
Abstract
Decellularized adipose tissue (DAT) scaffolds represent a promising cell-instructive platform for soft tissue engineering. While recent work has highlighted that mesenchymal stromal cells, including adipose-derived stromal cells (ASCs), can be combined with decellularized scaffolds to augment tissue regeneration, the mechanisms involved require further study. The objective of this work was to probe the roles of syngeneic donor ASCs and host-derived macrophages in tissue remodeling of DAT scaffolds within an immunocompetent mouse model. Dual transgenic reporter mouse strains were employed to track and characterize the donor ASCs and host macrophages within the DAT implants. More specifically, ASCs isolated from dsRed mice were seeded on DAT scaffolds, and the seeded and unseeded control scaffolds were implanted subcutaneously into MacGreen transgenic mice for up to 8 weeks. ASC seeding was shown to augment cell infiltration into the DAT implants at 8 weeks, and this was linked to significantly enhanced angiogenesis relative to the unseeded controls. Immunohistochemical staining demonstrated long-term retention of the syngeneic donor ASCs over the duration of the 8-week study, providing evidence that the DAT scaffolds are a cell-supportive delivery platform. Notably, newly formed adipocytes within the DAT implants were not dsRed+, indicating that the donor ASCs supported fat formation through indirect mechanisms. Immunohistochemical tracking of host macrophages through costaining for enhanced green fluorescent protein with the macrophage marker Iba1 revealed that ASC seeding significantly increased the number of infiltrating macrophages within the DAT implants at 3 weeks, while the fraction of macrophages relative to the total cellular infiltrate was similar between the groups at 1, 3, and 8 weeks. Consistent with the tissue remodeling response that was observed, western blotting demonstrated that there was significantly augmented expression of CD163 and CD206, markers of constructive M2-like macrophages, within the ASC-seeded DAT implants. Overall, our results demonstrate that exogenous ASCs enhance tissue regeneration within DAT scaffolds indirectly through multimodal mechanisms that include host cell recruitment and immunomodulation. These data provide further evidence to support the use of decellularized scaffolds as a delivery platform for ASCs in tissue engineering.
Impact statement
Decellularized tissues have been investigated as proregenerative scaffolds in tissue engineering. This study corroborates evidence demonstrating improved tissue remodeling and regeneration in scaffolds preseeded with adipose-derived stromal cells (ASCs). Importantly, this work investigated ASC-mediated mechanisms of tissue regeneration in decellularized adipose tissue (DAT) scaffolds, and showed that differences in the macrophage response contributed to enhanced tissue remodeling in the ASC-seeded DAT scaffolds relative to unseeded controls. The donor ASCs indirectly promoted tissue regeneration through multimodal mechanisms that include cell recruitment and immunomodulation. The results provide important mechanistic and preclinical data to support the continued development of this strategy for soft tissue augmentation and reconstructive procedures.
Introduction
Extracellular matrix (ECM)-derived bioscaffolds are of great interest in tissue engineering due to their ability to direct cell behavior and promote tissue regeneration.1,2 These scaffolds are derived using decellularization methods that remove antigenic cellular components while retaining important compositional, structural, and biomechanical cues of the native tissue microenvironment. 3 Due to their favorable safety profiles and capacity to stimulate in situ tissue regeneration, several decellularized ECM products are currently in clinical use, including those derived from human or animal small intestinal submucosa, bladder, dermis, and heart valves. 4
Decellularized adipose tissue (DAT) scaffolds have emerged as a promising tissue-specific regenerative platform for reconstructive and cosmetic applications in plastic surgery.5,6 Our laboratory has pioneered a detergent-free protocol to derive human DAT 7 that can be employed as a scaffold in its intact form,7,8 or formulated in a variety of scaffold formats including porous foams,8,9 hydrogels,10,11 and microcarriers. 12 We have shown that the DAT displays similar biomechanical properties to native breast adipose tissue,13,14 and also retains nonstructural bioactive ECM proteins including chemotactic, proangiogenic, and proadipogenic factors. 15
In vitro studies using human adipose-derived stromal cells (ASCs) demonstrated that the DAT strongly supports adipogenesis and is also adipoinductive.7,8,10–12 Further, several studies have shown that DAT scaffolds support neovascularization and adipogenesis after subcutaneous implantation in rodent models.16–18 Notably, a recent clinical study by Kokai et al. reported that an injectable DAT formulation showed no major adverse events, and supported volume retention and adipogenesis at 4 months postdelivery subcutaneously within the dorsal wrist of healthy human volunteers. 19
While decellularized scaffolds are promising for off-the-shelf tissue-engineering applications, emerging data have indicated that these materials can serve as effective delivery platforms for mesenchymal stromal cell (MSC) populations, including ASCs. 2 Furthermore, this combination strategy may be more effective for soft tissue regeneration compared with either scaffolds or cells alone.16,20,21 Indeed, ASCs are known to secrete a variety of chemotactic, proangiogenic, and immunomodulatory factors that can augment tissue repair and regeneration. 22 In the context of adipose tissue regeneration, xenogeneic 18 and allogeneic 23 ASCs incorporated into adipose ECM hydrogels enhanced angiogenesis and adipogenesis relative to ECM hydrogels alone within an athymic mouse model and immunocompetent rat model, respectively.
As inflammation may play an important role in neoadipogenesis,24,25 our group is interested in investigating the immunomodulatory effects of ASCs delivered within DAT scaffolds. In our previous study using an immunocompetent rat model, allogeneic ASCs enhanced angiogenesis and adipogenesis within intact DAT scaffolds. 26 In addition, ASC seeding modulated the inflammatory response as evidenced by an increased number of interleukin-10+ (IL-10+) cells and an increased fraction of macrophages expressing the proregenerative M2-like macrophage marker CD163. 26 Notably, cell tracking through fluorescence in situ hybridization revealed that the allogeneic donor ASCs were greatly diminished within the DAT scaffolds by 8 weeks postimplantation, supporting their capacity to aid in tissue regeneration through indirect paracrine mechanisms. 26
The objective of this study was to further probe cell recruitment and immunomodulation as mechanisms of tissue regeneration in ASC-seeded DAT scaffolds. We employed a syngeneic dual transgenic reporter mouse system to track and characterize both donor ASCs and infiltrating macrophages in tandem within DAT scaffolds. Syngeneic ASCs were used in this study to increase the genetic similarity of the donor cells and improve compatibility with host immune cells. We hypothesized that the exogenous ASCs would enhance angiogenesis and adipogenesis within DAT scaffolds by modulating cell recruitment and inflammation.
Materials and Methods
Materials
Chemicals and reagents were purchased from Sigma Aldrich (Oakville, Canada) unless otherwise stated.
