
Abstract
The tenocyte niche contains biochemical and biophysical signals that are needed for tendon homeostasis. The tenocyte phenotype is correlated with cell shape in vivo and in vitro, and shape-modifying cues are needed for tenocyte phenotypical maintenance. Indeed, cell shape changes from elongated to spread when cultured on a flat surface, and rat tenocytes lose the expression of phenotypical markers throughout five passages. We hypothesized that tendon gene expression can be preserved by culturing cells in the native tendon shape. To this end, we reproduced the tendon topographical landscape into tissue culture polystyrene, using imprinting technology. We confirmed that the imprints forced the cells into a more elongated shape, which correlated with the level of Scleraxis expression. When we cultured the tenocytes for 7 days on flat surfaces and tendon imprints, we observed a decline in tenogenic marker expression on flat but not on imprints. This research demonstrates that native tendon topography is an important factor contributing to the tenocyte phenotype. Tendon imprints therefore provide a powerful platform to explore the effect of instructive cues originating from native tendon topography on guiding cell shape, phenotype, and function of tendon-related cells.
Impact statement
Tendon-mimetic surface topographies replicated from in vivo healthy tendons enable studying tenocytes in vitro. Tenocyte cell shape was found to be a dominant factor in determining its phenotype, even in nonphysiological conditions. The dominant effect of topography on cell shape and overall phenotype can be exploited to search for treatment strategies in tendinopathy, generally characterized by a loss of cell shape.
Introduction
Tendon is a unique type of connective tissue that transmits muscle contraction forces to bones to produce motion and maintain body posture. 1 In healthy tendon, a typical hierarchical arrangement of parallel collagen fibrils and fibers forms a tendon unit, which in an unloaded state adopts a crimp-type/wavy configuration. 2 Tendon fibroblasts, that is, tenocytes, contribute to tissue homeostasis when exposed to this highly ordered collagen extracellular matrix (ECM) by producing ECM proteins and thus collagen assembly and turnover.3,4 However, due to its low cellularity and hypovascularity, tendons possess a low regenerative capacity that results in poor and slow healing.5–7 Injury or tissue damage often results in an increased ratio of type III collagen to type I collagen and additional deposition of glycosaminoglycans. 1 From a structural point of view, the organization of collagen fibers change from highly anisotropic to more isotropic and at the microlevel, they become more angulated and the number of small-diameter collagen fibers is increased. 8 At the cellular level, tenocytes become more stellate shaped 1,8–10 and the expression of chondrogenic genes such as Col2a1 and Aggrecan increases,9,11 whereas the expression of tendon-related markers, that is, Tenomodulin (Tnmd) 12 and Scleraxis (Scx) decreases. 13
Similar to in vivo, changes in tenocyte phenotype, shape, and function are observed during in vitro culture of tendon-derived cells on flat tissue culture plastic, which thus lacks the tendon ECM niche. On flat tissue culture polystyrene (PS), tenocytes quickly lose their elongated shape and decrease the expression levels of Scx. 14 Yao et al., reported that the amount of collagen type I (COL1) and decorin (DCN) protein expression levels decreased significantly from passage 0 to 8 during in vitro culture. 15 Mazzocca et al., showed that in addition to Scx, Dcn, and Col1, gene expression levels of other tenocyte markers, that is, Tenascin-C (Tn-C) and Tnmd, decline from passage 0 to 6. 16 This phenomenon is referred to as dedifferentiation, and is also a well-known phenomenon observed in chondrocytes. 17 These results point toward the lack of a healthy tendon niche, that is, biochemical and topographical changes, driving this loss of tenocyte phenotype, in analogy to changes in tissue organization in vivo that similarly affect tenogenicity.
Various strategies were used to redifferentiate the dedifferentiated tenocytes to rescue their tenocyte phenotype in vitro. Supplementing culture medium with various growth factors, such as VEGF, 18 IGF, 19 and GDF5, 20 increases expression of tenocyte marker genes, including Scx and Tnmd. Additionally, serum deprivation, that is, depletion of medium-associated growth factors rescued the lost tenocyte phenotype. 21 In addition, the small molecule tazarotene, which targets the retinoic acid receptor, and preserves tenocyte phenotype through Scleraxis. 22
Manipulating tenocyte shape has also been used as an approach to redifferentiate dedifferentiated tenocytes as well as induce tenogenic differentiation of stem cells, and one such approach that pushes cells into an elongated shape is by exposing cells to a topographical cue, that is, a substrate composed of anisotropic fibers23–32 or microgrooves.26,33 For instance, Kishore et al., used aligned collagen threads to mimic the packing density, alignment, and strength of native tendon, and observed an increase in gene expression of SCX, TNMD, and TN-C compared with the randomly organized fibers in human mesenchymal stem cells. 27 Similarly, Younesi et al., fabricated three-dimensional (3D) bio textiles from collagen and observed that compared with random fibers, expression of Col1a1 and Tnmd increased more than 6- and 11-fold, respectively, in human mesenchymal stem cells. 24 Tu et al., promoted tenogenic differentiation of mouse mesenchymal stem cells in vitro and tendon regeneration in vivo by aligned electrospun fibers derived from tendon ECM. 29 An increase in the expression of tenogenic marker genes, including Scx, Dcn, and Biglycan (Bgn), was observed both in vivo and in vitro. 29
Similarly, Zhu et al., reported that morphology of pig tenocytes become elongated upon culturing on anisotropic microgrooves, compared with flat surface, and expression of TNMD elevated. 26 We previously reported that tendon-derived cells populating a reconstituted collagen tissue, ranging from uniaxially constrained with spindle-shaped cells to biaxially constrained with stellate-shaped cells, display the strongest functional remodeling capacity and tenocyte phenotype when adopting a spindle-shaped morphology. 34 In another approach, we took the cell shape concept one step further by using a microtopographical screening platform with a large amount of unique microtopographies for identifying feature characteristics associated with Scleraxis levels, and revealed a strong correlation of specific cell shapes, that is, cell and nuclear area with Scleraxis intensity in rat tenocytes. 14 These studies underline the impact of stimulating the tenocyte phenotype by manipulating the cell shape.
Based on the above, we hypothesized that the topographical cues present in the tenocyte niche contribute to phenotypic maintenance of tenocytes. We tested this by exploiting an imprinting method to replicate the native tendon topography to PS. Results of this study showed the importance of the tendon topography, pushing cells in a spindle-shaped morphology, in the maintenance of the tenocyte phenotype in vitro, which can shed a light on further understanding of tendinopathy.
