
Abstract
Transforming growth factor beta (TGFβ) signaling is required for in vitro chondrogenesis. In animal models of osteoarthritis (OA), TGFβ receptor alterations are detected in chondrocytes in severe OA cartilage. It is not known whether such changes are dependent on the grade of human OA and if they affect chondrogenesis. Thus, the purpose of this study was to determine if human OA chondrocytes obtained from low-grade or high-grade disease could form cartilage tissue and to assess the role of the co-receptors, endoglin (ENG) and TGFβ receptor 3 (TGFBRIII), in the regulation of this tissue generation in vitro. We hypothesized that the grade of OA disease would not affect the ability of cells to form cartilage tissue and that the TGFβ co-receptor, ENG, would be critical to regulating tissue formation. Chondrocytes isolated from low-grade OA or high-grade OA human articular cartilage (AC) were analyzed directly (P0) or passaged in monolayer to P2. Expression of the primary TGFβ receptor ALK5, and the co-receptors ENG and TGFβRIII, was assessed by image flow cytometry. To assess the ability to form cartilaginous tissue, cells were placed in three-dimensional culture at high density and cultured in chondrogenic media containing TGFβ3. ENG knockdown was used to determine its role in regulating tissue formation. Overall, grade-specific differences in expression of ALK5, ENG, and TGFβRIII in primary or passaged chondrocytes were not detected; however, ENG expression increased significantly after passaging. Despite the presence of ALK5, P0 cells did not form cartilaginous tissue. In contrast, P2 cells derived from low-grade and high-grade OA AC formed hyaline-like cartilaginous tissues of similar quality. Knockdown of ENG in P2 cells inhibited cartilaginous tissue formation compared to controls indicating that the level of ENG protein expression is critical for in vitro chondrogenesis by passaged articular chondrocytes. This study demonstrates that it is not the grade of OA, but the levels of ENG in the presence of ALK5 that influences the ability of human passaged articular chondrocytes to form cartilaginous tissue in vitro in 3D culture. This has implications for cartilage repair therapies.
Impact statement
These findings are important clinically, given the limited availability of osteoarthritis (OA) cartilage tissue. Being able to use cells from all grades of OA will increase our ability to obtain sufficient cells for cartilage repair. In addition, it is possible that endoglin (ENG) levels, in the presence of ALK5 expression, may be suitable to use as biomarkers to identify cells able to produce cartilage.
Introduction
Osteoarthritis (OA) is a chronic disease affecting millions of people globally. 1 The cause is multifactorial and ultimately results in an imbalance in the catabolic and anabolic processes within the articular cartilage (AC) leading to progressive destruction of the tissue. Biological approaches to repairing damaged cartilage tissue are currently being investigated as an alternative to delay or replace total joint replacement for patients with severe OA. Chondrocytes are a good cell source for these therapies as they are easily attainable through arthroscopy. The cells, however, are obtained from tissues affected by OA, and a multitude of studies have demonstrated that OA chondrocytes have an altered phenotype compared to cells from nondiseased tissue, which includes increased expression of cytokine receptors 2 and production of catabolic enzymes.3,4 In addition, studies have demonstrated loss of expression of hyaline cartilage matrix genes by chondrocytes derived from diseased tissues compared to normal, 5 or from cartilage within the same joint with less advanced OA. 6 Studies analyzing RNA expression in chondrocytes isolated from minimal or severe OA cartilage within the same joint7–9 identified a multitude of differentially regulated genes and protein networks, such as those linked to ECM organization, mitotic cell cycle, skeletal development, transcriptional regulation, apoptosis, and Wnt signaling. 9 These observations suggest that even though cells from the same joint share similar systemic and locally generated factors, chondrocytes in sites of advanced OA are phenotypically distinct from those in sites that appear to have little or no OA changes. What remains unclear is whether these differences influence the ability of the cells to form new cartilage and ultimately whether they could be used for cell therapies.
Passaging human chondrocytes in monolayer culture is required to increase cell numbers; however, this results in dedifferentiation. 10 Chondrocyte dedifferentiation is characterized by a change in cell shape (from rounded to elongated) and loss of hyaline cartilage gene and protein expression, thus limiting the use of these cells clinically. In a previous study, we showed that growing passaged cells in three-dimensional (3D) culture in the presence of transforming growth factor beta (TGFβ) re-establishes the articular chondrocyte phenotype resulting in hyaline cartilage matrix macromolecule accumulation (collagen type 2 [Col2] and aggrecan [ACAN]) in vitro, resulting in cells that can form cartilaginous tissue. 11 Others have also demonstrated the importance of TGFB supplementation of the media to facilitate neocartilage formation in vitro, examples of which are bovine chondrocytes that are passaged more than three times 12 and human mesenchymal stem cells. 13 These studies suggest that TGFβ has an important role to play in regulating hyaline cartilage tissue formation in vitro.