Animals
All animal studies were performed in accordance with the Canadian Council on Animal Care (CCAC) guidelines and were approved by the Animal Care Committee at Western University (Protocol #2015-049). Mice were housed in clean barrier facilities under humidity- and light-controlled conditions, and fed ad libitum with free access to water. ASCs were isolated from 8- to 12-week-old male dsRed mice (B6.Cg-Tg(CAG-DsRed*MST)1Nagy/J, C57BL/6J background; Jackson Laboratory, Bar Harbor). Scaffold implantation studies were performed using a total of 33 hemizygous 8- to 12-week-old female MacGreen transgenic mice (B6N.Cg-Tg(Csf1r-EGFP)1Hume/J, C57BL/6J background; Jackson Laboratory), in which cells of the myeloid lineage express enhanced green fluorescent protein (EGFP). 27 An overview of the experimental design is provided in Figure 1.

Overview of experimental design, timeline, and assays. ASC, adipose-derived stromal cell; H, histology to evaluate cell infiltration, angiogenesis, and adipogenesis; IF, immunofluorescence to examine macrophage infiltration; IHC, immunohistochemistry to visualize dsRed+ ASC retention; WB, western blotting to measure expression levels of macrophage phenotypic markers.
ASC isolation and culture
ASCs were isolated from dsRed mice according to published methods 28 with the following modifications: a 100 μm filter (Corning, Inc., Corning) was used to strain the cell suspension after digestion, and Primaria™ T25 flasks (Corning) were used for initial plating of cells at passage zero (P0). Inguinal fat pads were pooled from 4 to 7 male mice (8–12 weeks of age) for each isolation. ASCs were cultured in proliferation medium consisting of Dulbecco's modified Eagle's medium:Ham's F12 with 10% fetal bovine serum (Gibco, Invitrogen, Burlington, Canada) and 1% penicillin–streptomycin (pen-strep; Gibco, Invitrogen) at 37°C and 5% CO2. The cells were passaged using 0.25% trypsin/0.1% ethylenediaminetetraacetic acid (trypsin/EDTA; Gibco, Invitrogen), counted on a hemocytometer, and replated at 6000–7000 cells/cm2 on standard tissue culture polystyrene flasks (Corning) for subsequent passages beyond P0. Media changes were performed every 2–3 days, and P2–P3 cells were used in all studies.
ASC identity was confirmed as CD90+CD29+CD44+CD31-CD45- (n = 3 independent ASC isolations; Supplementary Fig. S1) by flow cytometry following published protocols. 29 The following monoclonal APC-conjugated antibodies from eBioscience (Ottawa, Canada) were used for staining: anti-CD29 (1 μg/mL; cat. 17-0291), anti-CD31 (2 μg/mL; cat. 17-0311), anti-CD44 (0.24 μg/mL; cat. 17-0441), anti-CD45 (0.5 μg/mL; cat. 17-0451), and anti-CD90.2 (0.24 μg/mL; cat. 17-0902). Live singlets were gated based on the forward and side scatter properties, with further gating performed using fluorescence minus one control, including wild-type murine ASCs isolated from nontransgenic 8–12 week old male C57BL/6J mice that were cultured in parallel to the dsRed+ cells.
The in vitro adipogenic differentiation capacity was also verified (n = 3 technical replicates, n = 3–4 experiments from independent ASC isolations; Supplementary Fig. S2) by oil red O staining and glycerol-3-phosphate dehydrogenase (GPDH) enzyme activity according to published protocols.30,31 Adipogenic differentiation medium consisted of proliferation medium supplemented with the following components as adapted from the literature: 28 10 mg/mL troglitazone (first 72 h only), 0.25 mM IBMX (first 72 h only), 33 μM biotin, 17 μM D-pantothenic acid, 10 mg/mL human insulin, and 1 μM dexamethasone.
DAT scaffold fabrication and seeding
Subcutaneous human adipose tissue was obtained with informed consent from elective breast reduction or abdominoplasty surgeries at the Cumberland Laser Clinic, St. Joseph's Hospital, or University Hospital in London, Ontario with approval from the Research Ethics Board at Western University (HSREB #105426). Tissue decellularization was performed using an established 5-day detergent-free protocol. 7 After decellularization, the DAT was frozen at -80°C and lyophilized. The lyophilized DAT was cut into individual scaffolds (7.0 ± 0.5 mg), rehydrated in deionized water, and decontaminated with repeated 70% ethanol rinses, followed by PBS washes, as previously described. 8
Before seeding, the scaffolds were equilibrated in ASC proliferation medium overnight at 37°C and 5% CO2. Next, the ASCs were trypsinized and counted using a hemocytometer. Each DAT scaffold was transferred into a 15 mL vented cap conical tube containing 3 mL of proliferation medium and 2 × 106 ASCs. The scaffolds were dynamically seeded for 24 h using a BenchWaver™ 3D rocker (Mandel, Guelph, Canada) at 90 rpm (37°C, 5% CO2). To visualize ASC attachment on the DAT scaffolds, samples collected at 24 h postseeding were frozen over dry ice in Tissue-Tek® O.C.T. Compound (Sakura® Finetek, Torrance, Canada). The scaffolds were then stored at −80°C for subsequent sectioning (5–7 μm sections) and mounted with DAPI mounting medium (Abcam, Toronto, Canada). Images of the stained sections collected using the EVOS® FL Imaging System (Invitrogen, Ottawa, Canada) showed that the ASCs were predominantly localized within the periphery of the DAT scaffolds, with fewer cells distributed heterogeneously toward the interior (Supplementary Fig. S3).
Scaffold implantation and tissue harvest
Scaffolds were implanted in subcutaneous pockets on the dorsa of mice as previously described.8,26 One ASC-seeded and one unseeded DAT scaffold were implanted contralaterally in each mouse. At 72 h, 1, 3, and 8 weeks postimplantation, mice were euthanized by CO2 overdose, and the scaffolds were excised within their surrounding tissues. For histological processing (n = 6 mice per time point), the excised scaffolds were bisected, and half of each implant was fixed in 10% neutral buffered formalin for 24 h before being transferred to 70% ethanol for subsequent paraffin embedding. The remaining half of the scaffold was frozen over dry ice in O.C.T. Compound. For western blot experiments (n = 4–5 mice/time point), the entire scaffold was excised, snap-frozen in liquid nitrogen, and stored at -80°C until processing.
Masson's trichrome staining
Sections (5–6 μm) from paraffin-embedded tissues were deparaffinized in xylene and rehydrated in a graded ethanol series before Masson's trichrome staining.8,26 The tissue sections were then dehydrated in a graded ethanol series and mounted with Permount® (ThermoFisher). Slides were scanned at 20 × magnification using an Aperio ImageScope system and software (Vista). The number of erythrocyte-containing blood vessels, as well as the fraction of the implants infiltrated by cells or remodeled into adipose tissue, was quantified relative to the total implant area using Spectrum ImageScope software (Vista). The analysis was performed blinded, and data points were generated using the mean values from three different tissue sections/scaffold, each separated by 100–150 μm of tissue depth.