Materials and Methods
Tendon tissue imprinting
Fresh porcine Achilles tendons were obtained from crossbreeds of Great Yorkshire and Dutch land pigs between 6 and 8 months of age and between 85 and 95 kg of weight, supplied by a local slaughterhouse (Compaxo Meat B.V, the Netherlands). Muscle, fat, bone-like tissues, synovial sheath, and paratenon were aseptically dissected and the remaining tendon was cut into 1 cm3 blocks and stored at −80°C. Frozen tendons were embedded in a mixture of polyvinyl alcohol and polyethene glycol (OCT; Sakura) and fixed to the cutting base plate of a cryotome (Leica CM1950) after which longitudinal sections were cut with a thickness of 300 μm. Sections were washed with phosphate-buffered saline (PBS) and stored at −80°C until use.
Tendon sections were thawed at room temperature for 30 min and placed in a six-well plate. Polydimethylsiloxane (PDMS; Dow Corning Sylgard 184, 4019862) was mixed with the curing agent at a ratio of 10:1 (w/w) and mixed vigorously. To remove the air bubbles, the mixture was centrifuged at 3000 g for 10 min and poured on tendon sections. They were allowed to cure for 48 h at room temperature on a stable flat surface. Then, the tendon section was peeled off from the PDMS resulting in a negative imprint of a tendon on PDMS. In between two glass slides, a negative imprint of PDMS was placed onto a PS film (GoodFellow) and pressed together with clamps and incubated at 140°C for 30 min. Flat surfaces were prepared by performing the same embossing method but using a flat PDMS mold. To allow cell attachment, PS surfaces were oxygen plasma treated for 45 s at 75 mTor, 50 sccm O2, and 50 W.
Isolation of rat tenocytes
Rat tenocytes were isolated from the Achilles tendon of 23-week-old Cyp1a2ren strain rats, after euthanization, due to their surplus status from the breeding program. Briefly, tendons were cut into small pieces and digested in a buffer containing 3 mg/mL collagenase type II (Worthington Biochemical), 4 mg/mL dispase II (Sigma-Aldrich), and 100 U/mL penicillin/streptomycin (Thermo Fisher Scientific) for 4 h at 37°C in a humidified tissue culture chamber with 5% CO2. Then, the suspension was passed through a 70 mm cell strainer (Life Sciences) to obtain a cell-only suspension. The cell suspension was centrifuged at 300 g for 5 min and resuspended in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS) and 100 U/mL penicillin/streptomycin. Cells were cultured in T-25 flasks until 70% confluency.
Sterilization of tendon imprints and cell culture
Sterilization of flat and tendon imprints was performed by incubating the materials in 70% ethanol for 1 h, and remaining ethanol was air dried under sterile conditions. Next, samples were incubated with sterile PBS for 30 min at 37°C and subsequently with culture medium for 30 min at 37°C before use.
Rat tenocytes were seeded on the surfaces at a density of 5000 cells/cm2 in DMEM (Sigma-Aldrich) supplemented with 10% FBS (Sigma-Aldrich) and 100 U/mL penicillin/streptomycin (Thermo Fisher Scientific). Cells were trypsinized once they reached 70% confluency. Tenocytes at passage 4 were used, unless stated otherwise.
RNA isolation and quantitative polymerase chain reaction (real time-quantitative polymerase chain reaction)
Total RNA from each sample was isolated based on the protocol described in the RNeasy Mini Kit (QIAGEN). The yield and the quality of the RNAs obtained after RNA isolation is demonstrated in Supplementary Table S1. Reverse transcription was carried out based on the protocol provided by the iScript™ Select cDNA Synthesis Kit (Bio-Rad). Quantitative polymerase chain reaction (qPCR) was performed by using iQ™SYBR® Green Supermix (Bio-Rad) by using the Bio-Rad CFX manager. Ribosomal Protein L13a (Rpl13a) was used as a housekeeping gene and relative expression was determined using the ΔΔCt method. Primer sequences are listed in Table 1.
Primer Sequences Used in This Study
Immunofluorescence staining of Scleraxis
Cells attached to flat or tendon imprint surfaces were fixed at day 1, 3, or 7 after the start of culturing with 4% paraformaldehyde (PFA; Thermo Fisher Scientific) at room temperature for 20 min and then washed with PBS (Sigma-Aldrich), twice. Next, samples were permeabilized with 0.5% (v/v) Triton X-100 in PBS for 10 min at room temperature. After permeabilization, cells were blocked with 1:100 horse serum in PBS for 1 h at room temperature. Afterward, samples were incubated in primary antibody for Scx (1:200, ab58655; Abcam) dissolved in 0.01% (v/v) Triton X-100 and 0.5% bovine serum albumin (BSA) in PBS overnight at 4°C. Next, cells were washed with 0.01% (v/v) Triton X-100 and 0.5% BSA in PBS three times and incubated with anti-rabbit secondary antibody conjugated to Alexa Fluor 647 (1:200, A27040; Thermo Fisher), together with Phalloidin–Tetramethylrhodamine B isothiocyanate (1:200; Thermo Fisher) in PBS with 0.01% (v/v) Triton X-100 and 0.5% BSA in PBS for 1 h. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI, 1:500; Sigma-Aldrich) for 1 h after washing. Finally, samples were mounted on glass cover slides with mounting medium (Dako, Agilent). Imaging was performed by using Leica DMi8 with a TIRF Multi Color microscope (Leica Microsystems CMS) with lasers at excitation wavelengths of 532 and 647 nm, for phalloidin and scleraxis, respectively.
5-Ethynyl-2′-deoxyuridine labeling
To identify the proliferating cells, the Click-iT™ 5-ethynyl-2′-deoxyuridine (EdU) Cell Proliferation Kit (Invitrogen; C10340) for Imaging (Thermo Fisher) was used based on the manufacturer's instructions. Briefly, tenocytes were serum-starved for 24 h before EdU labeling to set the biological clock of the cells equally. Samples were fixed with 4% PFA (Thermo Fisher Scientific) at room temperature for 20 min and permeabilized with 0.5% (v/v) Triton X-100 in PBS for 20 min after 24 h of incubation in 10 μM EdU solution. Afterward, cells were treated with EdU reaction cocktail for 30 min in the dark and incubated in Hoechst for another 30 min. Images were taken with a Leica DMi8 with TIRF Multi Color microscope (Leica Microsystems CMS) at 20 × magnification. The reported number of proliferating cells was reported as the number of EdU-labeled cells/total number of cells.
Atomic force microscopy and Profilometer
Tendon imprints were imaged for surface architecture by using a tapping mode atomic force microscopy (AFM, XE-100; Park Systems) by using noncontact cantilevers (PPP-NCHR; Park Systems). Data were recorded with XEP software and GWYDDION software was used to image the data. A Keyence VK-H1XM-131 at 20 × magnification was used for profilometer images.
