formation on printable poly-(propylene-fumarate) (PPF) scaffolds for individualized grafting. Under COD + VEGF conditions, PPF scaffolds loaded with pericranium-derived cells displayed hallmarks of spongiform-like bone formation. Thus, the human pericranium may be a potential repository for bone-generating cells with applications in craniofacial bone repair using tissue printing.
Impact statement
The work presented in this study establishes continuous osteogenic differentiation (COD), without a preliminary chondrogenic differentiation phase (CD → OD) as the preferred method to generate vascularized, osteogenic graft tissue using human pericranium-derived cells (PDCs). Using nanoliquid chromatography-tandem mass spectrometry analysis, we identified nine angiogenic proteins that were either uniquely expressed or upregulated in COD compared to CD → OD: (i) endothelial markers JUP, PTGIS, PTGS2, and TYMP, (ii) tissue remodeling factors CHI3L1 and MMP14, and (iii) metabolic pathway modulators ANGPTL4, ITGA5, and WNT5A. PDCs grown under these conditions generated trabecular bone on poly-(propylene-fumarate) scaffolds for individualized grafting.
Introduction
Craniofacial reconstruction surgery involves repair of bone and soft tissue of the head and neck secondary to trauma, removal of malignant or benign tumors or tissue necrosis. A typical limitation is the regenerative capacity of local tissue to close-off the original defect. Critical bone defects cannot heal on their own and require a bone graft. Of the ∼600,000 bone grafts performed each year in the United States over 30,000 are for craniofacial reconstruction. 1 Oftentimes, tissue availability depends on pre-existing conditions. For example, fibular free tissue transfer is the “gold standard” for mandibular reconstruction, 2 but cannot be implemented in many cases. 3
Tissue engineering using effective combinations of scaffolds, stem cells, and cytokines may provide an alternative bone source. Useful scaffolds promote cellular adhesion, minimize immunogenicity, are biodegradable, and support differentiation. 4 Recently, poly-(propylene-fumarate) (PPF) scaffolds with optimal interconnected porous structures and printing capability were developed for personalized grafting.5,6 Stem cells impose a trade-off between pluripotent potential and unintended effects. For example, issues of ethics or teratoma formation limit use of embryonic and induced pluripotent stem cells, respectively. 7 By contrast, adult mesenchymal stem cells (MSCs) are free of these issues, abundant, and easy to harvest, but their osteogenic potential varies from tissue to tissue. To date, bone marrow- and adipose-derived mesenchymal stem cells (ADSCs) have been the preferred source for bone engineering applications.
Periosteum represents another possible source of MSCs for bone applications because it has the ability to heal large defects in both long and flat bones. 8 In the head and neck, the pericranium represents a large sheet of well vascularized tissue that is relatively easy to harvest with low morbidity during preplanned craniotomy.
The periosteum participates in bone fracture healing through timed production of a cartilaginous callus, tissue vascularization, and matrix mineralization followed by apoptotic chondrocyte death and osteoblast repopulation. 9 During healing, the chondrogenic callus precedes bone formation. As new bone adjoining the fractured ends has greater strength than the original tissue. there is the prospect that induction of an initial chondrogenic state may be beneficial for in vitro bone formation.
Growth factors (GFs) are critical components of osteogenesis and angiogenesis. 10 Vascular endothelial growth factor (VEGF) promotes new blood vessel formation 11 while platelet-derived growth factor (PDGF) increases vascularity and maturation of the vascular wall. 12 In endothelial cells, VEGF is sequestered in Weibel-Palade bodies (WPBs) as multimeric complexes with von Willebrand Factor (vWF), an endothelial cell biomarker, which also regulates tissue thrombosis. 13 vWF is a biomarker of early angiogenesis, being exclusively produced by endothelial cells and megakaryocytes. As such, vWF is routinely used for histochemical identification of capillaries in tissue sections.14,15 In addition, proinflammatory factors such as tumor necrosis factor (TNF)-α, interleukin (IL)-1, and IL-6 play critical roles in osteogenesis.15–17
We used spheroids to study pericranium-derived cells (PDCs), bone formation, and angiogenesis as in vitro three-dimensional (3D) models are relevant to in vivo tissue architecture, cellular interconnectivity, and vascular network conditions, 18 whereas two-dimensional (2D) models lack these features.
