
Abstract
Although mesenchymal stem cells (MSCs) transplantation is reportedly a promising strategy for repairing damaged articular cartilage, MSCs-based cartilage tissue engineering has numerous limitations, including poor implanted cell adhesion, phenotypic alteration of cells, regulation of mechanical properties, and engraftment rates after implantation. This study aimed to investigate the efficacy of transplantation of synovium-derived mesenchymal stem cells (SDSCs) encapsulated in a hyaluronic acid/collagen/fibrinogen (HA/COL/FG) composite gel by supplementing recombinant human transglutaminase 4 (rhTG-4) in treating osteochondral defects. RhTG-4 treatment induced the expression of integrin β1 and dynamic actin fiber, enhancing SDSCs adhesion to fibronectin. Supplementation of rhTG-4 significantly induced the proliferation of SDSCs encapsulated in the HA/COL/FG composite gel and increased the hardness of the extracellular matrix. Furthermore, supplementation of rhTG-4 significantly upregulated aggrecan and type II collagen mRNA. Pretreatment with integrin β1 siRNA markedly suppressed TG4-induced actin remodeling, activation mitogen-activated protein kinase (MAPK), and eventually the chondrogenesis-related genes. Moreover, transplantation of SDSCs encapsulated in HA/COL/FG/rhTG-4 composite gel in vivo yielded reconstructed tissue resembling native hyaline cartilage. These data suggest that rhTG-4 enhances cartilage regeneration of the SDSCs encapsulated in hydrogel in rabbits.
Impact statement
In this study, we investigated the effects of recombinant human transglutaminase 4 on the ability of synovium-derived mesenchymal stem cells encapsulated in a hyaluronic acid/collagen/fibrinogen composite gel to repair osteochondral defects. We believe that our study makes a significant contribution to the literature because it explores a method of improving an existing modality to mediate tissue repair.
Introduction
Tissue-engineered cartilage using mesenchymal stem cells (MSCs) has shown multiple differentiation potentials, easy availability, and rapid proliferation, rendering them a promising strategy for cartilage regeneration.1–3 Recent, injectable hydrogel-type scaffolds are effective for chondrogenesis of MSCs because these scaffolds have mechanical properties similar to those of articular cartilage components.4–6 Although the transplantation of MSCs encapsulated in hydrogel-type scaffolds is a promising strategy for the repair of damaged articular cartilage, hydrogel-type scaffolds-based cartilage tissue engineering has numerous limitations, including poor implanted cell adhesion, phenotypic alteration of cells, overly soft mechanical properties, and engraftment rates after implantation.6,7
Transglutaminases (TGs) are calcium-dependent enzymes catalyzing bond formation between glutamyl and lysyl residues of proteins.8,9 TGs can be used to enhance tensility and enzymatic resistance of collagen-based material. Purified TGs have been used to crosslink proteins to generate biocompatible and injectable tissue scaffolds.10–12 The transamidation reaction of TG-2 has been well studied and implicated in various physiological phenomena, including cell adhesion, differentiation, cell growth, and apoptosis. Recent studies have indicated that overexpression of TG-4 increases the adhesion and migration.13,14 TG-4 appears to share the function of regulating matrix adhesion with TG-2. TG-4 can induce the phosphorylation of FAK and paxillin, enhancing the cell-matrix adhesion. Furthermore, TG-4 interacts with integrin and initiates the cell-matrix adhesion sequences. The interaction between cells and their extracellular matrix (ECM) facilitates not only mechanical adhesion but also communication between the extracellular and intracellular environment.15,16
Therefore, we hypothesize that recombinant human transglutaminase-4 (rhTG-4) can enhance cartilage regeneration of synovium-derived mesenchymal stem cells (SDSCs) by enhancing cell-to-matrix adhesion. We previously developed an injectable hyaluronic acid/collagen/fibrinogen (HA/COL/FG) composite gel that encapsulated SDSCs. 17 To achieve more effective transplantation of SDSCs, this study aimed to investigate SDSCs encapsulated in HA/COL/FG composite gel by supplementing rhTG-4, thus enhancing SDSCs adhesion to the matrix.
Materials and Methods
Isolation of SDSCs from human synovium
After obtaining the ethical approval from the Institutional Review Board (protocol number: H-1104-110-359), isolation of human SDSCs was prepared using a previously described method. 18 In brief, synovial tissues were harvested from the knees of human donors during total knee arthroplasty for degenerative arthritis. The synovial tissues were digested with 0.02% collagenase (Sigma-Aldrich, St. Louis, MO) in a humidified 5% CO2 atmosphere. Thereafter, cells were washed and resuspended in a culture medium (high-glucose DMEM; Gibco, Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (Gibco).