Scanning electron microscopy
Samples were fixed with 2.5% glutaraldehyde (Fisher Scientific) at room temperature for 1 h. Then, they were washed with distilled water three times for 10 min, dehydrated in 25%, 50%, 75%, 90%, and 100% ethanol for 15 min each, and incubated in 100% ethanol for an additional 15 min. Next, samples were dried in hexamethyldisilazane (Sigma-Aldrich) for 1 h. Before imaging, samples were coated with 5 nm gold–palladium and imaged using a scanning electron microscope (SEM) (FEI Quanta 3D FEG Dual Beam).
Image analysis
An image analysis pipeline was created in CellProfiler version 3.19. To calculate expression levels of SCX and other shape parameters, 3–4 random images from three biological replicates was selected. Median values of all cells were per image calculated by CellProfiler and used to calculate the Spearman correlation between median SCX intensity and cell shape parameters. Shape parameters were calculated by taking the median value of all the cells in each single image, and subsequently depicted as a single data point in the figures. This median was derived from ∼70 cells per image in Figure 1, and ∼40 cells per image in Figures 3 and 5.
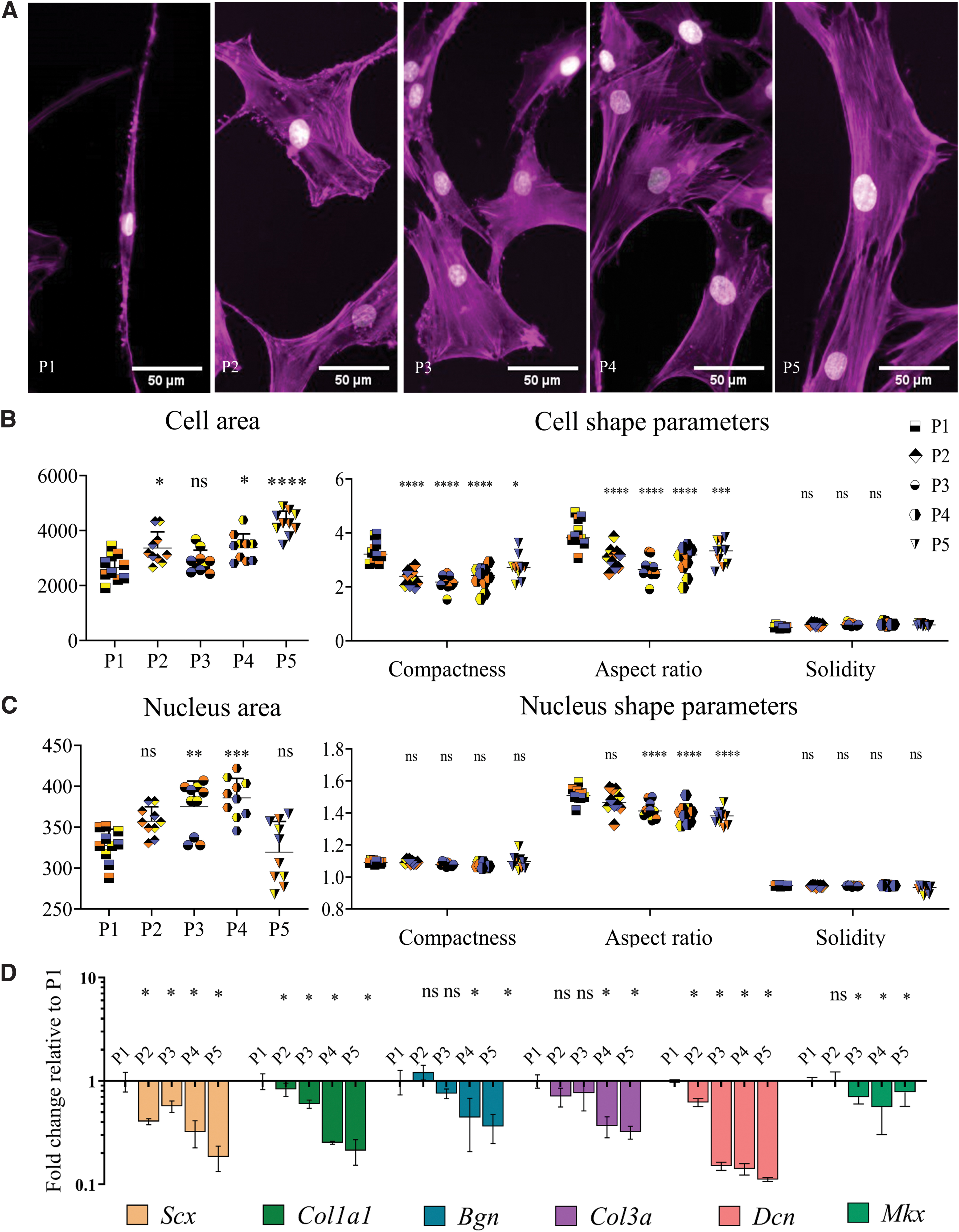
Tendon-derived cells rapidly lose their morphology and phenotype on a flat substrate.
Statistical analyses
All statistical analyses were performed by using GraphPad Prism version 8.0 (GraphPad Software, Inc., San Diego, CA). Student's t-test was performed to calculate statistical difference between cell and nuclear areas. One-way analysis of variance was carried out to calculate the statistical difference in real-time quantitative polymerase chain reaction (RT-qPCR) experiments and calculate cell/nuclear shape parameters between different passages and imprint versus flat surface. Spearman correlation coefficient was calculated by GraphPad correlation calculation option. For all statistical analyses, significance was set at p < 0.05 to determine the significance between means. All quantitative data represented in this study are based on triplicate experiments.
Results
Tendon-derived cells rapidly change morphology and phenotype on a flat substrate
Flat PS surfaces were used to assess the influence of passage number on tenocyte shape and tenocyte phenotype. Therefore, passage one (P1) rat tenocytes were cultured and subsequently passaged four times (Fig. 1). We first investigated the changes in cell and nuclear shape parameters, including area, compactness, solidity, and aspect ratio and changes in gene expression levels of tendon-related genes. Aspect ratio is calculated by dividing the length of the major axis by the length of the minor axis; so, the higher the value, the more elongated the cells. Compactness is correlated with elongation; high compactness values indicate more elongated cells. Solidity is related with cell branching and the closer its value to 1, the more solid the cells (Supplementary Fig. S1). Results showed a slight increase in cell area, that is, cell size, with passage number (Fig. 1B). Cell aspect ratio and compactness significantly decreased after P1, yet solidity remained the same in all groups (Fig. 1B). The increase in the nucleus area was statistically significant between P1 and P3, P4 but remained at similar values at P2 and P5 (Fig. 1B). Nucleus aspect ratio significantly reduced after P2, yet compactness and solidity remained unchanged between passages (Fig. 1C and Supplementary File S1 containing raw data). Overall, our data indicate that with passaging of tenocytes to P4 on flat tissue culture plastic, they undergo shape-related changes; cells and their nucleus steadily increase in size and transform from the original spindle-shaped/elongated morphology to a stellate-shaped/round morphology.