Immunohistochemical detection of dsRed+ donor cells
For dsRed staining, 5 μm sections from paraffin-embedded tissues were deparaffinized and rehydrated in a graded ethanol series, followed by heat-mediated antigen retrieval in citrate solution (pH 6.0) at 95°C for 30 min. Tissue sections were blocked (1 h at room temperature [RT] in 10% goat serum +0.1% Tween-20 in Tris-buffered saline [TBS]) and incubated overnight at 4°C with a rabbit polyclonal anti-RFP primary antibody (1:200 in TBS +5% goat serum +0.1% Tween-20, cat. 600-401-379; Rockland Immunochemicals, Limerick). The tissue sections were then washed in TBS, and endogenous peroxidases were quenched in 3% H2O2 for 15 min at RT. After applying a goat anti-rabbit HRP-conjugated secondary antibody (1:300 in TBS, 30 min at RT, cat. Ab6721; Abcam), sections were washed in TBS and incubated with 3,3′-diaminobenzidine (DAB) for 5 min at RT followed by counterstaining with Harris hematoxylin and lithium carbonate, dehydration in a graded ethanol series, and mounting with Permount (ThermoFisher).
Slides were scanned at 20 × magnification using an Aperio ImageScope system and software (Vista), and quantitative analysis of positively stained pixels was performed using Fiji software, an image processing platform based on ImageJ. 32 Blinded analysis was performed using three different tissue sections/scaffold each separated by 100–150 μm; data points represent the mean value obtained from the three tissue sections. DAT scaffolds implanted into dsRed+ mice and wild-type mice were used as positive and negative controls, respectively, along with no primary controls to confirm staining specificity.
Immunohistochemical assessment of macrophage infiltration
To examine infiltrating macrophages, immunohistochemical staining was performed for the macrophage marker Iba1 with costaining for EGFP. Five-micrometer sections from fresh-frozen tissues were fixed in acetone at −20°C for 10 min. After washing in TBS, the sections were blocked for 1 h at RT in TBS +10% goat serum +0.1% Tween-20. The primary antibody for Iba1 (1:100 dilution in blocking solution, rabbit monoclonal, cat. ab178846; Abcam) was added for overnight incubation at 4°C. The tissue sections were washed in TBS, and the secondary antibody (1:500 in TBS, goat antirabbit, Alexa 594, cat. ab150080; Abcam) was applied for 30 min at RT. After three TBS washes, tissue sections were blocked once more using TBS +10% rabbit serum +0.1% Tween-20 for 1 h at RT. An Alexa 647-conjugated rabbit polyclonal anti-EGFP antibody (1:100 dilution in blocking solution, cat. A-31852; ThermoFisher) was applied and incubated overnight at 4°C.
The tissue sections were washed and stained with Hoechst (ThermoFisher), and then mounted with Fluoroshield™ Mounting Medium (Abcam). Blinded quantitative analysis was performed in ImageJ by manually counting Hoechst+ cells that were positive for each marker. Average counts were recorded from six nonoverlapping images captured along the implant periphery (within 500 μm of the implant boundary) at three different tissue depths (100–150 μm in separation). All images were taken at 20 × magnification with the EVOS FL Imaging System (Invitrogen). Tissue-positive mouse spleen controls and no primary controls were used to confirm staining specificity for each marker.
Western blots
Snap-frozen western blot samples were placed in cold sample buffer containing 20.8 mM Tris-HCl (pH 6.8), 6.7% glycerol, 0.7% sodium dodecyl sulfate (SDS), 1% dithiothreitol, 1.7% β-mercaptoethanol, and 0.001% bromophenol blue in distilled water. The samples were then homogenized, denatured (95°C, 10 min), and centrifuged (12,000 g, 10 min). The protein concentration of sample supernatants was measured using the Pierce BCA 660 nm Protein Assay Reagent supplemented with the Ionic Detergent Compatibility Reagent (ThermoFisher). Supernatants were then diluted using 4 × Laemmli buffer, aliquoted to avoid freeze-thaw cycles, and stored at −20°C.
Western blot analysis was performed using Bio-Rad stain-free technology. 33 The BLUelf Prestained Protein ladder (FroggaBio, Toronto, Canada) and 200 μg of sample protein/well were loaded into 12% polyacrylamide gels prepared using the TGX Stain-Free™ FastCast™ Acrylamide Kit (Bio-Rad) according to the manufacturer's instructions. The gel was run using the Mini-PROTEAN® Electrophoresis System (Bio-Rad) at 250 V in running buffer (25 mM Tris, 190 mM glycine, 0.1% SDS). The TGX Stain-Free gels were then activated, and total protein was visualized (Gel Doc™ EZ Imager; Bio-Rad), followed by transfer to a polyvinylidene difluoride (PVDF) membrane using the Trans-Blot® Turbo™ Transfer System (1.3 A, 25 V, 10 min; Bio-Rad) and a Trans-Blot® Turbo™ RTA LF PVDF Transfer Kit (Bio-Rad). 33
After the transfer step, membranes were washed three times in TBS +0.1% Tween-20 (5 min each, 100 rpm on an orbital shaker), blocked (TBS +5% bovine serum albumin [BSA] + 0.1% Tween-20 for 1 h at RT, 100 rpm on an orbital shaker), and incubated with the primary antibody against CD163 (1:2000 dilution, rabbit monoclonal, cat. ab182422; Abcam) or CD206 (1:1000 dilution, rabbit polyclonal, cat. ab64693; Abcam) in TBS +1% BSA +0.1% Tween-20 overnight at 4°C under agitation (100 rpm on an orbital shaker). After three washes in TBS +0.1% Tween-20, the secondary antibody (goat antirabbit HRP conjugated, 1:10,000 dilution in TBS +1% BSA TBS +0.1% Tween-20, cat. ab6721; Abcam) was applied for 1 h at RT under agitation (100 rpm on an orbital shaker).
Total protein content and proteins of interest were imaged using the Gel Doc EZ Imager and the ChemiDoc™ Gel Imaging System (using Clarity™ Western ECL Substrate System; Bio-Rad), respectively. Analysis was performed using Image Lab Software (Bio-Rad), and densitometry measurements for the proteins of interest were first normalized to total protein densitometry, followed by normalization between blots using samples as loading controls.
Statistical analysis
Statistical tests were performed using GraphPad Prism 6.0 software (GraphPad Software, San Diego) and are detailed in the individual figure legends. Differences were considered statistically significant at p < 0.05. Implants were analyzed from six mice per time point (n = 6 mice, n = 6 ASC-seeded, 6 unseeded DAT implants/time point); two outliers were identified using Grubb's test for statistical outliers and removed from the following datasets: one data point in the cell infiltration dataset (ASC-seeded scaffold group, 3-week time point) and one data point in the adipogenesis quantification dataset (unseeded scaffold group, 8-week time point).