Studies in animal models have demonstrated the importance of having sufficient TGFβ receptor expression to maintain the integrity of AC.14–16 Signaling by the TGFβ receptor ALK5, through the phosphorylated SMAD-2/3 complex, is known to promote expression of cartilage matrix genes. 17 In mouse models, ALK5 expression was shown to be diminished in severe OA cartilage 14 and this loss was also thought to play a role in the pathogenesis of human OA. 18 In addition to ALK5, two TGFβ co-receptors have been identified in human chondrocytes, TGFβ receptor 3 (TGFβRIII or betaglycan) and endoglin (ENG or CD105). 19 These have been shown, in other cell types, to enhance TGFβ signaling20–22 and confer chondrogenic potential.23,24 The role these co-receptors play in modulating in vitro chondrogenesis by articular chondrocytes is still undetermined. Furthermore, it is unclear whether human chondrocytes obtained from severe OA cartilage have altered TGFβ receptor and co-receptor profiles compared to cells from tissues with less advanced OA in the same joint, which may influence the ability of the cells to form cartilage in response to TGFβ supplementation. Understanding these phenotypes is critical for developing effective cartilage repair approaches.
Translation of cartilage repair approaches has been limited by the amount of cartilage that can be used as a source of chondrocytes. Furthermore, we do not know if cells obtained from high-grade OA tissue have the same capability as cells from low-grade OA to form cartilage tissue in vitro. Thus, the purpose of this study was to determine if human OA chondrocytes obtained from low-grade or high-grade disease could form cartilage tissue and to assess the role of the TGFβ co-receptors, ENG and TGFBRIII, in the regulation of this tissue generation in vitro. We hypothesized that the grade of OA would not affect the ability of cells obtained from these tissues, following passaging, to form cartilage in vitro and the level of expression of the co-receptor, endoglin, would be critical in the regulation of tissue formation.
Materials and Methods
Isolation of chondrocytes from cartilage
Human AC was obtained from femoral condyles resected during joint arthroplasty for OA following informed patient consent and with Research Ethics Board (Mt. Sinai Hospital) approval. Cartilage was either taken from areas that macroscopically appeared undamaged and termed “Low OA” or from areas that showed signs of severe OA (cartilage thinned or adjacent to erosion) and termed “High OA.” Representative fragments were submitted for histological assessment to confirm the OA grade using the Osteoarthritis Research Society International (OARSI) grading scale. 25 Chondrocytes were isolated from the remainder of the tissue by enzymatic digestion as described previously). 11 Cells from each patient (Low OA or High OA) were cultured separately (cells were not pooled). The cells were either placed in 3D culture directly or in monolayer culture at 2000 cells/cm2 in high-glucose Dulbecco's modified Eagle's media (DMEM) supplemented with 20% fetal bovine serum (FBS). Chondrocytes were passaged twice (P2) to allow for approximately nine population doublings as described previously. 11
Three-dimensional culture
Primary (P0) or passaged (P2) cells (2 × 106) were placed on Col2-coated (c9301; Sigma-Aldrich) membranes (12 mm diameter; EMD Millipore™) and cultured in serum-free chondrogenic media (SFM) as previously described. 11 Ten nanograms per milliliter human recombinant TGFβ3 (R&D Systems) was added after 24 h. Cells were cultured for 3 weeks and the media were changed thrice per week.
Tissue content analysis
Detailed description of the methods can be found in the Supplementary Data. DNA content was quantified by the Hoechst 33258 dye binding assay and fluorometry (excitation λ = 365 nm and emission λ = 458 nm). 26 Hydroxyproline content was quantified following HCl hydrolysis of the papain digest using the chloramine-T/Ehrlich's reagent assay and spectrophotometry (λ = 560 nm). 27 Sulfated glycosaminoglycan (GAG) content was quantified by dimethylmethylene blue dye binding assay and spectrophotometry (λ = 525 nm). 28
Histology
Tissues, native and bioengineered, were fixed in 10% neutral buffered formalin (pH 7.4) overnight, placed in a 30% sucrose/phosphate buffered saline (PBS)−/− solution overnight at 4°C, and then snap-frozen in Tissue-Tek OCT (Sakura Finetek) for cryosectioning. Seven-micrometer sections were either stained with hematoxylin and eosin or toluidine blue. Native tissues were evaluated using the OARSI grading scale. 25 Tissues formed in vitro by cells treated with either ENG or control siRNA were evaluated using the Bern Score. 29
Immunohistochemistry
Details of the methods used can be found in the Supplementary Data. Briefly, tissues were incubated overnight (4°C) with antibody reactive with type I collagen (Col1, 1:250, CL-50111AP-1; Cedarlane), Col2 (1:100, MAB8887; EMD Millipore), type X collagen (ColX, 1:1500, ab49945; Abcam), or ACAN (1:500, AHP0022; ThermoFisher Scientific). Tissues were subsequently incubated with Alexa-594 goat anti-rabbit IgG (1:1000, A11012; Life Technologies), Alexa-488 goat anti-mouse IgG (1:1000, A-11008; Life Technologies), or Alexa-594 goat anti-mouse IgM (1:1500, A-21044; Life Technologies), depending on the primary antibody, for 1 h at room temperature. Tissues were counterstained with DAPI (0.5 mg/mL).