To further evaluate the loss in tenocyte phenotype, gene expression levels of Scx, Col1a1, Col3, Dcn, Mkx, and Bgn were measured (Fig. 1D). Scx, which is a tenogenic transcription factor, 35 showed a significant decrease after passage one. A similar reduction was observed for the expression of Col1a1, which is the most abundant protein in the tendon ECM, Mkx, which is a homeobox gene involved in the regulation of tendon differentiation, 36 and Col3. 37 Furthermore, we measured the expression of genes coding for noncollagenous matrix proteins Dcn and Bgn that bind to collagen, which displayed a decreasing trend from P1 to P5. 38 With these results, we confirmed that tenocytes dedifferentiate by rapidly losing their morphological characteristics and expression of tenocyte marker genes on a flat surface during in vitro subculture.
The native tendon imprint as cellular niche on PS
After establishing that with passaging, tenocytes lose their phenotype on flat PS, the effect of the tendon surface topography, as an isolated effect, was assessed. To this end, we used the soft embossing method to imprint a native tendon topography onto PS (Fig. 2). The imprinting protocol involved polymerization of PDMS using a 10:1 ratio of monomers and curing agent on a native tendon (Figs. 1–3 and 2A), and embossing the PMDS negative imprint on PS (Figs. 2A and 4–6). The imprinted material was imaged using SEM (Fig. 2B and Supplementary Video S1), AFM (Fig. 2C), and profilometry (Fig. 2D and Supplementary Video S2). SEM and profilometer images reveal that native tendon can indeed be imprinted onto PS, and the PS imprint possesses a tendon-like topography. AFM analysis shows that surface topographies on tendon imprints can reach a depth of 5 μm.
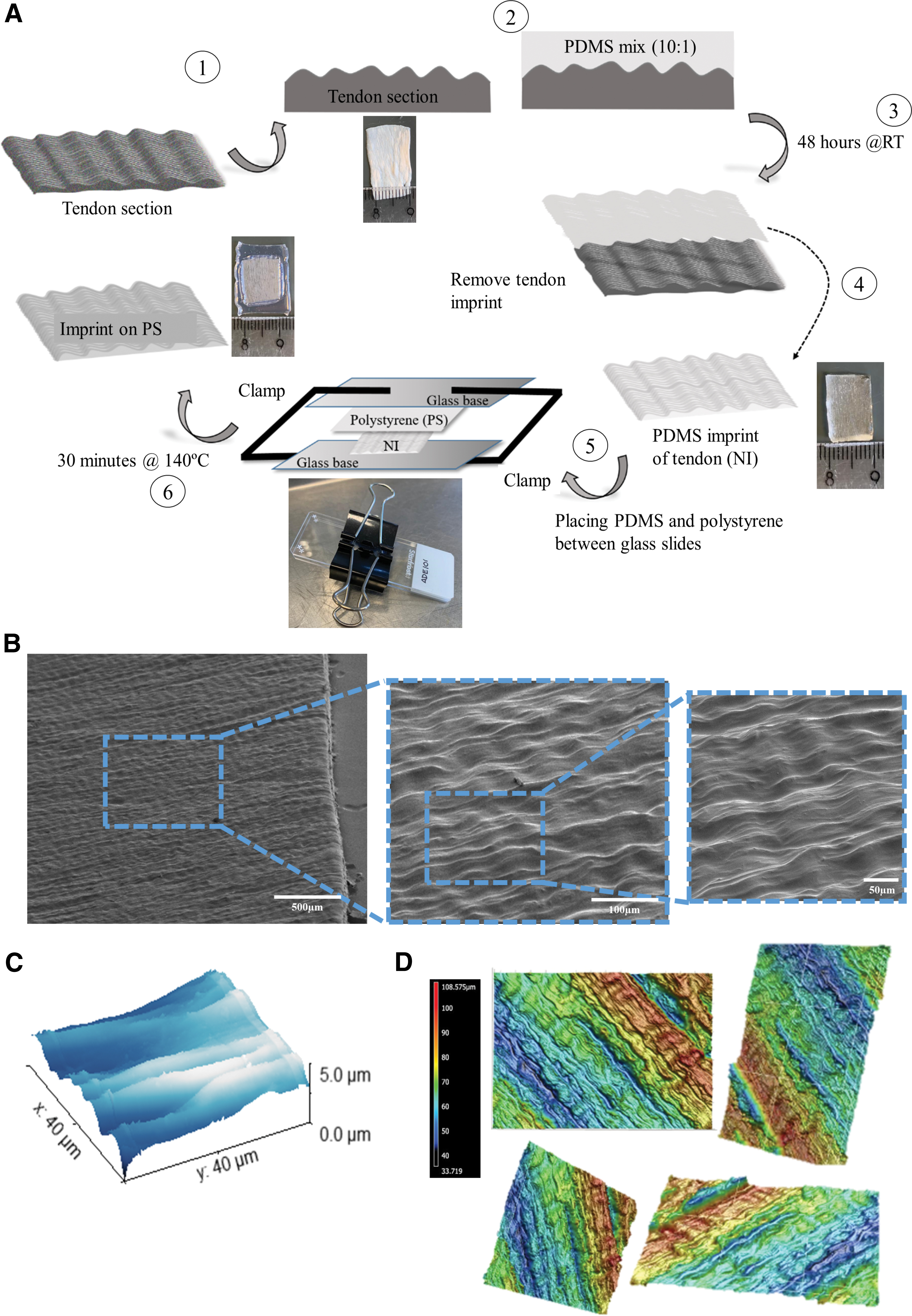
The native tendon imprinted as a cellular niche on polystyrene.