Parametric tests were used for all analyses except for the correlation analyses and the adipose tissue remodeling dataset. For correlation analyses, the Shapiro-Wilk normality test reported significant results (p < 0.001), suggesting that the data were not normally distributed; thus, Spearman's correlation analysis was selected as a nonparametric test. To analyze the adipose tissue remodeling dataset, the Scheirer–Ray–Hare test 34 (computed using the “rcompanion” package 35 in R statistics software 36 ) was selected as a nonparametric test given that visual inspection of the data showed a violation of normality. Scaffold seeding and time postimplantation were assigned as the main effects in the model.
Results
ASCs enhance cell recruitment and tissue remodeling in DAT scaffolds
After implantation, ASC-seeded and unseeded DAT scaffolds were well tolerated; the incision sites healed within the course of the first week, and there were no detectable signs of inflammation or irritation in any of the animals. Both ASC-seeded and unseeded DAT scaffolds were recoverable over the duration of the study (Fig. 2 and Supplementary Figs. S4 and S5). In Masson's trichrome-stained sections, a thin fibrous capsule was visualized along the scaffold periphery starting at the 1-week time point and persisting over the duration of the study. For analysis, all blood vessels and adipocytes contained within this fibrous capsule were considered newly formed tissues. Macroscopically, the implants retained their volume over the course of the 8-week study (Fig. 2 and Supplementary Figs. S4 and S5).
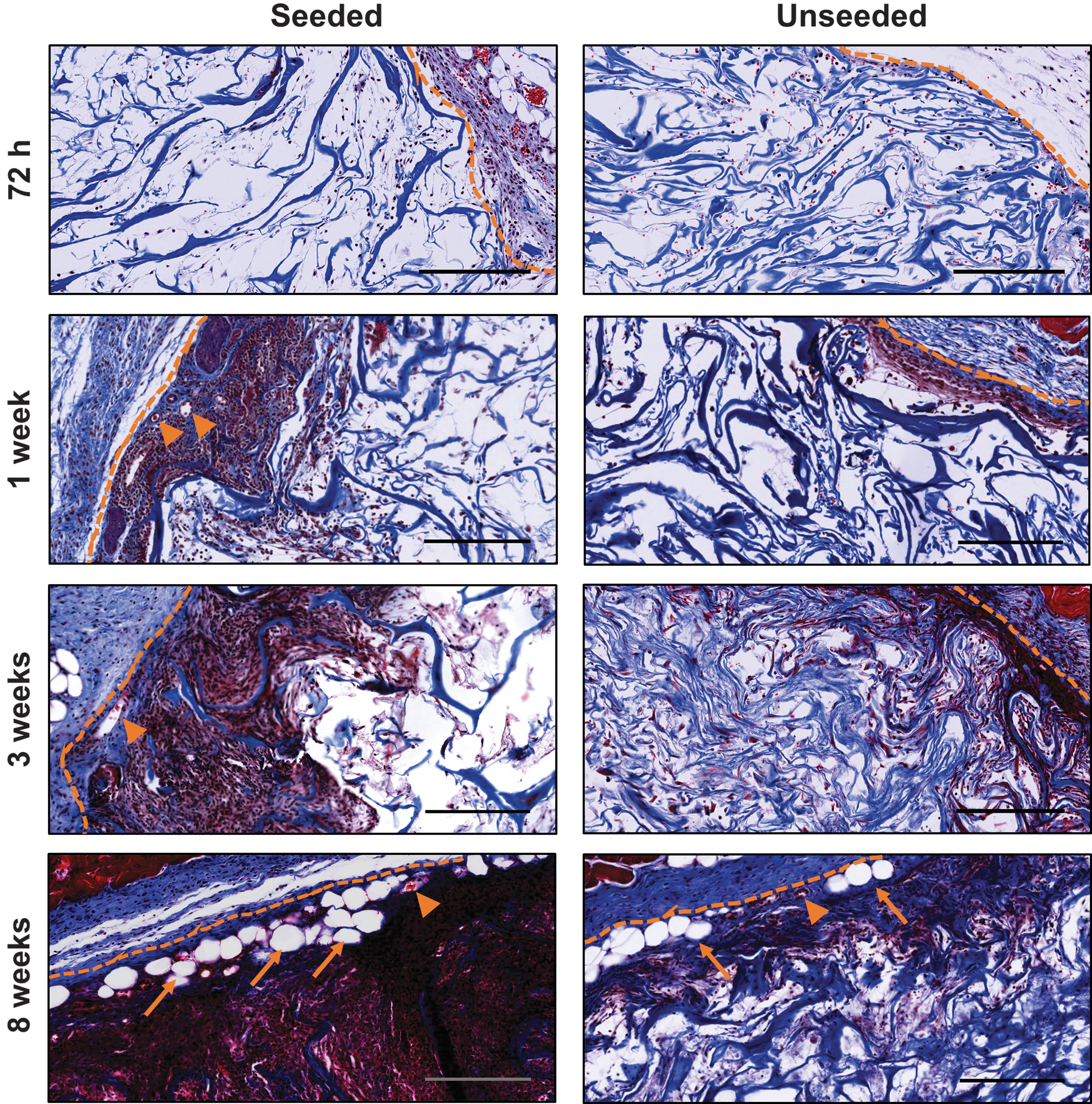
Cell recruitment, angiogenesis, and adipogenesis were enhanced in ASC-seeded DAT scaffolds relative to unseeded controls. Representative Masson's trichrome-stained sections of ASC-seeded and unseeded DAT implants at each time point are shown. Dashed line: scaffold boundary; arrow heads: erythrocyte-containing blood vessels; arrows: adipocytes. Scale: 200 μm. DAT, decellularized adipose tissue.