The goals of our study were twofold. First, we compared two models of PDC osteogenesis: continuous osteogenic differentiation (COD) and osteogenesis that is preceded by a chondrogenic phase (CD → OD). And second, we compared the effect of VEGF alone to that of VEGF supplemented with a growth factor mix (GFM) containing FGF, IL-1, IL-6, PDGF, and TNF-α, with possible synergistic effects.
In the absence of GFs, we find no difference in osteogenesis/angiogenesis scores between the two models. However, in COD, VEGF supplementation increased both osteogenesis and angiogenesis. An overexpressed, highly conserved, five-protein network consisting of ANGPTL4, ITGA5, PTGIS, PTGS2, and WNT5A was identified by nanoliquid chromatography coupled to tandem mass spectrometry (nLC-MS/MS) analysis of spheroids grown in COD + VEGF media. Furthermore, electron micrographs of printable PPF scaffolds populated with PDCs and grown in COD + VEGF media showed hallmarks of spongiform-like bone formation. These results help establish protocols for bone tissue engineering using PDCs with downstream applications in personalized reconstructive craniofacial surgery.
Methods
Isolation of PDCs
Following Institutional Review Board approval for the study (IRB 18-000782), consent was obtained to harvest pericranium tissue during preplanned open craniotomy. With the pericranium exposed, roughly 1–3 cm2 of pericranium was harvested. The tissue was placed in sterile saline, and immediately processed for PDC extraction using established protocols for MSC isolation. 4 Buffers and media were from Life Technologies, unless otherwise stated. Briefly, pericranium tissue was minced with surgical scalpels in 3.0–5.0 mL of freshly prepared 1–1.5 mg/mL collagenase type 1 solution (C0130-1G; Sigma, St. Louis, MO) in advanced minimal essential media (AMEM, 12492013) containing 10% stem cell fetal bovine serum (12662-029) and 1% Glutamax/Pen–Strep (35050-061 and 15240-062, respectively). After 2 h of incubation at 37°C, the fat layer was separated by centrifugation at 500 g for 5 min and discarded. The pellet containing PDCs and other debris was washed, strained through a 70 μm sieve to remove large particulate material, and the cellular fraction was recovered by centrifugation at 500 g for 5 min. Cells were resuspended in AMEM and allowed to attach to polystyrene tissue culture flasks overnight in a 37°C, 5% CO2, and 95% humidity atmosphere. The following day, media containing nonadherent cells were removed and fresh AMEM added. Fresh media were added every 2–3 days and cells passaged 1:2 at 60–80% confluency. Between 4 and 6 passages were needed to collect ∼107 cells at which time aliquots were stored for cryopreservation and Matrigel incubation experiments started. Human adipose MSCs used in control experiments on PPF scaffolds were a gift from Al Dietz.
PDC differentiation on 3D scaffolds
For Matrigel experiments, 24-well plates were coated with 0.5 mL ice-cold Matrigel (Corning; 356234) and allowed to gel 30 min at 37°C. Then, 1 × 106 PDCs in 0.5 mL of stem cell media (SCM, 12492-013) with 10% Matrigel were layered on top. After 2–3 days growth in SCM, spheroids were differentiated using Life Technologies differentiation kits A1007201 (COD) or A1007101 (CD → OD). Total differentiation time was 21 days (Fig. 2A). For CD → OD, spheroids were incubated 7 days with 100% chondrogenic media, 3 days with 50:50 mixture of chondrogenic and osteogenic media, and then 11 days with 100% osteogenic media. To study GF effects, 50 ng/mL final concentration each of VEGF, FGF, IL-1, IL-6, PDGF, and TNF-α was added in SCM media. For PPF scaffold incubations, 24-well plates were precoated with poly-HEMA to minimize cell adhesion to the well 19 and maximize scaffold loading. Printed 1 × 1 × 1 cm, 44% porosity PPF scaffolds were sterilized with 95% ETOH and ultraviolet light, loaded with 2.5 × 106 cells, and gently rocked overnight to enhance pore penetration. The following day, fresh media were added, and tissue culture was maintained for 21 days with medium changes every 3–5 days.