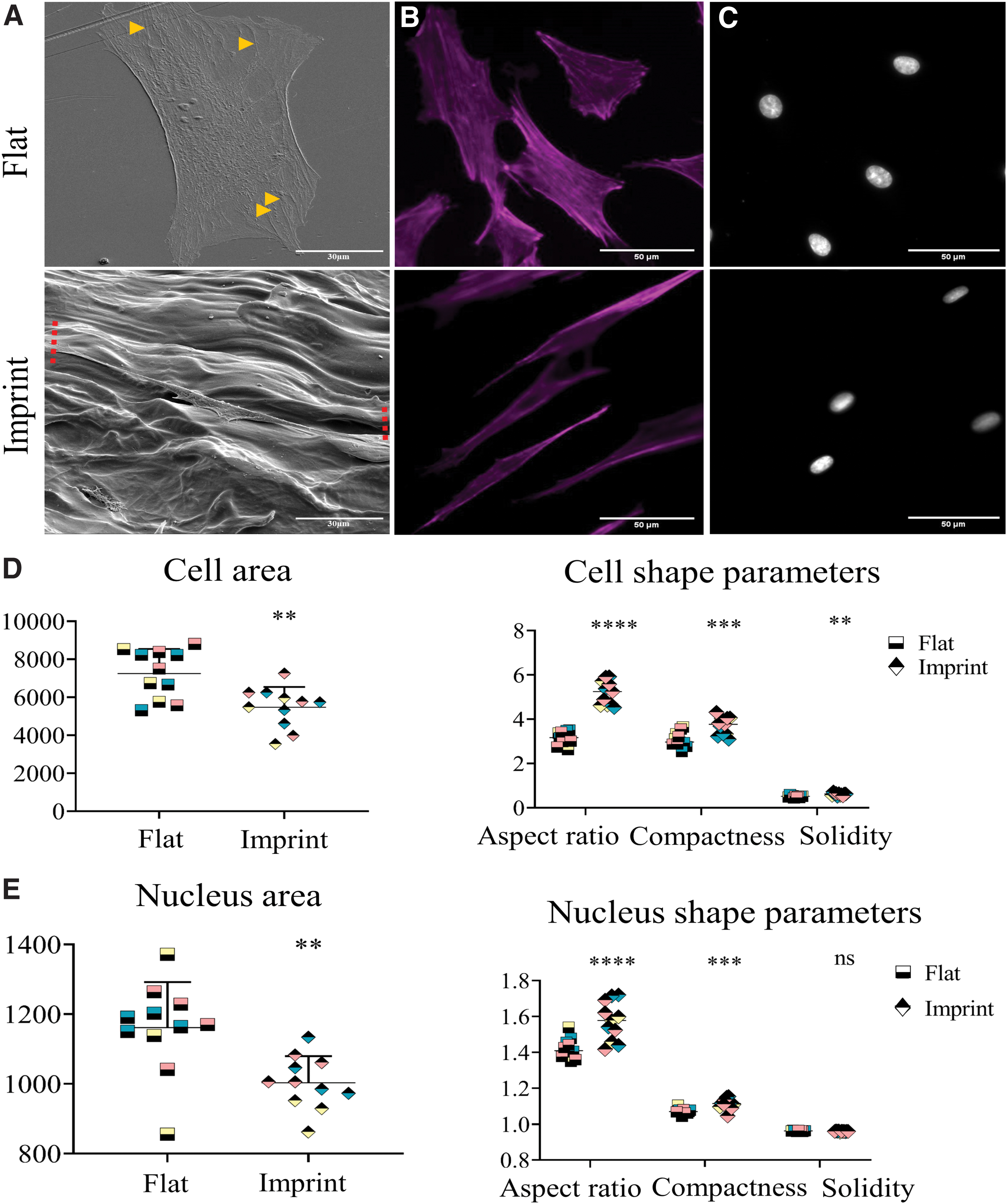
Tendon imprint topography manipulates tenocytes to their natural morphology. Tendon imprint topography pushes tenocytes to their natural morphology.
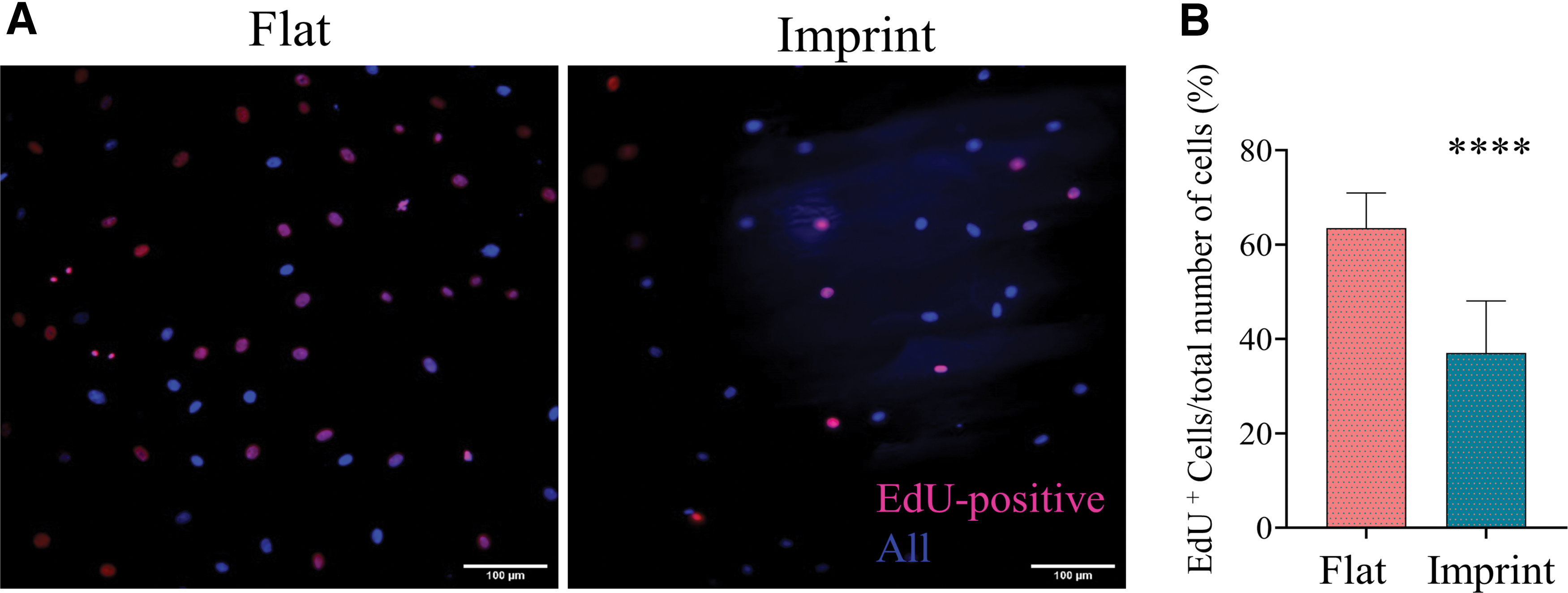
Tendon imprint topography lowers cell proliferation capacity.