Flow cytometry
Cells were either serially passaged to P2 or plated as P0 overnight (60,000 cells/cm2) in monolayer culture (DMEM with 20% FBS) to allow the cells to recover from the digestion. In a previous study, we showed that P0 chondrocytes do not dedifferentiate until day 3 of culture. 30 P2 cells were harvested at confluency. Accutase (A6964; Sigma-Aldrich) was used to detach cells for flow cytometry analysis.
ENG and TGFBRIII: staining was performed on live cells. P0 or P2 cells were suspended in PBS−/− with 2% FBS (1 × 106 cells/200 μL) and co-stained with 5 μL of ENG APC-conjugated antibody (562408; BD Biosciences) and 10 μL TGFBRIII FITC-conjugated antibody (FAB242F; R&D Systems) for 1 h on ice. DAPI (0.02 μg/mL) was added to assess viability. Unstained cells, APC and FITC only single stained cells, and cells permeabilized with 0.01% Triton and stained with DAPI were used as compensation controls.
ALK5: cells were fixed and permeabilized (Fix & Perm™ Cell Permeabilization Kit, GAS003; ThermoFisher Scientific) as the antibody is reactive with the intracellular portion of the receptor. Briefly, cells were fixed with Medium A (1 × 106 cells/100 μL) for 15 min and subsequently permeabilized with Medium B. Cells were resuspended in PBS−/− with 10% goat serum and 20% FcR Blocking Reagent (130-059-901; Miltenyi Biotec) for 10 min. Primary antibody (1:100, ab31013; Abcam) was added directly and incubated for 30 min at RT. Cells were washed with PBS−/−, resuspended in PBS−/− with 10% goat serum, and incubated for 30 min with secondary antibody (4 μg/mL, Alexa 488, A11008; Invitrogen). Cells were washed and resuspended in PBS−/− for analysis. Hoechst dye was added (2 μg/mL) and flow cytometry was performed. Unstained cells and cells stained with secondary antibody only (no primary) were used as compensation controls.
Briefly, immunostained cells were resuspended in a volume of 50 μL PBS−/− with 1% FBS. Images were then acquired on a 5 laser 12 channel ImageStream MKII imaging flow cytometer at 60 × magnification following ASSIST calibration (Amnis Corporation). Description of the method can be found in the Supplementary Data
Fluorescence-activated cell sorting for ENG
P0 or P2 cells were resuspended in PBS−/− with 2% FBS (10 × 106 cells/200 μL). Five microliters of ENG APC-conjugated antibody (562408; BD Biosciences) was added per 200 μL and incubated for 30 min at RT. Cells were washed once with PBS−/− and resuspended at 5 × 106 cells/mL in a solution of PBS−/− with 25 mM HEPES and 1% heat inactivated FBS. Cells were sorted using the MoFlo Astrios EQ cell sorter (Beckman Coulter, Miami). Unstained cells were analyzed to set the boundary for positive ENG-APC fluorescence. P0 chondrocytes were sorted into three populations, ENG−, ENG+DIM, and ENG+BRIGHT based on fluorescent intensity. Unstained cells were used as controls for gating.
Population doubling time analysis
Population doubling time was calculated using the following formula: DT = duration * log2/log(final concentration) − log(initial concentration). The details are provided in the Supplementary Data.
siRNA-mediated ENG knockdown
When P2 cells reached 80–90% confluency, cells were serum starved for 4 h in DMEM. ENG expression was knocked down using Lipofectamine RNAiMAX Reagent (13778150; ThermoFisher Scientific) as per the manufacturer's instructions and 10 nM (final concentration) of ENG siRNA (s4679, 4392420 or s4677, 4392420; ThermoFisher Scientific) or control siRNA (4390843; ThermoFisher Scientific) in DMEM for 24 h. Cells were either placed in 3D membrane culture or cultured in monolayer culture in fresh media (DMEM supplemented with 20% FBS) for up to 5 days for polymerase chain reaction (PCR) analysis. For 3D membrane culture, cells were seeded at 1 × 106 cells/membrane and cultured as described for up to 2 weeks. Cells were transfected every 5 days to maintain knockdown throughout the 3D culture period.
RNA extraction and reverse transcription polymerase chain reaction
Detailed methods and primer sequences can be found in the Supplementary Data. Total RNA was extracted using TRIzol reagent (15596026; Thermo Fisher Scientific, Canada) and reverse transcribed to cDNA using Superscript II Reverse Transcriptase (18064014; Invitrogen, Canada). Relative real-time PCR based on SYBR Green (Invitrogen) was performed using primers specific for 18S rRNA or ENG. Each reaction contained 40 ng of cDNA, 6 μL of SYBR Green master mix, and 1 μL each of forward and reverse primers (10 mM). mRNA levels were determined from Cq values according to Pfaffl's mathematical model for relative real-time PCR. 31 18S rRNA was used for normalization.
Statistical analysis
Unless otherwise stated, three independent experiments were performed using cells or tissues from three different patient samples (n = 3). Each condition was done in triplicate. The data was pooled, expressed as mean ± SEM, and analyzed using a paired t-test or one-way analysis of variance with Tukey's Post Hoc if more than two conditions. Significance was assigned at p < 0.05.