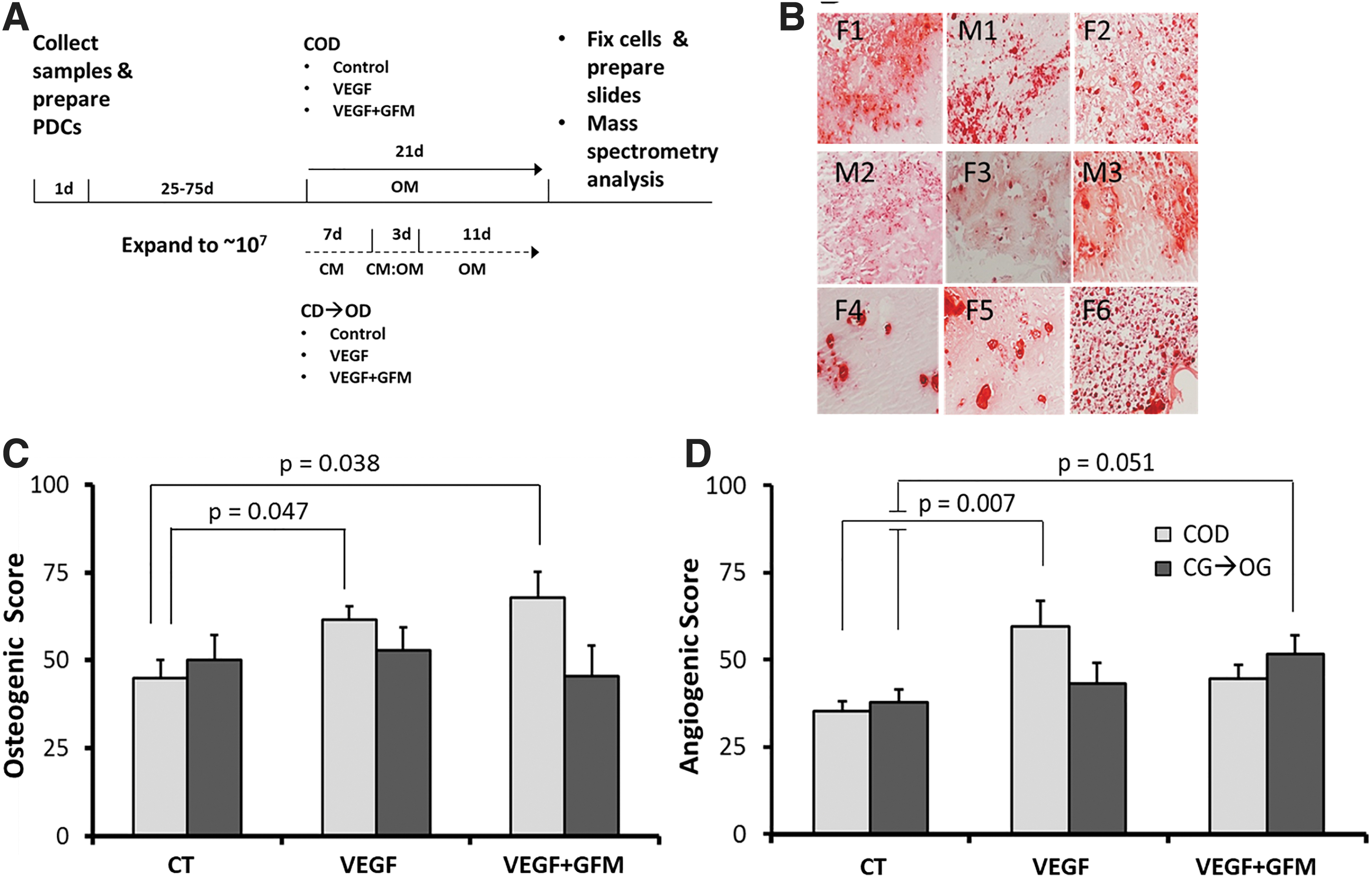
Osteogenesis and angiogenesis of PDC spheroids.