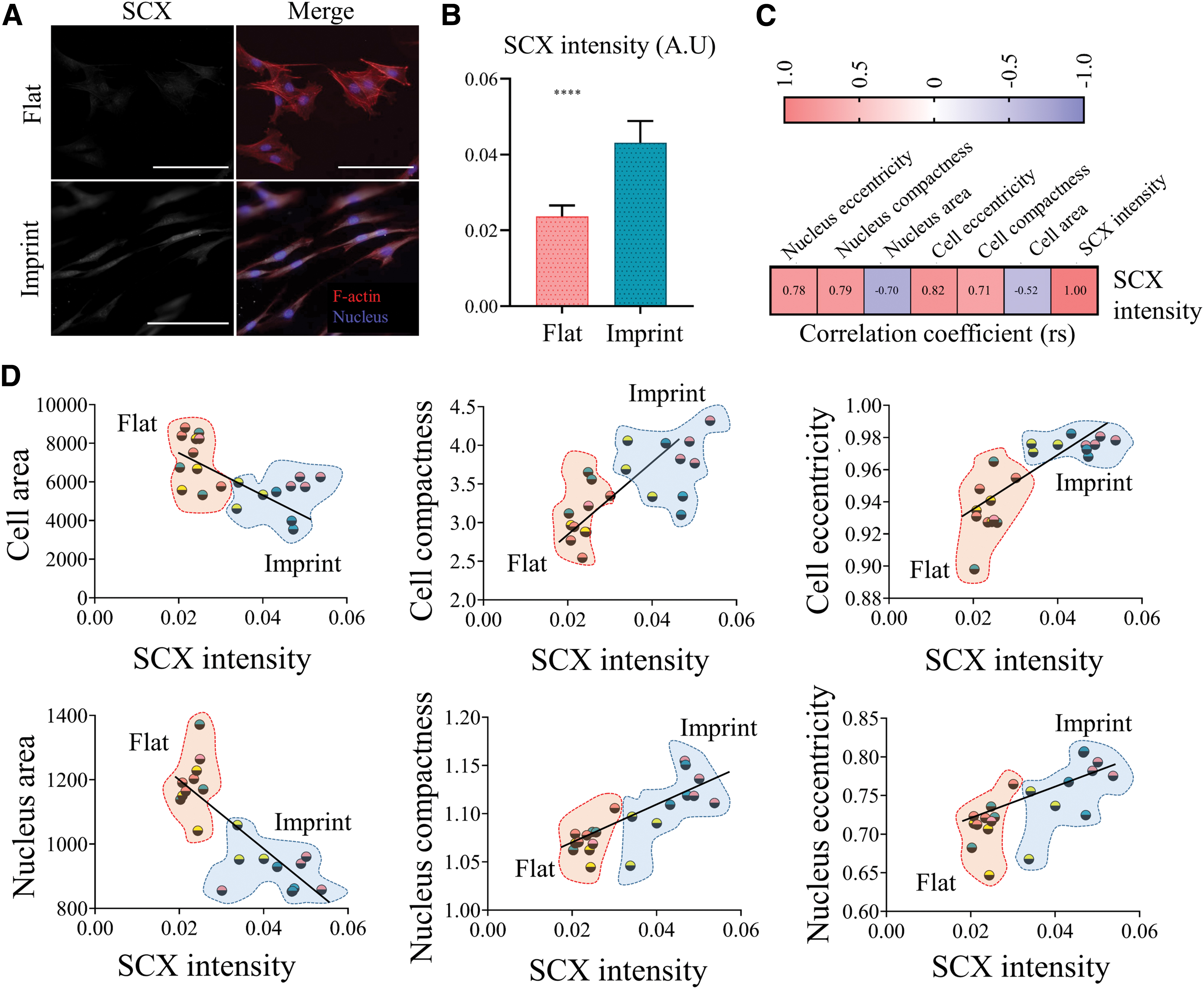
SCX intensity correlates with cellular and nuclear features. Tenocyte phenotype is maintained by tendon imprints
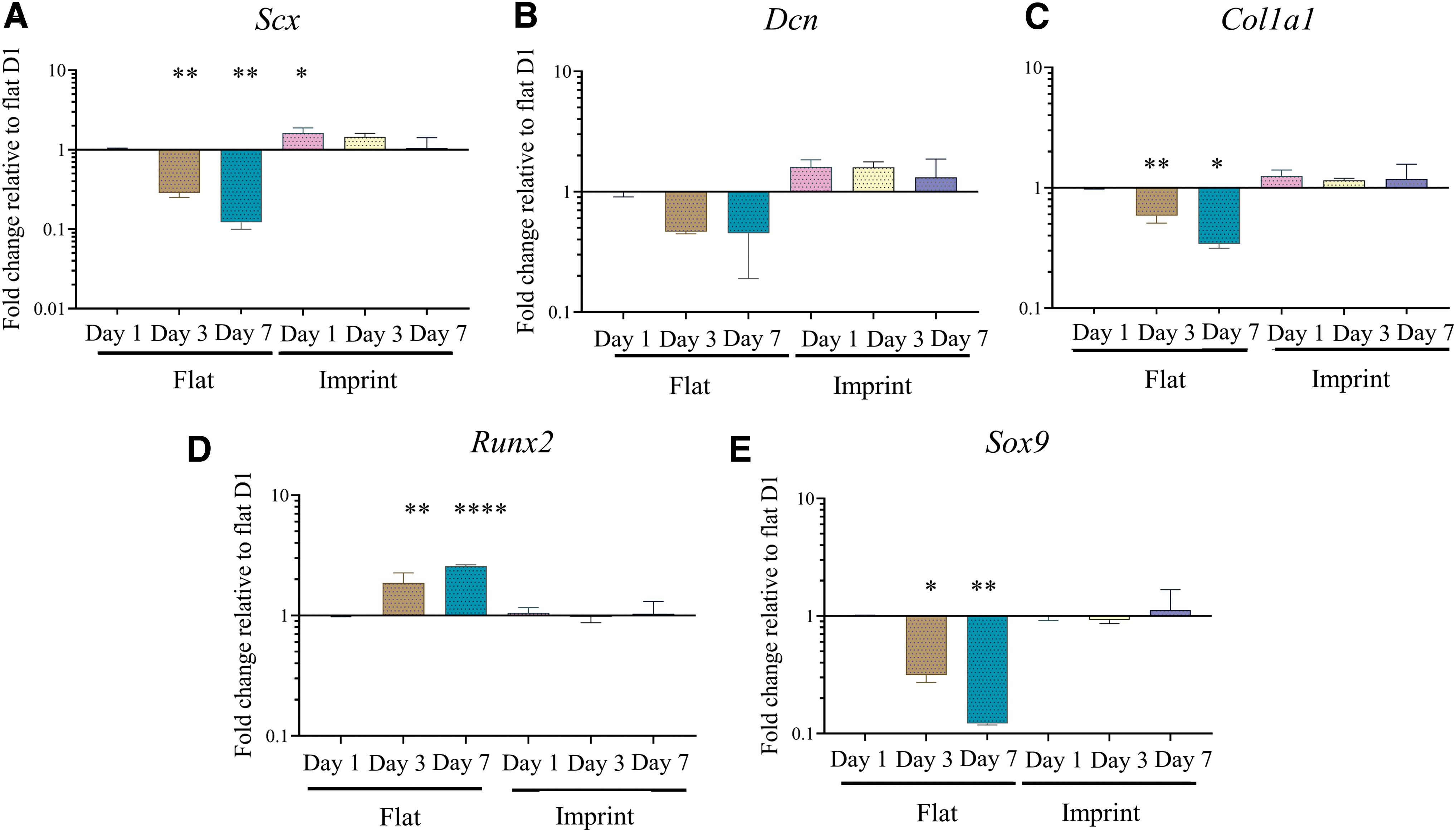
Tenocyte phenotype is maintained on tendon imprint topography. Gene expression levels of
Late-passage tenocytes adapt early passage tenocyte morphology on tendon imprints
Next, the morphological response of tenocytes to the tendon imprint was assessed (Fig. 3). Flat control surfaces were prepared by imprinting a flat PDMS mold to the PS sheets. Rat tenocytes (P4) were seeded on flat and imprint surfaces and stained with phalloidin to visualize F-actin (gray) and DAPI to visualize nuclei (blue). On the flat surface, (Fig. 3A, B, top panel) a spread morphology was observed, contrary to tendon imprints, where cells on each replicate displayed an elongated shape (Fig. 3A, B, bottom panel).