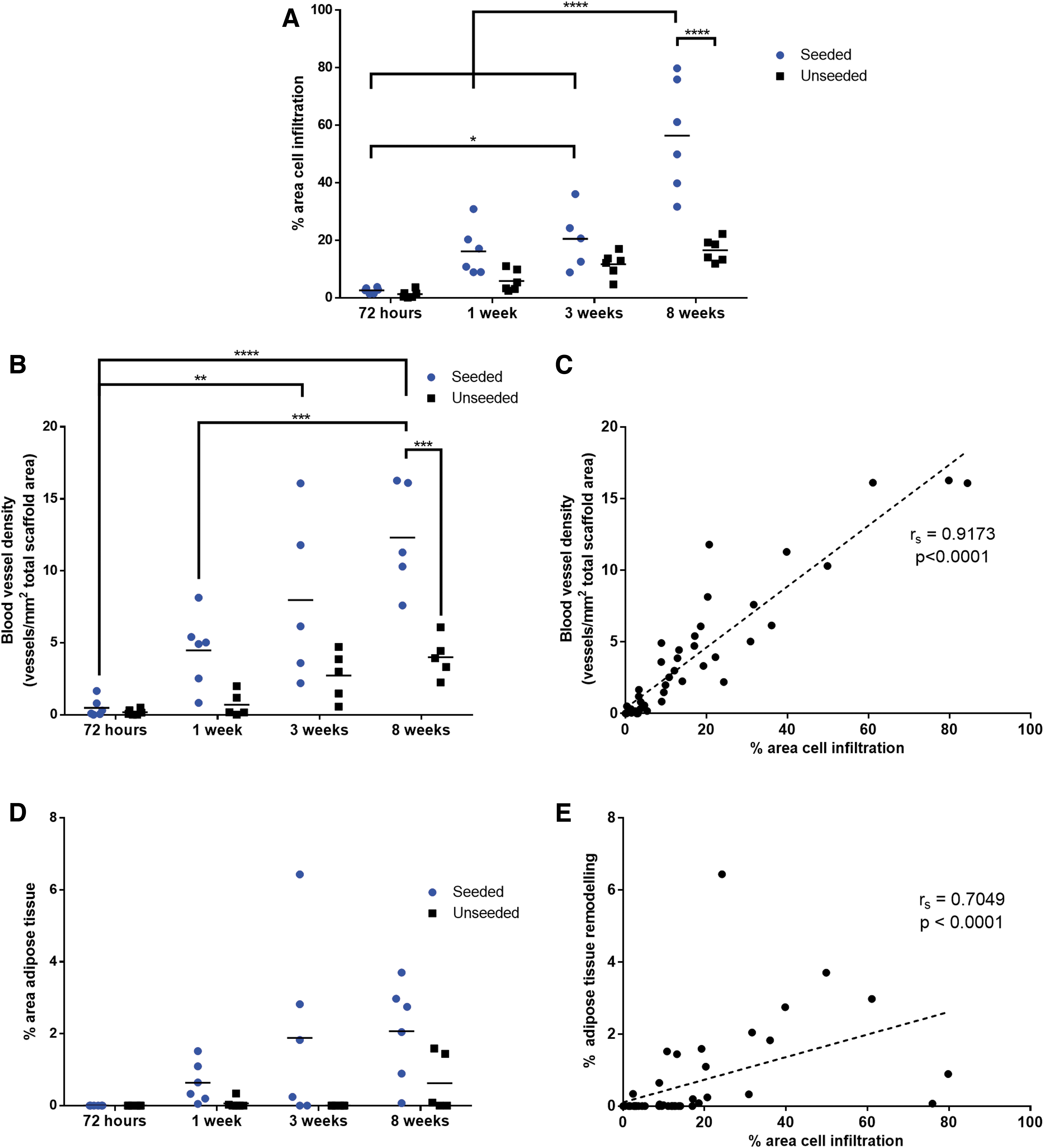
Quantitative image analysis revealed that the scaffold area infiltrated by cells was significantly increased at 8 weeks in the ASC-seeded DAT relative to the unseeded DAT, indicating that the syngeneic ASCs promoted cell recruitment within the scaffold region (Fig. 3A). In addition, a significant difference was observed over time in the seeded group, with higher infiltration at 8 weeks relative to all other time points. In contrast, no significant changes in the cell infiltration area occurred within the unseeded scaffolds over time.
ASC seeding augments cell recruitment, angiogenesis, and adipogenesis within the DAT scaffolds in the MacGreen mouse model.
Quantification of erythrocyte-containing blood vessels within the Masson's trichrome-stained sections demonstrated statistically significant increases in the blood vessel density across time points within the ASC-seeded scaffolds, with a significantly greater blood vessel density in the ASC-seeded versus unseeded scaffolds at 8 weeks postimplantation (Fig. 3B). No significant differences were observed in the unseeded DAT scaffolds over time. Correlation analysis revealed a strong and significant positive correlation between cell infiltration area and blood vessel density (Fig. 3C).
Assessment of adipose tissue remodeling demonstrated a modest increase in the percentage of the scaffold area that had remodeled into adipose tissue within the ASC-seeded scaffolds (Fig. 3D). Within the unseeded DAT scaffolds, adipose tissue remodeling was observed within only a small number of the implants. Simple main effects analysis revealed that the extent of adipose tissue remodeling of the DAT was significantly increased by ASC seeding and with extended time. Furthermore, a significant positive correlation between cell infiltration area and adipose tissue remodeling was observed (Fig. 3E).
Donor ASCs are retained in DAT scaffolds but do not contribute directly to newly formed adipose tissue
To assess syngeneic donor dsRed+ ASC retention within the DAT scaffolds, immunohistochemical staining for dsRed was performed on ASC-seeded implants and unseeded controls. These donor dsRed+ ASCs expressed the defining ASC surface markers and displayed robust in vitro differentiation capacity toward the adipogenic lineage (Supplementary Fig. S1). Positive dsRed staining was detected within the ASC-seeded implants at all time points examined (Fig. 4A and Supplementary Fig. S6), and quantification revealed no significant changes in staining levels across the time points examined (Fig. 4B), suggesting that the donor cells were retained over the course of the study. DsRed staining did not appear in association with newly formed adipocytes within the scaffold region, indicating that these tissues were host derived. As expected, no positive staining for dsRed was observed within the unseeded DAT scaffolds (Fig. 4A).