Preparation of rhTG-4 using a baculovirus expression system
After cloning of human TG-4 into a baculovirus transfer vector pVL1392, expression of human TG-4 in Baculovirus Expression System (BD, Heidelberg, Germany) was performed in accordance with the manufacturer's instructions. In brief, cotransfection of 5 × 105 SF21 insect cells was carried out using the baculovirus DNA (1 μg) and the recombinant transfer vector pVL1392/hTG-4 (2.5 μg) to generate a recombinant baculovirus. After individual plaques were sonicated for 1 min, the supernatant was loaded on a NTA agarose column. Elution of His-tagged human TG-4 was achieved using an elution buffer.
SDSCs adhesion assay
The adhesion assay was carried out by directly counting the number of cells using the trypan blue staining. In total, 1 × 105 SDSCs were seeded into a fibronectin (2.5 μg/mL)-coating 12-well plate. rhTG-4 was then supplemented at different concentrations (0, 100, and 400 μg/mL). The well plates were incubated for 5, 15, 30, and 60 min at 37°C in 5% CO2. Unattached cells were washed out, the attached cells were enumerated using the trypan blue staining. To visualize the adherent cells, 50 μL of 0.5% crystal violet solution was added to each well in 12-well culture plate. The well plates were incubated for 20 min at room temperature. After washing with PBS, stained cells were visualized and photographed.
Immunofluorescence assay
To visualize cell adhesion and cytoskeletal proteins, SDSCs were treated with 400 μg/mL rhTG-4 and plated on fibronectin-coated glass coverslips for 60 min. Cystamine (Sigma C8707) at 100 μM was supplemented in the culture media to inhibit the TG-4 activity. Cells were fixed with 3% paraformaldehyde in PBS and permeabilized with 0.1% Triton X-100. After several washes with PBS cells were double-stained with 10 mg/mL polyclonal anti-beta-actin antibody (Sigma) and 10 mg/mL mouse mAb against human integrin β1. After incubation with primary antibodies, cells were stained with fluorescein-labeled goat anti–rabbit IgG and rhodamine-conjugated goat anti–mouse IgG (Chemicon). Stained cells were analyzed and photographed using a microscope (Nikon, Tokyo, Japan).
Culturing of SDSCs in HA/COL/FG composite gel and cell proliferation/viability assay
HA/COL/FG composite gel was generated using a previously described method. 15 In brief, FG was dissolved in an aprotinin solution (Green Cross). The FG solution was mixed with collagen (COL) and hyaluronic acid (HA). The mixture ratio of composite gel was HA:COL:FG = 0.5:0.5:2.0. In total, 2 × 107 SDSCs were resuspended in 200 μL of HA/COL/FG solutions. The HA/COL/FG composite gels and supplement of rhTG-4 in HA/COL/FG composite gels containing the SDSCs were transferred to six-well culture plates and cultured in a defined chondrogenic medium (low-glucose DMEM supplemented with 100 mM ascorbate-2-phosphate, 100 mg/mL pyruvate, 40 mg/mL proline, 100 nM dexamethasone, 100 ng/mL of BMP-2, and 10 ng/mL TGF-β1). The proliferation assay was carried out by counting the number of cells using trypan blue staining. 6 × 106 SDSCs were encapsulated in composite gels. At day 1, 3, 7, 14, and 28 of cultivation, the SDSCs encapsulated in composite gels were digested with 100 mL of 0.1% collagenase (Sigma-Aldrich) for 3 h at 37°C in a 5% CO2 atmosphere in the low-glucose DMEM containing 1% antimycotic-antibiotic solution and the number of cells was counted by trypan blue staining. The Live/Dead Viability Kit (Molecular Probes) was used for the assessment of the viability of encapsulated SDSCs. At day 7 after cultivation of samples, they were treated in a solution containing 2 mM ethidium homodimer-1 and 4 mM Calcein AM in PBS for 40 min. The samples were washed in PBS and thin slices of the stained samples were prepared on glass slides. They were observed using a fluorescence microscope. Live cells were visualized by a green color and dead cells were visualized by a red color
Evaluation of ECM hardness
SDSCs were harvested and resuspended into 96-well V-bottom culture plates at 1 × 106 cells/mL. The cells were incubated at 37°C with 5% CO2 for 24 h to generate micromass cultures. Thereafter, chondrogenic differentiation was carried out using a defined chondrogenic medium. Varying concentrations of rhTG-4 (0, 25, 50, 100, and 400 μg/mL) were then added. The medium was changed every 2 days, and micromasses were harvested on day 14. To evaluate the mechanical properties of the micromasses, the hardness (GPa) of 3D micromasses was evaluated using a nanoindentation instrument.