Results
Passaging enables chondrocytes to form hyaline-like cartilaginous tissues regardless of whether cells were obtained from High OA or Low OA grade tissues
Macroscopically, Low OA tissues appeared intact, while High OA tissues were thinner and irregular in appearance (Fig. 1A). Histological sections of AC from these regions were evaluated using the OARSI histological grading scale 25 to determine if macroscopic examination was sufficient to differentiate minimally altered (Low OA) from severely degraded (High OA) cartilage (Fig. 1). Histologically, tissues derived from what was considered High OA regions of the joint microscopically showed deep fibrillations, tissue erosion, loss of proteoglycans, and chondrocyte cloning (Fig. 1C, E), whereas Low OA (Fig. 1B, D) cartilage showed, at most, only minor surface discontinuity and the proteoglycan content appeared intact as determined by toluidine blue staining (Fig. 1D, E). As the grades were significantly different between Low OA and High OA tissues (Fig. 1F), this confirms that macroscopic examination could differentiate between low- and high-grade OA cartilage.

In a previous study, we showed that primary chondrocytes derived from Low OA cartilage did not form cartilaginous tissues when cultured in SFM in the presence of TGFβ3 when grown in 3D culture. 11 In this study, we found that primary high-grade OA chondrocytes, similar to low-grade OA cells, also were unable to form cartilage tissue under these conditions. There were no significant differences in the extracellular matrix-poor tissues generated by P0 Low OA or High OA chondrocytes with respect to the amount of sulfated GAG (Fig. 2A) or collagen accumulation (Fig. 2B). Histologically, the tissues were thin, noncartilaginous (Fig. 2C, D) and little, if any, proteoglycan accumulated as shown by toluidine blue staining (Fig. 2E, F).
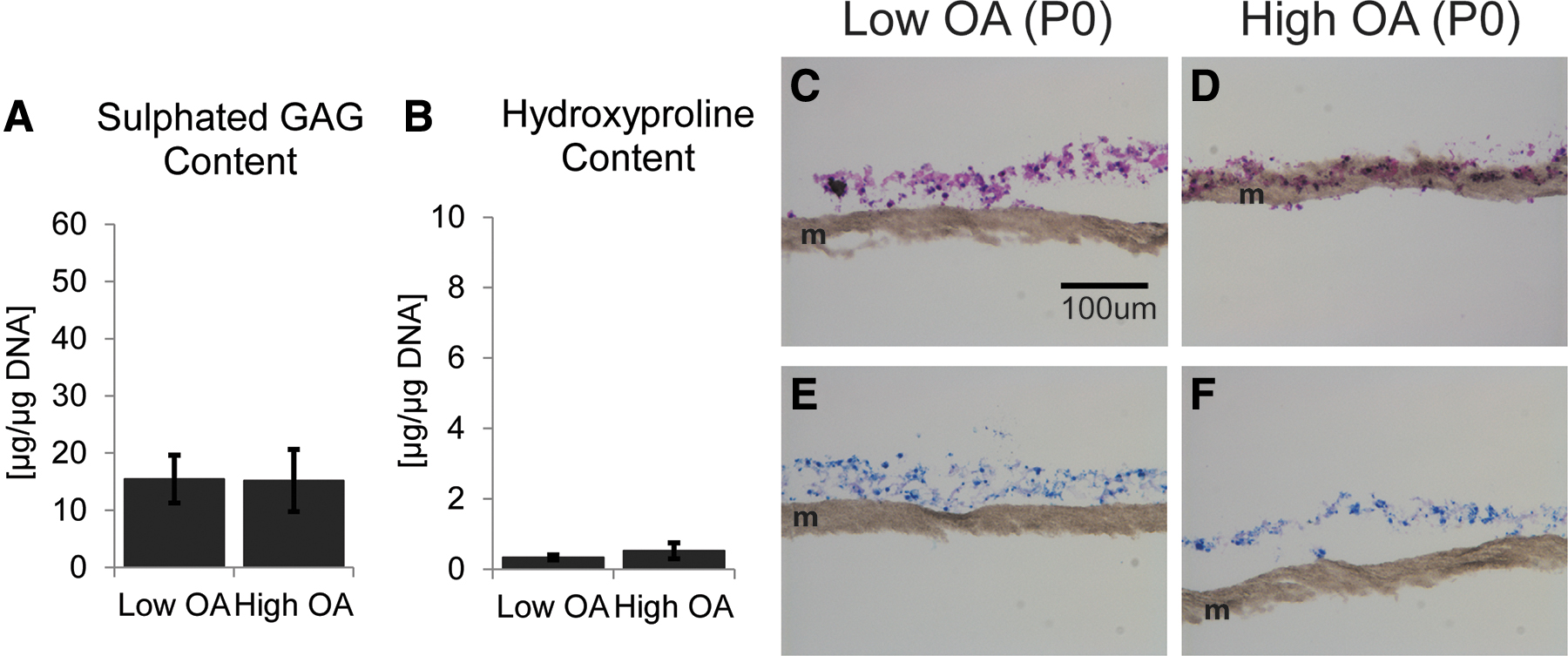
Primary chondrocytes do not form cartilaginous tissues in vitro regardless of the grade of OA. Primary chondrocytes were cultured in 3D culture on membranes in SFM supplemented with 10 ng/mL TGFβ3 for 3 weeks.