Tissue processing, staining, and image analysis
After 21 days, spheroids were fixed in 4% paraformaldehyde, paraffin embedded, and 5 μm tissue sections were prepared. For staining, slides were deparaffinized in xylene or xylene substitute (2 × 3 min), rehydrated with sequential 100%, 90%, and 70% ETOH (2 × 2 min each), and washed with distilled water. Staining for osteogenic differentiation with Alizarin Red (Sigma; 5533-25G) was performed according to the manufacturer's protocols. vWF staining was performed with Alexa 488-conjugated rabbit anti-human vWF monoclonal antibody EPSISR15 from Abcam. 20 Image capture and analysis were performed with IHC Profiler plugin for ImageJ. 21 In all cases, scores for five slides were averaged. The result was a digital osteogenic and angiogenic score for each sample and incubation condition. To capture differences between samples, final scores were expressed as a percentage of the maximum score (taken as 100%) for each condition.
Scanning electron microscopy imaging
PPF scaffolds with cells fixed in 4% paraformaldehyde were washed in Trump's fixative 22 for at least 1 h at 4°C, washed in phosphate buffer, rinsed in water, and dehydrated through graded ethanol solutions before critical point drying with carbon dioxide. Finally, scaffolds were mounted on an aluminum stub and sputter coated for 60 s with gold-palladium. Imaging was performed with a Hitachi S-4700 cold field emission scanning electron microscope (SEM) at 5 kV accelerating voltage.
nLC-MS/MS analysis
Protein recovery from formalin-fixed paraffin-embedded tissue slides and processing for tandem mass spectrometry analysis were performed as previously described. 23 After Matrigel digestion with MatriSperse, 24 equal cell counts recovered from M2, F3, and F6 were pooled and duplicate analyses were performed. Database searching and criteria for protein inclusion were previously described. 23 The full sample preparation protocol is available online 25 Only proteins with Log2 ion peaks >20 and two or more unique peptides were compared across groups. 26
Statistical analysis
One-way analysis of variance (ANOVA) for treatment differences, regression analysis for age- and sex-related correlations, Fisher's exact test for sex-related effects, and student's t tests for group mean differences were performed in Excel or R. Differences in “Angiogenesis” gene ontology (GO) term enrichment were calculated with z scores. In all cases, statistical significance was assigned at p < 0.05.
Results
Preparation of PDCs
A total of 19 patients undergoing craniotomy and specimen collection were enrolled in the study. Four preparations become contaminated and were discarded. Six specimens were cryopreserved and not further processed. Clinical information on the nine patients who provided experimental data is shown in Table 1. Patients ranged in age from 20 to 81 years, 67% were females and 33% males. A typical starting material is shown in Figure 1A. With one exception (M1), all PDC preparations reached 1 × 107 cells within ∼40 days, requiring 4–6 passages, except F6 that required six passages (Fig. 1B). No age or sex correlations with osteogenic or angiogenic scores were observed (Supplementary Fig. S1 and Supplementary Table S1). These initial data show that human PDCs from both sexes and various ages expand in tissue culture in sufficient numbers to provide starting material for bone grafting.