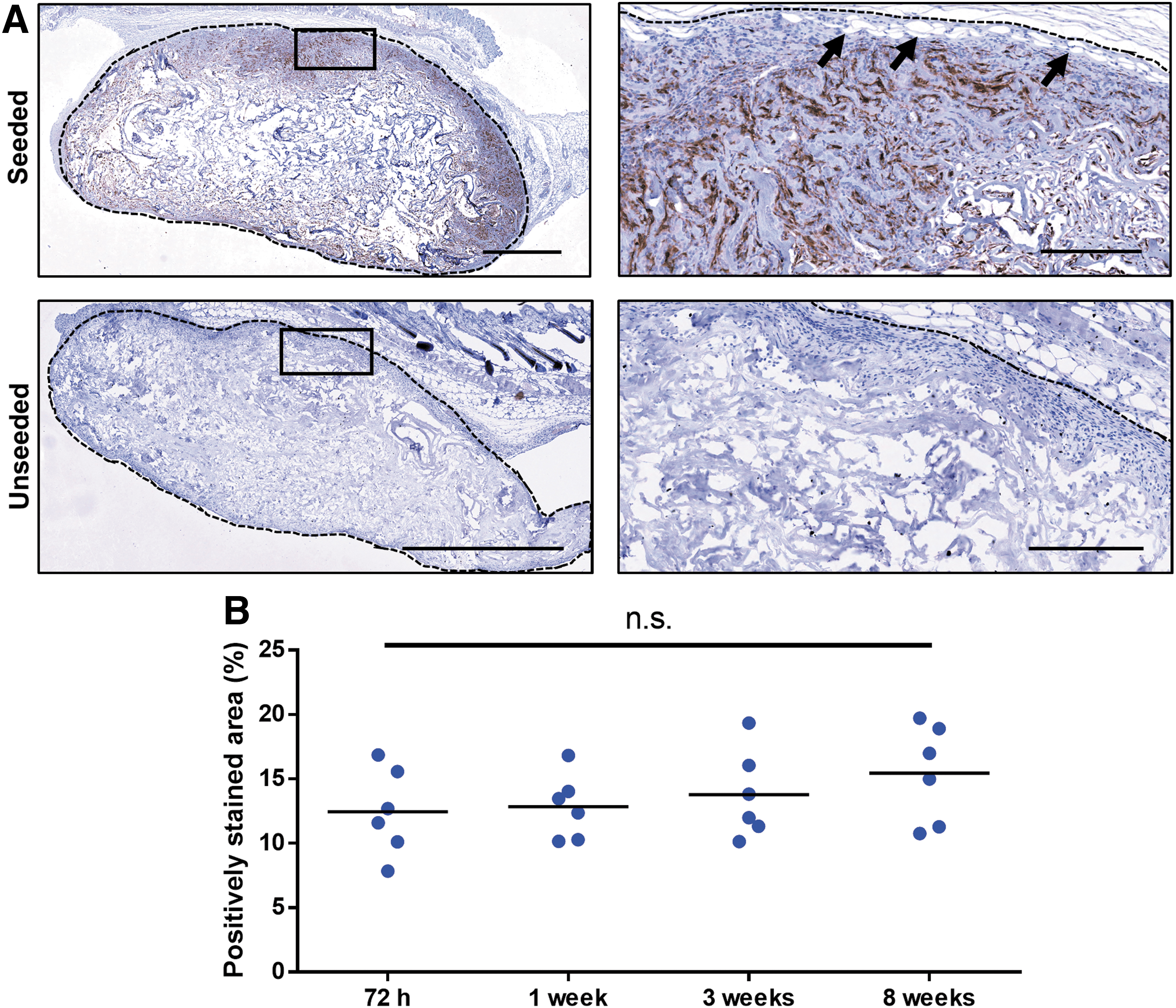
Syngeneic dsRed+ donor cells were retained in the ASC-seeded DAT scaffolds over 8 weeks and did not contribute to the newly formed adipocytes within the implants.
ASCs modulate macrophage recruitment and phenotype in DAT scaffolds
To investigate macrophage infiltration into the DAT implants, costaining was performed for EGFP, expressed by myeloid cells within the MacGreen mouse model, 27 and the macrophage marker Iba1.37,38 EGFP+Iba1+Hoechst+ cells were observed in both seeded and unseeded implants at all time points examined, with peak levels observed at 3 weeks postimplantation in both groups (Fig. 5 and Supplementary Figs. S7 and S8).
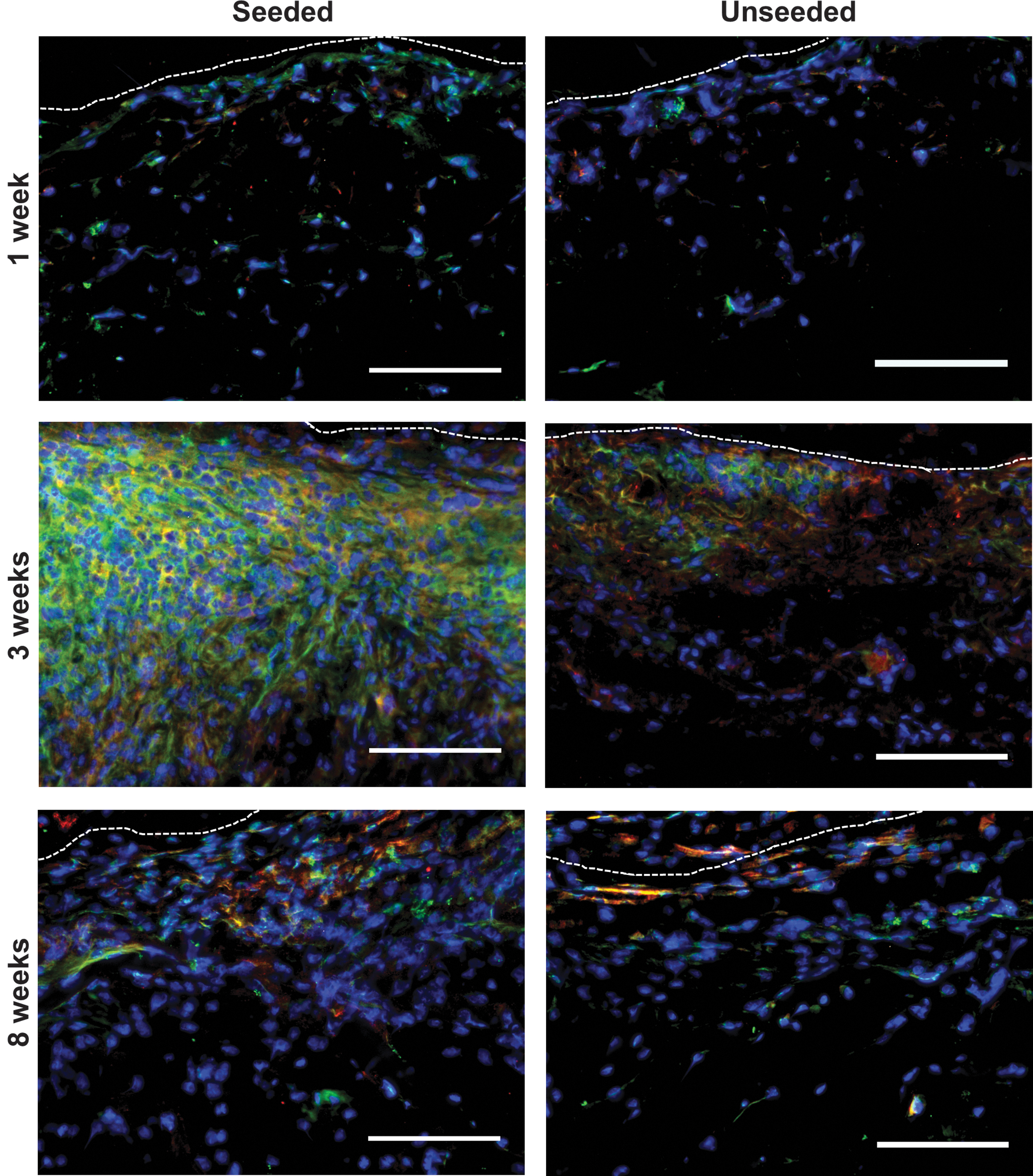
Macrophage infiltration peaked at 3 weeks postimplantation in ASC-seeded and unseeded DAT scaffolds. EGFP+Iba1+Hoechst+ cells were observed at 1, 3, and 8 weeks postimplantation, with peak infiltration at 3 weeks. Dashed line: scaffold boundary. Blue: cell nuclei; green: EGFP; red: Iba1. Scale: 100 μm. EGFP, enhanced green fluorescent protein.