After serial passaging, however, P2 chondrocytes cultured under the same conditions accumulate approximately three times more proteoglycan (Figs. 2A and 3A) and 12 times more collagen (Figs. 2B and 3B) than P0 cells. Histological evaluation (Fig. 3C, F) and immunohistochemical studies (Fig. 3G, H) show no difference between the cartilaginous tissues formed by Low OA and High OA chondrocytes as both accumulate Col2 and ACAN throughout the tissue. Neither Col1 nor ColX was detected by immunohistochemistry in either group at 3 weeks.

Passaged (P2) chondrocytes form cartilaginous tissues with similar amounts of matrix regardless of whether cells were obtained from High or Low OA tissues. P2 chondrocytes were cultured in 3D culture in SFM supplemented with 10 ng/mL TGFβ3 for 3 weeks.
Cells from Low OA and High OA show increased ENG expression with serial passaging, but no change in ALK5 or TGFβRIII levels
To investigate why P2 and not P0 chondrocytes form cartilaginous tissues in culture with TGFβ3, image flow cytometry was used to assess expression of the chondrogenesis-promoting TGFβ receptor ALK5, and co-receptors ENG and TGFβRIII in P0 and P2 chondrocytes derived from High and Low OA (Fig. 4). The number of cells expressing ENG as well as the density of ENG/cell (assessed by median fluorescent intensity value) increased significantly with passaging to P2 for both Low and High OA cells (Fig. 4A). No significant difference in Eng expression between the two grades was detected in either the P0 or P2 cells. TGFβRIII was minimally expressed (<5%) in both P0 and P2 cells and the receptor density decreased after passaging for both Low OA and High OA cells (Fig. 4B). ALK5 was expressed in the majority of P0 cells (∼95%), and this expression was maintained in P2 cells with no significant change in the density of ALK5 (Fig. 4C). No significant grade-specific differences in the expression of ALK5 were identified, as Low OA and High OA populations showed similar levels of expression.
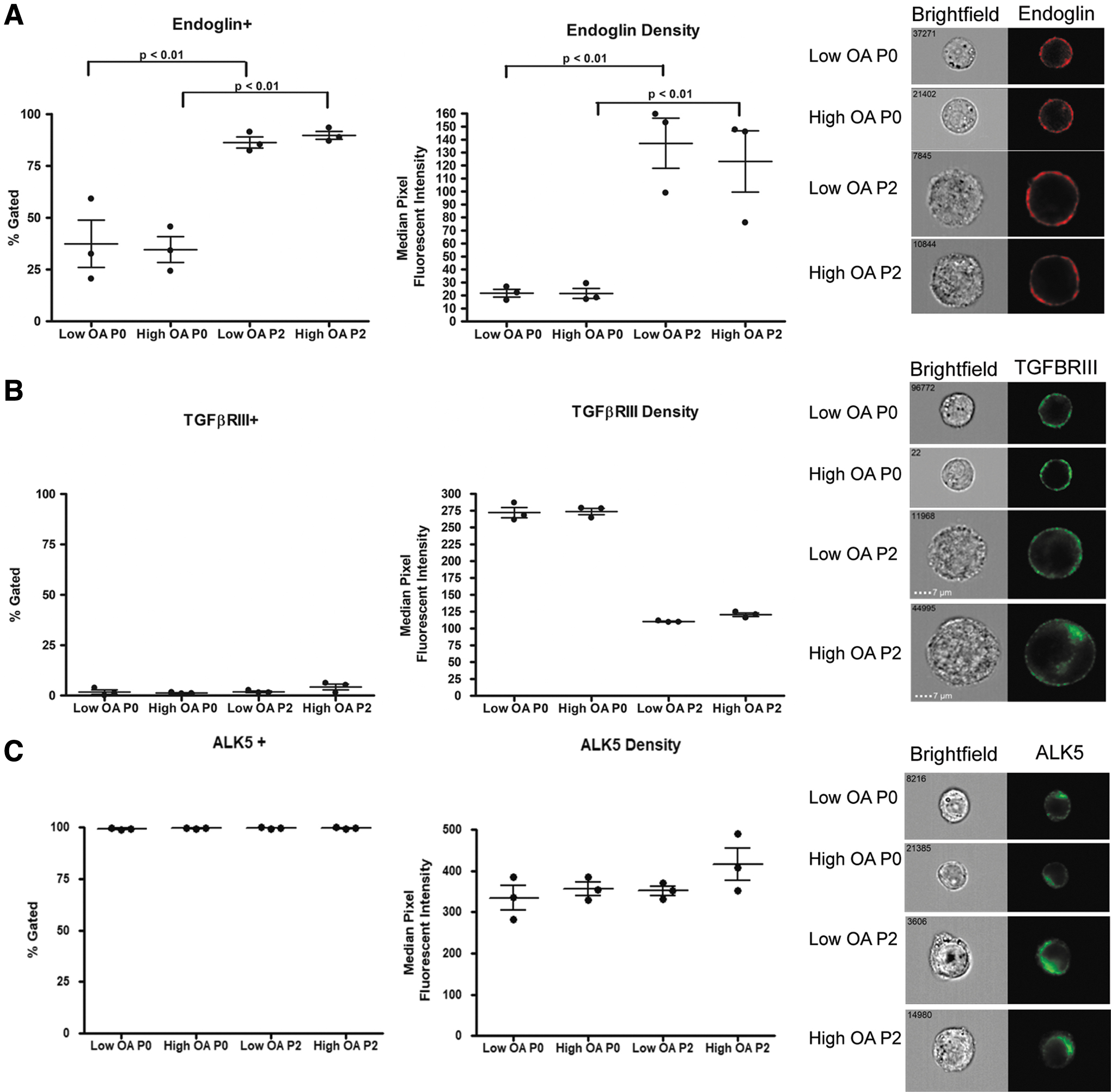
Image flow cytometry shows no grade-specific differences in expression of ENG, TGFBRIII, and ALK5. P0 or P2 cells were assessed using image flow cytometry following staining with antibodies reactive with
ENG expression is acquired with serial passaging in monolayer
As the number of cells expressing ENG as well as the density increased significantly with passaging (Fig. 4A), we examined whether this phenotype was acquired because of monolayer cell culture and passaging, or because the ENG+ population proliferated faster. Fluorescence-activated cell sorting was used to isolate P0 chondrocytes into three populations, ENG−, ENG+DIM, or ENG+BRIGHT. These were then placed in monolayer culture separately. To