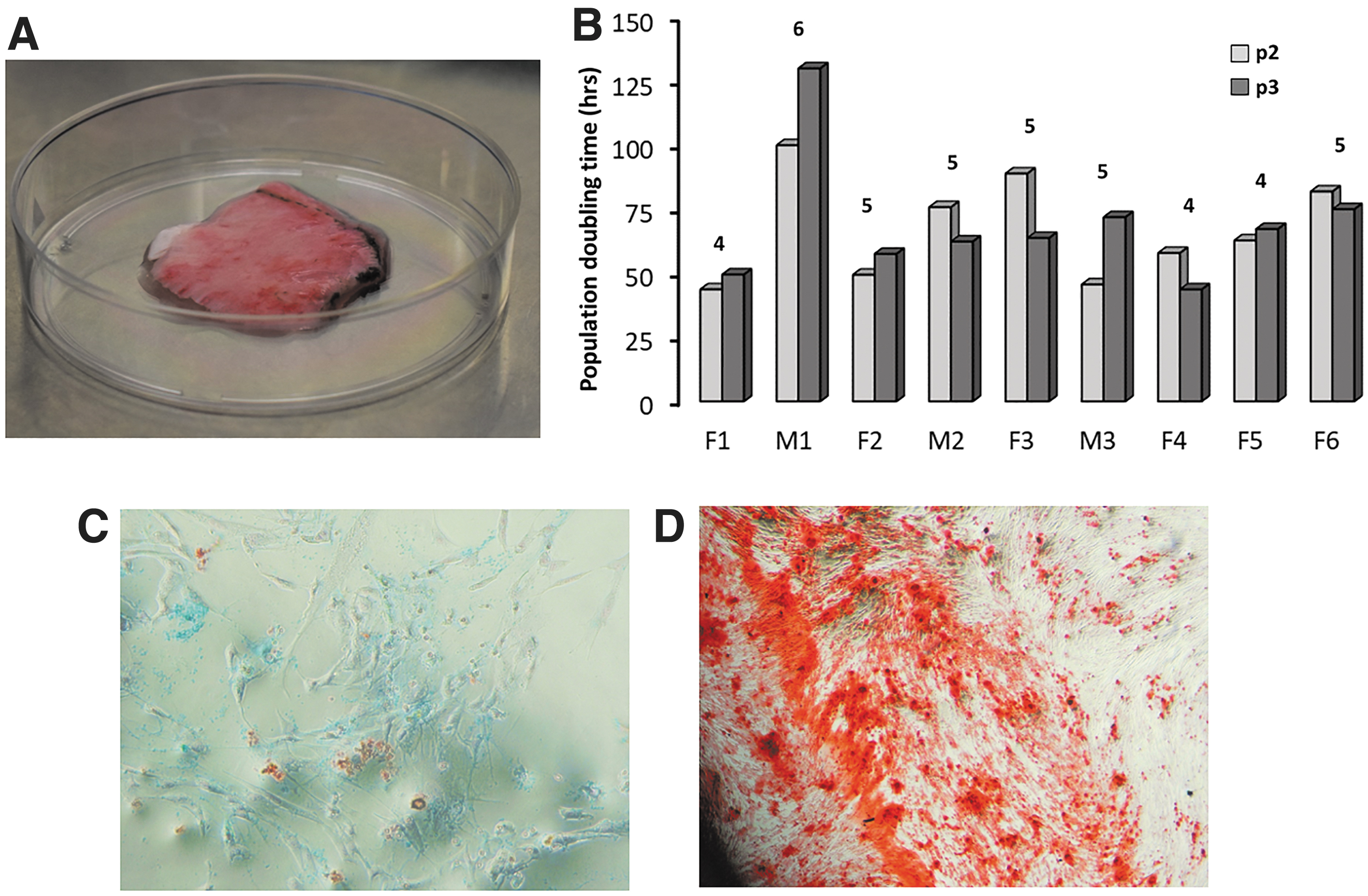
Extraction, expansion, and differentiation of PDCs.