Quantitative analysis of EGFP and Iba1 costaining was performed using images captured on the implant periphery (within 500 μm of the implant boundary), which represented the major region in which macrophage infiltration was observed across all groups. Analysis of macrophages (EGFP+Iba1+Hoechst+ cells) revealed a significant peak in cell numbers at 3 weeks compared with the 1-week time point, and this macrophage influx was significantly greater in the ASC-seeded group relative to the unseeded scaffolds (Fig. 6A). At 8 weeks postimplantation, macrophage numbers were significantly reduced relative to 3 weeks in the ASC-seeded groups.
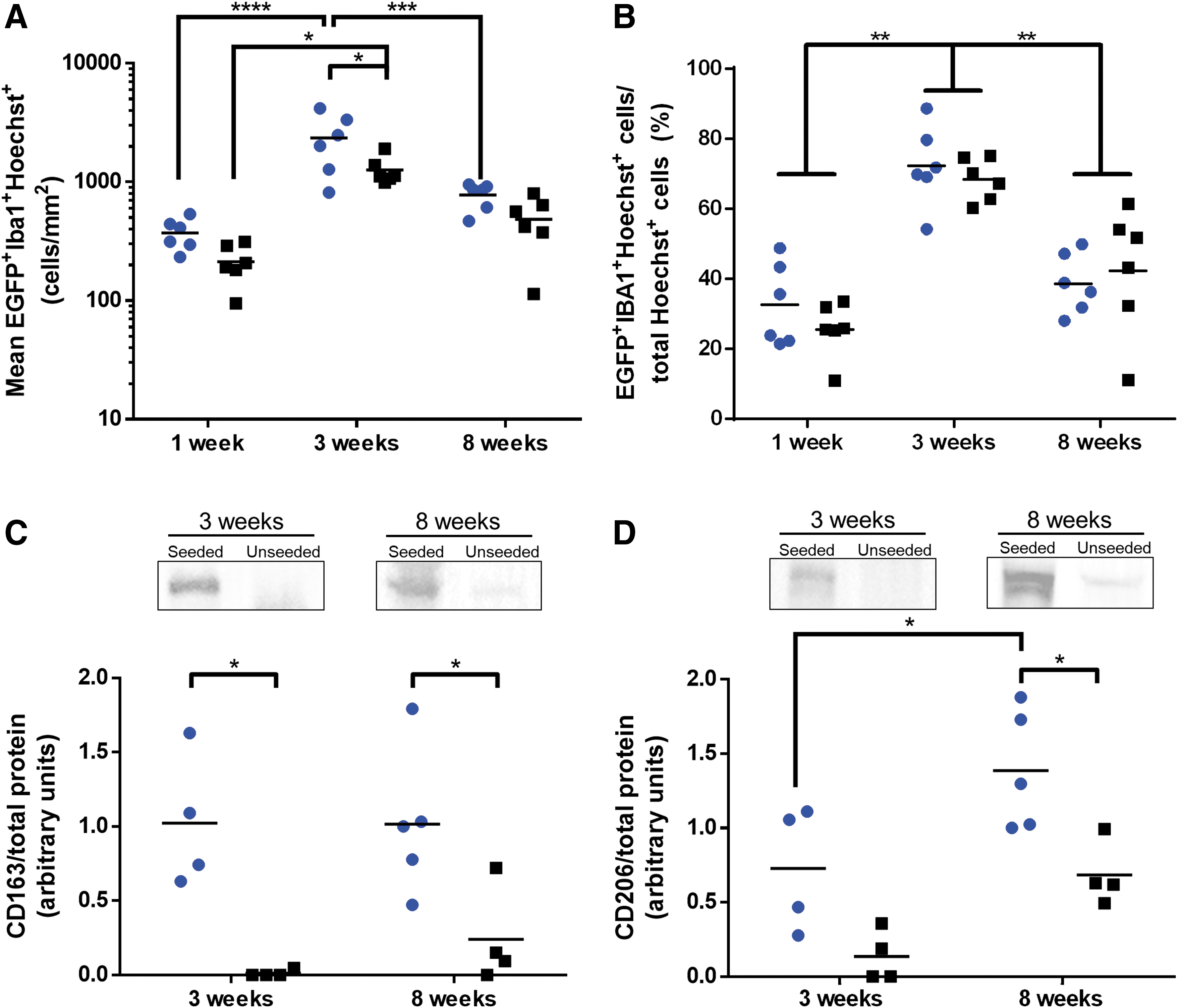
ASC seeding enhanced the macrophage density within the DAT scaffolds at 3 weeks and increased the expression of markers associated with proregenerative M2-like macrophages.
To account for differences in cell density observed along the implant periphery, macrophage numbers were normalized to the total number of Hoechst+ cells counted within each region (Fig. 6B). Similar fractions of macrophages were observed within ASC-seeded versus unseeded implants at each time point examined. Notably, the relative macrophage fraction represented >60% of all cells at the 3-week time point, and this fraction was significantly greater at 3 weeks relative to the 1- and 8-week time points.
To investigate macrophage phenotype, quantitative western blotting was performed to examine expression of the scavenger receptors CD163 and CD206, which are associated with a proregenerative M2-like macrophage phenotype. 39 The 3- and 8-week time points were selected for this analysis since increased blood vessel and adipose tissue formation were observed at these time points. Our data revealed significantly enhanced expression of CD163 in the ASC-seeded versus unseeded DAT scaffolds at the 3- and 8-week time points (Fig. 6C and Supplementary Fig. S9). In addition, there was significantly greater CD206 expression in the ASC-seeded DAT as compared with the unseeded controls at the 8-week time point, with a significant increase in the seeded group from 3 to 8 weeks (Fig. 6D and Supplementary Fig. S9).
Discussion
The combined use of decellularized bioscaffolds with proregenerative MSC populations has shown promise for a range of tissue-engineering applications. In the context of adipose tissue engineering, DAT scaffolds offer a tissue-specific platform for regeneration that can be enhanced through the addition of ASCs.18,23,26 However, greater mechanistic understanding of ASC-mediated regeneration is needed to facilitate the design of improved strategies to promote stable and predictable soft tissue regeneration.