assess proliferation, growth curves were generated (Supplementary Fig. S1). Analysis of doubling time revealed that the ENG+BRIGHT proliferated the fastest of the three groups (ENG− = 5.4 days, ENG+DIM = 4.7 days, and ENG+BRIGHT = 4.3 days). Given the minimal differences in doubling time, we then determined if the P0 ENG− population would become ENG+ with passaging. The vast majority of the cells became ENG+ by P2 (Fig. 5A), suggesting that cells acquired ENG expression with passaging. Unsorted cells, which were used as controls, became predominantly ENG+. When P2 cells obtained from either ENG-sorted (ENG−, ENG+DIM or ENG+BRIGHT) or unsorted P0 cells were seeded separately in 3D membrane culture in the presence of TGFβ3 for 3 weeks, they all formed cartilage tissue. No significant difference in the amount of collagen or proteoglycan accumulated was detected between the different tissues (Fig. 5B). Histological assessment and immunohistochemistry showed no difference between the tissues formed by the four different populations of cells with respect to presence and distribution of Col2, Col1, ColX, and ACAN (Fig. 5C).

P0 chondrocytes become ENG+ as a result of serial passaging.
ENG knockdown prevents TGFβ-mediated cartilaginous tissue formation in vitro by P2 chondrocytes
Given the significant changes in ENG expression, the role of ENG in chondrogenesis by passaged chondrocytes was examined. P2 cells were transfected with ENG specific siRNA or control siRNA for 24 h and then seeded in 3D culture on membranes in SFM in the presence of TGFβ3. To maintain knockdown, cultures were transfected every 5 days and cultured for up to 2 weeks. Studies revealed that following a single transfection, mRNA expression of ENG was significantly decreased for up to 5 days (Fig. 6A). Lower ENG protein expression in ENG siRNA-treated cell populations was confirmed by flow cytometry (Fig. 6B) at 3 days post-transfection. Approximately 60% of cells treated with ENG siRNA completely lost protein expression of ENG. In addition, the median fluorescent intensity (MFI), indicative of the amount of protein, was approximately eight times lower in ENG siRNA-treated cells versus control siRNA treated. Tissues formed by ENG siRNA-transfected cells were thinner at both 7 (Fig. 6C) and 14 days (Supplementary Fig. S2) and contained significantly less sulfated GAG (Fig. 6D) than tissues formed by control cells (transfected with control siRNA). Grading using the Bern System showed that tissues formed by control cells had a greater average score than tissues formed by ENG siRNA-treated cells (7.1 ± 1.0 vs. 4.1 ± 0.6, respectively). 29 DNA content and OH-proline content, which is low at this time point, were similar in the two tissues at 7 days (Fig. 6D).