Clinical Data for Tissue Donations That Yielded Experimental Data
CFS, cerebrospinal fluid; CPA, cerebellopontine angle.
COD and CD → OD differentiation of PDCs
Following bone fracture, a cartilaginous callus appears at the reduction site that later progresses to new bone tissue with greater strength than the original bone. 9 Experiments were performed to study the effect of a chondrogenic state preceding osteogenesis compared to uninterrupted osteogenesis on new bone formation and angiogenesis using Matrigel spheroid cell culture. We compared the effect of VEGF 27 alone or supplemented with FGF, IL-1, IL-6, PDGF, and TNF-α. The differentiation scheme is shown in Figure 2A. To demonstrate osteogenic differentiation, duplicate incubations of 1 × 106 PDCs per patient were performed in osteogenic media followed by staining with Alizarin Red. Representative tissue sections are shown in Figure 2B. Similar incubations in chondrogenic media were performed to demonstrate chondrogenic differentiation (not shown). Osteogenic and angiogenic scores were calculated with the IHC Profiler 21 and are shown in Supplementary Tables S3, S4, S5. Osteogenic and angiogenic scores across all six groups were initially compared by one-way ANOVA, resulting in p = 0.016 for angiogenesis and p = 0.217 for osteogenesis (Supplementary Data). Further analysis was performed for COD and CD → OD separately. Under COD conditions, VEGF alone (p = 0.047) and VEGF + GFM (p = 0.038) statistically significantly increased osteogenic scores compared to control media (Fig. 2C). In addition, VEGF alone (p = 0.007), but not VEGF + GFM (p = 0.051), also statistically significantly increased angiogenic scores (Fig. 2D). By contrast, In CD → OD, neither VEGF nor the VEGF + GFM was effective compared to control media (Fig. 2C, D). These results suggest that factor(s) in the GFM may oppose VEGF angiogenesis, but not its osteogenic effect.
Tandem mass spectrometry analysis of protein expression
For nLC-MS/MS analysis, spheroids from three patients were pooled and duplicates analyses were performed. Protein peaks common between duplicate runs were filtered to include only those with associated Gene Names and two or more unique peptides. 28 For the COD group, 662 control (CT), 673 (VEGF), and 725 (VEGF + GFM) proteins were further considered, and for the CD → OD group, 569 (CT), 700 (VEGF), and 600 (VEGF + GFM) were considered. Next, a search for angiogenic proteins was performed using the amigo.geneontology.org search engine with the term “Angiogenesis.” Of the 4267 gene products identified, 598 are expressed in Homo sapiens, and 490 are nonredundant protein terms (Supplementary Data). To determine if any of the treatments produced an enrichment in angiogenic proteins, we performed Venn analysis (Fig. 3A, B) followed by z-score statistical analysis of “Angiogenesis” GO enrichment (Fig. 3C). Slightly different enrichment scores were observed, but none reached statistical significance. Next, we analyzed expression levels of 44 angiogenic proteins representing the most inclusive pool across all samples (Table 2). We identified nine proteins that were either uniquely expressed or upregulated in COD compared to CD → OD: the endothelial markers JUP, PTGIS, PTGS2, and TYMP, the tissue remodeling factors CHI3L1 and MMP14, and the activators/inhibitors of metabolic pathways ANGPTL4, ITGA5, and WNT5A (Table 2). Of these proteins, ANGPTL4, CHI3L1, JUP, and TYMP were >2-fold increased in COD + VEGF compared to COD + VEGF + GFM. Using STITCH network linking analysis, 29 we identified a highly conserved five-protein network (ANGPTL4, ITGA5, PTGIS, PTGS2, and WNT5A) that is overexpressed in the COD + VEGF (Fig. 3D). The protein–protein interaction enrichment p-value was 0.000162, indicating more interactions among themselves than what would be expected for a random set of proteins of similar size, drawn from the genome. Hence, the proteins are at least partially biologically connected as a group.