We further investigated the effect of tendon topography on cell and nuclear shape parameters, including aspect ratio, area, solidity, and compactness (Fig. 3D, E). We first visualized the difference in cell shape. Rat tenocytes were exposed to either a flat surface or tendon imprint topography and subsequently SEM imaging was performed (Fig. 3A). On the flat surface, we observed that tenocytes are more spread and stress fibers were observed (yellow arrows) (Fig. 3A, top panel). On tendon imprints, rat tenocytes were highly elongated (red arrow) (Fig. 3A, bottom panel). We observed similar results when we stained tenocytes with phalloidin to visualize the actin cytoskeleton (Fig. 3B) and DAPI to visualize nuclei (Fig. 3C). Cell and nuclear area were significantly smaller on tendon imprints compared with flat (Fig. 3E, F). Additionally, cellular and nuclear aspect ratio and compactness were significantly higher on tendon imprints, compared with flat (Fig. 3E). Cell solidity on imprints (0.61 ± 0.05) was significantly higher and closer to 1, compared with the flat surface (0.51 ± 0.04); indicating that cell branching was less pronounced on tendon imprint. Nuclear solidity was not significantly different for both topographies. Therefore, results indicate that tendon imprints induce an elongated shape and reduces cytoskeletal branching, resembling the early passage tenocytes on flat PS.
Tendon imprint topography decreases tenocyte proliferation
Surface topography has been identified as a regulator of cell proliferation.39,40 Therefore, the effect of tendon imprint topography on tenocyte proliferation compared with the flat surface was assessed. Rat tenocytes were thus cultured on a flat surface and tendon imprint, and after 24 h, the proportion of cells in the S-phase of the cell cycle (pink/purple) to all cells (blue) was measured by EdU labeling (Fig. 4A, B). On tendon imprints, 37% of the tenocytes were EdU positive, whereas on a flat surface, 63% of the cells were EdU positive. This indicates that tendon topography relates to a less proliferative tenocyte state.
SCX intensity correlates with cellular and nuclear features
Tendon imprints significantly affect tenocyte cell and nucleus shape. Next, the influence of tendon imprint topography on tenocyte marker gene expression was assessed (Fig. 5). Therefore, rat tenocytes at P4 were cultured on tendon imprints and flat surfaces for 24 h and the amount of SCX was measured by immunocytochemistry and subsequent quantification (Fig. 5A). SCX intensity were significantly higher on tendon imprint after 24 h (Fig. 5B), which was subsequently correlated with cell shape, using a Spearman correlation calculation. A positive and significant correlation was detected of SCX intensity level with cell compactness (rs = 0.71), nucleus compactness (rs = 0.71), cell eccentricity (rs = 0.82), and nucleus eccentricity (rs = 0.78) (Fig. 5C, D and Supplementary File S2 containing raw data). In addition, a negative and significant correlation was detected of SCX intensity with cell area (rs = −0.58) and nucleus area (rs = −0.70) (Fig. 5C, D). SCX level thus correlates with elongated cell and nucleus shape, driven by tendon surface topography.
Tenogenic gene expression is maintained on tendon imprints
After showing that native tendon topography can increase SCX at protein level on the short term, we investigated whether tenocyte phenotype can be maintained for 7 days of culture (Fig. 6). Therefore, P4 tenocytes were cultured on flat surfaces and imprints for 7 days and gene expression levels of Scx, Col1a1, Sox9, Dcn, and Runx2 were measured on day 1 (D1), day 3 (D3), and day 7 (D7). On the flat surface, expression of tendon-related genes Scx, Dcn, and Col1a1 decreased over 7 days of culturing, indicating ongoing dedifferentiation (Fig. 6A–C). Expression of Sox9, a chondrogenic transcription factor that is also expressed at the tendon–bone junction, also decreases on the flat surface (Fig. 6E). Lastly, Runx2 gene expression, an osteogenic-associated transcription factor, significantly increased on the flat surface. Contrary, on tendon imprints, gene expression levels of Scx, Dcn, Col1a1, and Sox9 increased at D1 and subsequently remained stable over the 7 days, indicating that tendon imprints can both induce a slight increase in tenogenic gene expression and prevent the dedifferentiation that occurs on the flat surface (Fig. 6A–C and E). Lastly, gene expression of Runx2 did not increase on the tendon imprints.
Discussion
Surface topography of various organs and tissues in nature give a series of evolutionary advantages and provide specialized functions. They provide cells with signals to differentiate, migrate, proliferate, or maintain their phenotype. The surface topography of tendon tissue is also considered to provide such strong cues. However, whether the tendon topography alone, devoid of other cues (e.g., ligand type and density, changing mechanical properties, etc.), provides a strong cellular cue remained to be explored. Therefore, the soft embossing method was exploited to imprint native tendon on a PS surface to study the effect of tendon surface topography on cell shape, proliferation, and phenotype, when compared with flat PS surfaces. Results of this study confirm that flat PS surfaces drastically alter tenogenic cell shape and phenotype, and highlight that: (1) native tendon topography can be imprinted onto a PS surface to conduct tendon-related studies in vitro, (2) cellular proliferation is decreased on imprint topographies, compared with flat surfaces, (3) tenocyte phenotype is maintained on tendon imprint topographies, and (4) remarkably, intensity levels of Scleraxis, a key tenocyte marker, beautifully correlates with specific cell and nuclear shape parameters. Concordantly, results of this study imply that imprinting the natural tendon surface on materials, such as PS, provides a tool to control tenocyte cell shape and maintenance of the tenocyte phenotype, as summarized in Figure 7.

Summary of the results. Tenocytes on imprint surfaces display a lower cell area and higher aspect ratio, compactness, and solidity, which results in increased SCX intensity. Color images are available online.