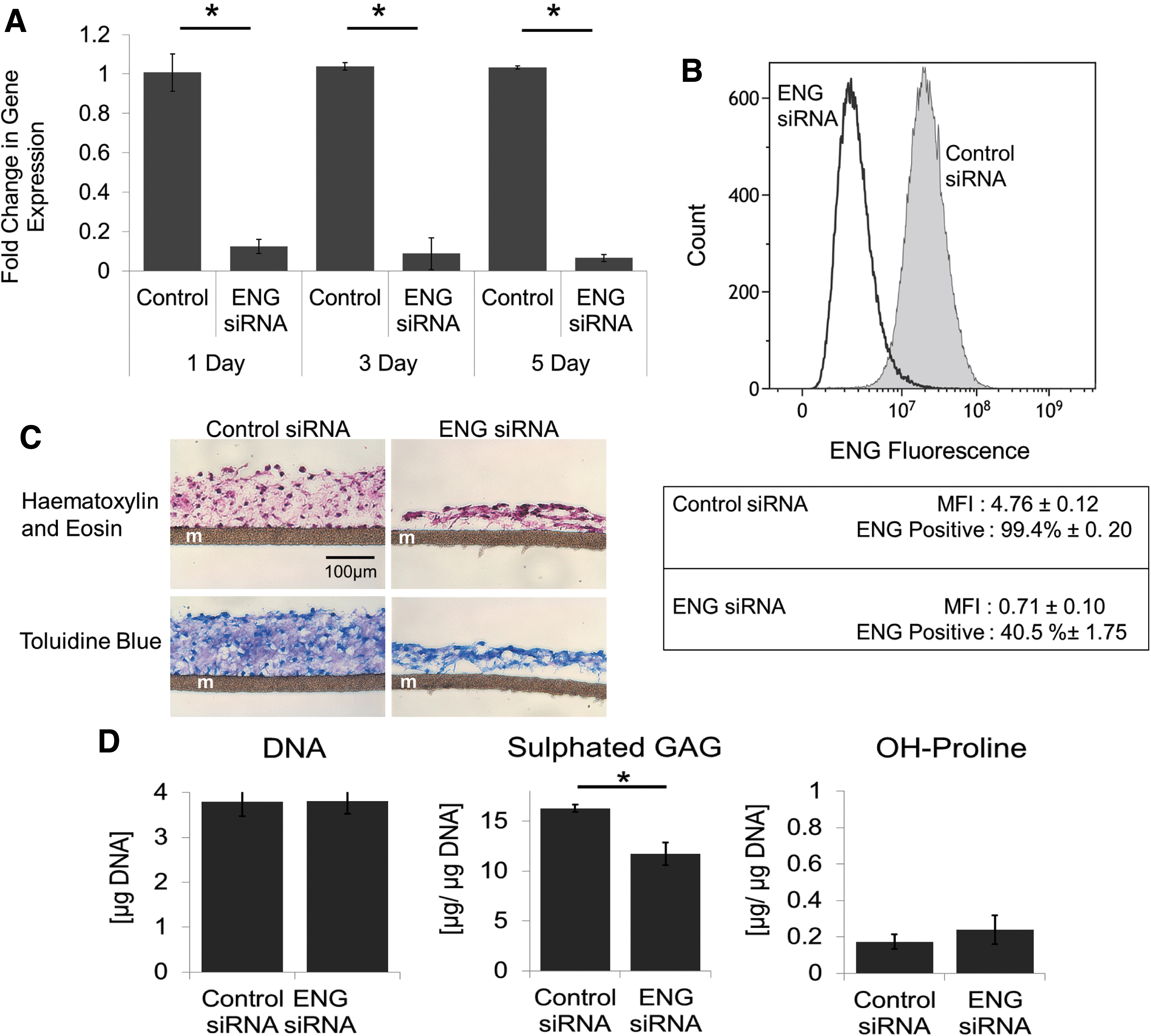
Knockdown of ENG modulates cartilaginous tissue formation by P2 chondrocytes.
Discussion
This study demonstrated that the grade of OA from which chondrocytes are obtained does not influence the ability of the cells to form cartilaginous tissue in vitro. After serial passing in monolayer followed by culture in chondrogenic conditions 11 in the presence of TGFβ3, cells from Low OA and High OA formed tissues of similar quality as determined by histology, immunohistochemical staining of matrix molecules, and biochemistry. In contrast, primary (P0) cells were unable to form cartilage under these chondrogenic conditions. The P0 cells expressed ALK5 and low levels of the co-receptors endoglin and TGFβRIII. Interestingly, with passaging, no significant changes in ALK5 or TGFβRIII were detected, but there were significantly more cells that expressed endoglin at a higher density/cell. Knockdown of ENG in P2 cells resulted in cells that were unable to generate cartilaginous tissue. This indicates an important regulatory role for endoglin in cartilage tissue formation in vitro. Our data also suggest that the presence of ALK5 alone is not sufficient for chondrogenesis, as ∼95% of P0 cells expressed this receptor and yet could not form tissue.
Although both normal and passaged chondrocytes have been shown to express ENG by others,32–34 to the best of our knowledge, no study has been performed examining the role of ENG in modulating chondrogenesis by passaged human articular chondrocytes. In keeping with our findings, investigations using adipose and synovium derived stem cells have shown that cells enriched for ENG (also known as CD105) expression demonstrate enhanced chondrogenic potential.23,24 Cleary et al. showed that ENG(−) MSCs had a diminished capacity for attachment and proliferation and lacked the ability to form pellets during chondrogenic differentiation. 35 In vivo, Asen et al. found that ENG-expressing cells were the predominate cells contributing to repair tissue formation in osteochondral defects in minipigs. 36 Interestingly, this group utilized TGFβ-releasing scaffolds and demonstrated an increase in ENG+ cells in the repair tissue, along with enhanced repair. Although there is one report suggesting that ENG positivity does not predict chondrogenesis potential, this study utilized MSCs at higher passage number (P4) 35 ; in conglomerate, the studies on progenitor cells support our observation that ENG functions to promote TGFβ-mediated chondrogenesis.