Angiogenic protein expression in PDC spheroids.
Angiogenic Proteins in Pericranium-Derived Cell Spheroids
Spheroids were prepared as described in section “Methods” and analyzed by nLc-MS/MS. For simplicity, only data for the COD family are shown. Group to group (e.g., COD CT vs. CD → OD CT) and within family (e.g., COD CT vs. COD VEGF) changes in protein levels indicated. Group to group: bold, ≥2-fold upregulated, italic, ≥2-fold downregulated. Within family: underlined, uniquely expressed,
nLc-Ms/MS, nanoliquid chromatography coupled to tandem mass spectrometry; COD, continuous osteogenic differentiation; CD, chondrogenic differentiation; VEGF, vascular endothelial growth factor; CT, control.
SEM analysis of osteogenic differentiation on PPF scaffolds
Biodegradable PPF scaffolds with adjustable porosity were previously described.5,6 These scaffolds are printable, thus amenable for personalized grafting. To investigate osteogenic differentiation, 2.5 × 106 PDCs were applied to 1 × 1 × 1 cm, 44% porosity PPF scaffolds (Fig. 4A) and incubated with osteogenic media (COD CT) or osteogenic media supplemented with VEGF (COD + VEGF). For comparison, 2.5 × 106 ADSCs were also incubated with control osteogenic media. Using SEM, minimal osteogenesis was observed in the ADSC control group at 21 days, consistent with reported osteogenic differentiation potential for these cells (Fig. 4B). By contrast, enhanced osteogenesis with striations reminiscent of trabecular bone formation was observed with PDCs (Fig. 4C). Finally, addition of VEGF further enhanced bone formation, with long, organized bone trabeculae (Fig. 4D). The results demonstrate enhanced bone formation with VEGF supplementation on PPF scaffolds.

Trabecular bone formation on PPF scaffolds.
Discussion
In craniofacial reconstruction surgery, free tissue flaps are used to repair critical defects, but grafting material is often restricted by comorbidities or associated diseases. Therefore, tissue engineering solutions are actively investigated. In this study, we take advantage of the reported potential of the periosteum to fill large bone defects. 8 We used Matrigel to develop methods for 3D osteogenesis and vascularization for translation to PPF scaffolds that can be tailor-made to patient specifications. PDCs extracted from tissue samples from male and female patients 20–81 years old demonstrated ability to expand and differentiate into bone cells irrespective of sex and age. In the absence of GFs, no change in osteogenesis and angiogenesis scores was observed in COD compared to CD → OD. However, addition of VEGF uniquely increased angiogenesis in COD. Both VEGF and VEGF + GFM also increased osteogenesis in COD. By contrast, in CD → OD, addition of GFs was ineffective on these measures. nLC-MS/MS analysis of spheroid tissue sections revealed that factor(s) in GFM oppose the proangiogenic effect of VEGF, resulting in downregulation of a highly conserved network involved in angiogenesis containing ANGPTL4, ITGA5, PTGIS, PTGS2, and WNT5A.