Previous studies have shown that donor ASCs improved angiogenesis and adipogenesis in ECM-derived scaffolds, and suggested that ASC paracrine mechanisms play a major role in tissue regeneration.18,23,26 To expand on this work, we employed a dual transgenic mouse system to track both implanted ASCs and host macrophages. This model enabled us to identify implanted versus host-derived cells and highlighted cell recruitment as an important regenerative mechanism that was greatly augmented by the donor ASCs. Further, correlation analysis indicated that increased cell recruitment was linked to enhanced blood vessel and adipose tissue formation within the implants.
These findings suggest that the increased cell infiltration was not a maladaptive immune reaction against the donor ASCs, but rather supported tissue remodeling. ASCs are known to release a variety of chemotactic factors linked to the recruitment of cells, including leukocytes and endogenous progenitors,40,41 thereby representing a possible mechanism by which the donor ASCs mediated these effects. In alignment with our findings, several groups have investigated the loading of ECM-derived scaffolds with chemotactic factors such as stromal cell-derived factor-1, reporting augmented cell recruitment and tissue remodeling relative to scaffolds alone.42,43
In addition to enhanced cell recruitment, neovascularization was significantly augmented in ASC-seeded DAT relative to unseeded scaffolds. Angiogenesis is critical for soft tissue engineering since mature adipose tissue is densely vascularized. Moreover, angiogenesis is tightly coupled to adipogenesis in development and in obesity. 44 Using a tissue-engineering chamber incorporating ECM, Ting et al. 45 demonstrated that the addition of the proangiogenic factors VEGF-A, FGF2, and PDGF-BB significantly increased both angiogenesis and adipose tissue formation in the chamber relative to nongrowth factor-loaded controls, further supporting the important role of neovascularization in adipogenesis.
Immunohistochemical analysis of donor dsRed+ ASC retention revealed that the syngeneic ASCs were retained over the course of the 8-week study. Assessment of donor cell retention is valuable in cell-based regenerative approaches to evaluate the efficacy of the cell delivery platform, as well as immune tolerance to implanted cells, and the mechanisms of tissue regeneration. However, relatively few studies have quantitatively examined donor cell retention within decellularized scaffolds over time. Importantly, our cell tracking data showing long-term ASC retention further substantiate the use of ECM-derived scaffolds to provide a cell-supportive niche for transplanted cells.1,2
In addition to the scaffold delivery platform, the genetic inter-relationship between donor MSCs and the host may be another important factor influencing long-term donor cell retention. While the immunosuppressive properties of MSCs have facilitated their use in allogeneic contexts, cell-mediated and humoral immune responses have been reported using MHC-mismatched donor MSCs. 46 Within immunocompetent animal models, several authors have reported retention of autologous or syngeneic MSCs within decellularized scaffolds over a period of ≥8 weeks.47–49 In contrast, studies using allogeneic sources have typically shown retention of donor cells only within 4 weeks postimplantation.26,50 Allogeneic MSC sources are attractive from a translational perspective as an off-the-shelf product and have shown promise in a variety of tissue-engineering contexts. However, regardless of their source, the question of whether prolonged retention of donor MSCs in scaffolds provides improved effects relative to transient retention has not yet been resolved.
Analysis of the dsRed+ donor ASCs also revealed that the newly formed adipocytes within the implants were host derived, consistent with previous reports from our group 26 and others18,51 that donor ASCs indirectly promote adipogenic remodeling in ECM-derived scaffolds. Collectively, these results suggest that paracrine signaling may be the predominant mechanism through which ASCs promote adipogenesis within a tissue-engineering context; however, additional cell tracking studies using immunocompetent animal models are required to more fully elucidate the effects over time.
As inflammation has been indicated to play a role in adipose tissue remodeling within tissue-engineered implants,24,25,52 we examined macrophage infiltration and phenotype in the ASC-seeded and unseeded DAT implants. In our model, there was a higher density of macrophages in the ASC-seeded scaffolds at 3 weeks. However, the fraction of macrophages relative to the total cell density was similar between the two groups, suggesting that the ASCs enhanced total cell infiltration, rather than specifically stimulating macrophage recruitment. Regardless, the increased presence of macrophages may have contributed to scaffold remodeling.
In our previous study in the Wistar rat model, we showed that CD68+ macrophages persisted at similar levels at 1, 4, 8, and 12 weeks in ASC-seeded versus unseeded DAT scaffolds, while the fraction of CD163+ cells was significantly higher in the ASC-seeded group at 12 weeks, correlating with enhanced angiogenesis and remodeling of the DAT into host-derived adipose tissue. 26 In contrast, this study revealed that macrophage infiltration peaked at 3 weeks postimplantation in both scaffold groups, and then declined by 8 weeks.
In parallel to the decline in the macrophage population, our data showed increasing levels of the M2-like macrophage marker CD206 at 8 weeks relative to the 3-week time point in the ASC-seeded DAT scaffolds, suggesting that the microenvironment became more proregenerative over time. In addition, CD163 expression was augmented in the ASC-seeded DAT scaffolds at both 3 and 8 weeks. However, further studies would be required to confirm whether the augmented levels were related to increased expression of CD206 and/or CD163 on the cell surface of the macrophages, or potentially to soluble receptors that can be shed from macrophages and may have persisted in the implants over time. 53
Within a tissue-engineering context, other recent studies have shown that MSCs seeded in ECM-derived scaffolds increased the abundance of CD206+ macrophages within scaffolds implanted in murine models, and also enhanced regeneration.54,55 Further, Badylak et al. have demonstrated that CD163+ and CD206+ macrophages are key determinants of tissue remodeling in ECM-derived scaffolds.56,57 Importantly, the increased expression of these markers may be linked to the angiogenic response observed within the ASC-seeded DAT, as M2-like macrophages can function in the stabilization and remodeling of nascent blood vessels.58,59
Conclusion
Taken together, our results further support the combined delivery of ASCs and DAT as a strategy for soft tissue regeneration. The dual transgenic mouse system provided a useful model to track and characterize donor ASCs and macrophages to investigate their roles in tissue remodeling. Notably, the donor ASCs persisted within the DAT implants over the duration of the study. The results demonstrate that ASC seeding promoted the recruitment of beneficial host cell populations, as well as the expression of M2-like macrophage markers, and was associated with increased angiogenesis and adipogenesis within the DAT. Overall, this study provides valuable preclinical data supporting the use of ASC-seeded DAT scaffolds for future clinical applications in soft tissue augmentation and reconstruction, along with further evidence of the proregenerative immunomodulatory capacity of ASCs delivered within tissue-engineered bioscaffolds.
Footnotes
Acknowledgments
The authors thank Cody Brown, Gagandeep Singh, and Hisham Kamoun for their technical assistance; Dr. Frank Beier for providing access to his Gel Doc imaging system; and Drs. Aaron Grant, Brian Evans, and Robert Richards for their clinical collaborations to support this work.
Disclosure Statement
No competing financial interests exist.
Funding Information
Funding for this work was provided by the Canadian Institutes of Health Research (CIHR) (FRN 119394).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.