It is not known why ENG expression increases with cell passaging. There are three possible explanations, ENG has been shown to be upregulated by TGFβ and hypoxia. 37 However, in this study, cells were passaged in normoxia and TGFβ was not a component of the culture media. Therefore, it is unlikely these factors are playing a role. Several groups have demonstrated that chondrocytes become MSC-like when passaged in monolayer culture.38,39 As ENG is a known in vitro marker of MSCs, this may explain why P2 chondrocytes acquire expression of this protein. Third, ENG has an integrin binding domain, 40 which has been shown to play a role in cell adhesion and migration in other cell types.41,42 Upregulation of ENG with adhesion has been demonstrated for macrophages, 42 so it is possible that in our system, increased ENG expression in P2 cells may be a result of cell adhesion with serial passaging in monolayer culture.
It is likely that ENG alone does not predict chondrogenic potential and that ALK5 is also needed to mediate chondrogenesis as expression levels did not vary between P0 and P2 cells. ENG enhances TGFβ signaling by interacting with one of the two TGFβ receptors, ALK5 and ALK1, which signal through SMAD2/3 or SMAD1/5/8, respectively. Signaling through ALK5-SMAD2/3 in human chondrocytes has been shown to be responsible for an anabolic effect, while signaling through ALK1-SMAD1/5/8 is associated with catabolism.14,17 Investigations by Pomeraniec et al. demonstrated that in endothelial cells, ENG forms complexes with both TGFβRII and ALK5, bringing these receptors into close proximity. 43 Studies in other cell types have shown that ENG can act through ALK5-SMAD2 signaling 22 and differentially regulate signaling in favor of ALK5. 44 Finnson et al. demonstrated that in primary chondrocytes, ENG enhanced signaling by ALK1 and SMAD1/5/8 and inhibited signaling through the ALK5 and SMAD2/3 pathway. 32 In MSCs, which express ENG, chondrogenesis appears to be regulated through SMAD2/3 signaling.45,46 Thus, ENG may be acting in the P2 cells in our system in a similar manner to enhance signaling through ALK5-SMAD2/3 pathway to mediate hyaline cartilaginous tissue formation. However, it is possible that ENG has multiple roles in cells and acts in a context- and tissue-specific manner. The mechanism by which it may be acting in our system requires further investigation.
TGFβRIII has also been shown to function to enhance TGFβ signaling.21,47 TGFβRIII binds all three isoforms of TGFβ, with highest affinity for TGFβ2,20,47 which signals poorly in the absence of TGFβRIII. 21 In this study, we showed that human articular chondrocytes had few cells expressing TGFβRIII. This correlates with the low amounts of TGFβ2 present in adult normal and OA chondrocytes. 48 Contrary to our findings, Parker et al. show abundant TGFβRIII expression by immortalized chondrocyte cell lines C-28/I2 and tsT/AC62. 19 It is possible that the process of immortalization may affect TGFβRIII expression, resulting in levels that differ from those we detected in human articular chondrocytes. Interestingly, a recent study by Zheng et al. showed that downregulation of TGFβRIII in human MSCs significantly enhanced TGFβ3-induced chondrogenic differentiation. 49 Low levels of this co-receptor, as present in the cells grown in our culture system, may be playing a role to support chondrogenesis by passaged chondrocytes. Further characterization will be required to confirm this.
This study has several limitations. First, cells that have undergone more extensive passaging (beyond P2) and may be senescent were not investigated for ENG expression or their ability to form cartilaginous tissues in these culture conditions. Second, overexpression of ENG in P0 chondrocytes, which have low endogenous ENG expression, was not examined and should be explored in future studies to determine whether this can restore the ability of these cells to form cartilaginous tissue. Third, downstream signaling pathways in the presence of high or low expression of ENG need to be determined to understand how ENG is regulating cartilaginous tissue formation. Finally, patient factors that may play a role in regulating endogenous ENG levels, such as age, gender and chronic illness, should be investigated in future work.
In summary, chondrocytes obtained from OA cartilage, either high or low grade, acquire a similar ALK5 and ENG profile after two passages, which is permissive for chondrogenesis. The presence of ALK5 alone was insufficient to allow chondrocytes to form cartilage tissue in the presence of TGFβ3, as both P0 and P2 cells contained ALK5 in equivalent abundance, yet P2 cells, which had increased levels of ENG relative to P0 cells, were able to form cartilaginous tissue. Knockdown studies confirmed that this response was dependant on the upregulation of ENG. Clinically, the use of chondrocytes, which express both high levels of ALK5 and ENG, should be explored in future studies of cartilage repair. Developing therapeutic agents or scaffolds that promote upregulation of these two molecules to sufficient levels might also be beneficial.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.