Drastic changes in cell shape of freshly isolated or early passage cells upon culturing on tissue culture plastic were shown for a wide variety of cell types, including tenocytes.15,14,41–46 Vermeulen et al. and Yao et al. showed that the elongated shape of tenocytes at P0 transforms to stellate-shaped at P1,14,15which is in agreement with the changes observed in cell and nucleus shape of the current study. Similarly, our observation on the decrease in tenogenic marker genes corroborate with previously published reports,14–16 indicative for a dedifferentiation process. Due to the rather limited availability of the early passage cells (P0 or P1), dedifferentiation has become an obstacle for in vitro research and associated interpretation of acquired data of many cell types. This gives rise to the interesting concept of redifferentiation to promote/restore the native phenotype.
The interplay between cell shape and cell fate (e.g., differentiation, proliferation, apoptosis etc.) is an established concept.17,47 Following that concept, because healthy tenocytes in vivo adopt an elongated morphology, that is, the nucleus and cell body, which are aligned between collagen fibers, redifferentiation research has focused on pushing cells toward similar morphologies. Among these approaches, cells have been exposed to anisotropic fibers24,27,31,32,48–55 or nano/microgrooves.14,26,56 For instance, Zhu et al., reported that dedifferentiated tenocytes become elongated upon culturing on microgrooves, which partially reversed the dedifferentiation process. 26 A similar study by Schoenenberger et al., showed that tendon fibroblasts displayed a higher expression of the tendon-related marker Mohawk (MKX), COL, BGN, and DCN on aligned polycaprolactone (PCL) fibers, compared with random fibers. 57
Despite their impact on tenocyte phenotype, microgrooves or anisotropic fibers may not reflect topographical cues that tenocytes or mesenchymal stem cells experience when exposed to imprint substrates as the delicate variations in roughness, ridges, and valley that create specific spatial cellular attachment sites are absent. For this, replication of native tendon topography on PDMS 39 and silica 58 have previously been performed to differentiate mesenchymal stem cells toward the tenogenic lineage, and both increased to an elevation in the expression of TNMD, compared with flat surfaces. This also suggests that tendon imprints can promote tenogenic differentiation. Results of the current study support the concept that enforcing a spindle-shaped tenocyte morphology is beneficial for maintaining/restoring their native phenotype. Specifically, our results showed that elongated cell shape, that is, high cell aspect ratio, eccentricity, and compactness, is correlated with SCX levels. However, a degree of heterogeneity in cell phenotype, that is, shape parameters and SCX levels, was observed especially for tendon imprints. Explanations for this may relate to the imprinting technique that potentially disturbs native architecture locally, or the fact that cells on imprints are exposed to a two-dimensional surface with more freedom to adopt a preferential shape, rather than the strongly confined 3D in vivo environment. Despite that, the expression of tenocyte marker genes Scx, Dcn, and Col1a1 was still shown to be stabilized on tendon imprints, contrary to a decay observed on flat surfaces.
The observation in the current study that osteogenesis (Runx2) increased on flat surfaces, compared with imprints, was also observed in another study, using osteocalcin and alkaline phosphatase as osteogenic marker genes. 31 Possibly, this relates to the increased proliferation rate on flat surfaces resulting in cell confluency, a phenomenon that was previously linked to increased Runx2 gene expression.14,24,59 Although tenocytes are the most abundant cell type in tendon tissue, depending on the location of the cells within the tendon, a Sox9-positive subpopulation of tenocytes exists that is located at the junction between bone and tendon/ligament.60,61 The expression of Sox9 was previously shown to remain stable when tenocytes were cultured, either cyclically stretched or left unstretched, on microgrooved substrates, which induces an elongated morphology. 33 Therefore, the naturally present Sox9 gene expression level in elongated tenocytes, which is stabilized on imprint surfaces, is lost on flat surfaces since the differentiation process is initiated. Suggestively, different topographical cues present in vivo in the tendon toward the enthesis, contribute to differences in phenotype of locally present cells.
Healthy tendon comprises of relatively quiescent tenocytes, whereas proliferation increases during the remodeling phase of tendon during healing, during which the strong tissue anisotropy is absent. 62 In analogy, tendon imprints resulted in the suppression of proliferation, when compared with flat surfaces. Similarly, proliferation capacity of human mesenchymal stem cells is also lower on aligned collagen fibers or anisotropic topographies, compared with randomly oriented substrates. 27 One explanation for different proliferation rates on different topographies is a difference in cell shape and alignment of stress fibers, as previously shown. 63 In elongated smooth muscle cells (SMCs), DNA synthesis was decreased, indicating that the directed cell spreading can be positively linked with DNA synthesis and thus cell proliferation. 64 A possible underlying mechanism for this could relate to NOR-1, a gene involved in SMCs proliferation. NOR-1 expression is among others modulated by protein kinase C (PKC). In elongated cells, the activity of PKC was shown to be decreased, which was associated with a decreased NOR-1 expression and thus ultimately decreased cell proliferation. 63 Considering the similarities in shape influences between SMCs and tenocytes, this could be a possible explanation.
Another, or additional, explanation is that changes in cell proliferation are linked to surface topography-induced mechanotransduction. Surface topography was shown to alter integrin activation and focal adhesion dynamics, which affect cell proliferation.65,66 Flat surfaces result in specific cell spreading and integrin binding, resulting in focal adhesion kinase phosphorylation and activation, which in turn activates the ERK pathway, cyclin D1, and finally the G1–S cell cycle transition. 67 However, activation of integrins α5 and α6 has been shown to create the opposite effect. 68 This indicates that the exact mechanism by which tenocyte proliferation is coupled to cell shape, surface topography, mechanotransduction pathways, and specific integrin recruitment on tendon imprints should further be explored.
The use of tendon imprints in research has several advantages. First, the production of tendon imprints requires only PDMS, PS, and tools to perform soft embossing; therefore, upscaling the production can be cheap and fast. Second, they bear the potential to be used as a model system to study the molecular mechanism behind the dedifferentiation and redifferentiation process, including the role of surface topographies on the activation of mechanotransduction pathways. Third, the imprinting technology is facile enough that it can be applied to any polymer that does not degrade above 140°C. Therefore, in addition to the influence of healthy tendon surface topography, the effect of other polymers on tenocyte phenotype can be investigated. Finally, the technique also allows imprinting the pathogenic or damaged tendon topography on PS; therefore, the role of altered surface topography on tenocyte phenotype can be further explored in vitro.
Conclusion
This study shows that tenocyte cell shape and expression of tenogenic marker genes drastically change during in vitro subculture, which leads to decreased tenogenic characteristics. Furthermore, we demonstrated that tendon imprints, which carry the topological cues from the native tendon, led to an elongated cell shape and resulted in maintenance in the expression of tenogenic-associated marker genes, which is positively correlated with elongated cell and nuclear shape parameters, that is, aspect ratio, compactness, and eccentricity. Overall, results of this study support the concept of cell shape to phenotype in tenocytes and stresses the role of surface topography on tenocyte phenotype.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.