To optimize conditions for cell growth, we first studied PDC spheroids on Matrigel. Matrigel is a basement membrane-derived preparation extracted from Engelbreth–Holm–Swarm mouse sarcoma that provides an extracellular matrix-like 3D environment for cell anchorage and growth. It has been widely used to study spheroid and organoid biology with better inferences for translational work than 2D tissue culture. 30 Non-GF reduced Matrigel (“Matrigel for Organoid Culture”; Corning) was used throughout these experiments motivated by the observation that osteogenic properties are accelerated in MSC spheroids cultured on complete Matrigel. 31 This preparation contains over 1300 different proteins (of which ∼63% are also present in GF reduced Matrigel) when analyzed by mass spectrometry, 30 including transforming growth factor (TGF)-β, epidermal growth factor, insulin-like growth factor 1, FGF, and PDGF, 32 which were shown to promote osteoblast proliferation in vitro. 33 In addition, β-FGF, TGF-β, and PDGF in serum-free medium were necessary for in vitro osteogenic differentiation of human bone marrow-derived MSCs. 34
We hypothesized that prior induction of a chondrogenic state in PDC spheroids may affect 3D osteogenesis and vascularization. This was predicated on observations that, in bone fractures, periosteum initiates a cartilaginous callus with nascent blood vessels. 35 As cartilage-surrounding extracellular cell matrix becomes mineralized, chondrocytes undergo apoptosis, and osteogenesis takes place with the end results that bone with better strength is produced. 9 However, in the absence of GFs, osteogenic and angiogenic scores for COD and CD → OD were similar even though exposure to osteogenic media in the CD → OD group was approximately half of that in the COD group (11 days compared to 21 days). The data do not allow us to distinguish if chondrogenic and osteogenic media exposure induced similar differentiation processes or short exposure to osteogenic media is sufficient to activate them.
vWF is a multidomain protein-binding glycoprotein present in plasma and endothelial cells and an early biomarker of angiogenesis. VWF is essential for WPB formation that regulates angiogenin-2 (Ang-2) levels by controlling storage and synthesis. Ang-2 released from WPB synergizes with VEGFR2 signaling to destabilize blood vessels and promote angiogenesis.13,36 Our data show that in COD spheroids, VEGF uniquely increased vWF levels relative to control media and VEGF + GFM (Fig. 2D). Recent observations that vWF promotes angiogenesis through its heparin-binding domain 37 raise the possibility that proteins that compete for binding may sequester vWF into nonproductive complexes. To further understand protein production by GFs, we performed nLc-MS/MS analysis of spheroids. Among the sets of proteins considered for analysis, there was no statistically significant enrichment in angiogenic proteins, suggesting that differences in angiogenesis scores may be caused by relative protein expression levels rather than a larger pool of angiogenic proteins. We identified five proteins that were more than twofold downregulated in VEGF + GFM compared to VEGF alone: ANGPTL4, ITGA5, PTGIS, PTGS2, and WNT5A. Given these proteins' interaction, as a highly conserved proangiogenic network, the results indicate that a factor or factors in the GFM opposed VEGF on angiogenic protein synthesis. Unfortunately, our design does not allow us to identify factor(s) involved. Furthermore, the effects may be tissue and development stage specific. For example, IL-1α and TNF-α reportedly increased WNT5A in keratinocytes, 38 but in adipocytes, IL-1α upregulation is associated with downregulation of ANGPTL4. 39 Further experiments where the effect of FGF, IL-1, IL-6, PDGF, and TNF-α is separately investigated are required.
Finally, when knowledge gained from growing PDC spheroids on Matrigel was applied to PPF scaffolds, we noted a substantial bone-like growth upon COD + VEGF incubation (Fig. 4D).
Recently, five separate mechanisms of bone formation have been recognized 40 : (i) endochondral ossification, in which MSCs develop separate chondrogenic and osteogenic lineages, (ii) intramembranous ossification, in which osteoblast development occurs in the absence of a chondrocyte template, (iii) chondrocyte to osteogenic precursor, in which immature chondrocytes give rise to osteogenic precursor cells, (iv) chondrocyte de-differentiation into precursor bone cells that proceed to differentiate into bone cells, and (v) direct transdifferentiation of chondrocytes into osteoblasts. Further development of these models complete with identification of biomarkers and pathways involved is greatly anticipated.
In summary, our results show that human PDCs can expand in cell culture irrespective of age and sex. Furthermore, PDCs form spheroids with higher osteogenic and angiogenic scores on Matrigel after supplementation with GFM. GFM supplementation also increased new bone formation on PPF scaffolds. Finally, beginning with a chondrocytic medium to mimic callus formation and then switching to osteogenic media did not augment bone formation or angiogenesis. These observations will catalyze future research into bone substitutes for the repair of critical craniofacial defects.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.