Reverse transcription-polymerase chain reaction
After extraction of total RNA using RNeasy Mini Kit (Qiagen), reverse transcription was performed using the cDNA Synthesis Kit (Fermentas Life Sciences, Waltham, MA). PCR amplification of type I COL, type II COL, type X COL, aggrecan, Sox9, or GAPDH was performed using AccuPower PCR PreMix (Bioneer) for 30 cycles. The following primers were used 15 : 5′-CGTGGTGACAAGGGTGAGAC-3′ and 5′-TAGGTGATGTTCTGGGAGGC-3′ for type I COL, 5′-TTCAGCTATGGAGATGACAATC-3′ and 5′-AGAGTCCTAGAGTGACTGAG-3′ for type II COL, 5′-CCCTTTTTGCTGCTAGTATCC-3′ and 5′-CTGTTGTCCAGGTTTTCCTGGCAC-3′ for type X COL, 5′-TGAGGAGGGCTGGAACAAGTACC-3′ and 5′-GGAGGTGGTAATTGCAGGGAACA-3′ for aggrecan, and 5′-CCCGATCTGAAGAAGGAGAGC-3′ and 5′-GTTCTTCACCGACTTCCTCCG-3′ for Sox9. PCR fragments were normalized with the levels of GAPDH (5′-ATTGTTGCCATCAATGACCC-3′ and 5′-AGTAGAGGCAGGGATGATGTT-3′). PCR products were separated through agarose gel electrophoresis (1% agarose), and DNA bands were visualized with ethidium bromide.
Evaluation of intracellular signal transduction pathway
SDSCs and HA/COL/FG composite gels were treated with 400 μg/mL rhTG-4. Integrin β1 siRNA was transfected to inhibit integrin β1 expression. An immunofluorescence assay was performed to visualize cell adhesion and cytoskeletal proteins. RT-PCR analysis was performed to evaluate the mRNA expression of type I COL, type II COL, type X COL, aggrecan, and Sox9. Western blotting was performed to evaluate the activation of mitogen-activated protein kinase (MAPK) by assessing the activation of extracellular signal-regulated kinase (ERK), c-Jun N-terminal Kinase (JNK), and p-38.
Surgical procedure
New Zealand white rabbits (3.5–4.0 kg weighting, 8-month-old males) were anesthetized through intramuscular administration of xylazine hydrochloride (5 mg/kg; Bayer) and ketamine hydrochloride (35 mg/kg; Yuhan). The anesthesia was maintained with isoflurane. The knee joints were incised by medial parapatellar approach, and the patella was eversed laterally to expose the femoral trochlea articular surface in rabbits. A full-thickness osteochondral defect model was established using a previously described method. 15 In brief, a full-thickness osteochondral defect (3 mm in depth and 4 mm in diameter) was made in the trochlea groove of the distal femur of New Zealand white rabbits. Fifty-six knee joints were divided into three groups. The osteochondral defects in the left knees of the control group (n = 16) were not treated with the HA/COL/FG composite gel; they served as controls. The osteochondral defects in knees of the Gel/SDSC group (n = 20) were filled with the HA/COL/FG composite gel (150 μL/defect) loaded with human SDSCs (2 × 107 cells/mL, 150 μL/defect). The osteochondral defects in the right knees of Gel/SDSC+TG-4 composite gels group (n = 20) were filled with the supplement of rhTG-4 in HA/COL/FG composite gels loaded with human SDSCs (2 × 107 cells/mL, 150 μL/defect). The experimental in vivo animal groups are summarized in Table 1. All experimental procedures related to the animal model were approved by the Institutional Animal Care and Use Committee.
Summary of the Experimental In Vivo Animal Groups
Macroscopic and histological assessment
For macroscopic scoring, the gross appearance of the defects was assessed through the international cartilage repair society (ICRS) cartilage repair assessment (degree of defect repair, macroscopic appearance, integration to border zone, and overall repair assessment). 19 Sagittal sections (5-mm thick) were stained with hematoxylin and eosin (H&E), and safranin-O/Fast Green. Histomorphologic findings (n = 8 at each time point) for each section were evaluated and scored by three investigators in accordance with the O'Driscoll histological grading parameters (cell morphology, matrix staining, structural integrity, thickness/defect filling, osteochondral junction, adjacent bonding, basal integration, cellularity, clustering/distribution, and adjacent cartilage). 20 Dil-labeled SDSCs distribution in the center of defect was performed. Dil (1,1′-Dioctadecyl-3,3,3′,3′-Tetramethylindocarbocyanine Perchlorate) staining was used for SDSCs distribution in cartilage. Fifty microliter of Dil (Molecular Probes) cell-labeling solution (2 mg/mL) was added directly to 10 mL culture media for labeling of 2 × 107 SDSCs. After 30 min of incubation at 37°C, the cells were washed in PBS twice. Then, the Dil-labeled SDSCs were encapsulated in the composite gels. After transplantation of SDSC encapsulated gel, Dil-labeled SDSCs distribution in cartilage was observed using a fluorescence microscope.
Statistical analysis
All data are presented as mean ± standard deviation values. The Mann–Whitney U test was performed to assess the differences in the mean values among different groups. All analyses were performed using SPSS 12.0 for Windows software (SPSS). p-Values <0.05 were considered statistically significant.
Results
Effect of TG-4 on synovial-derived stem cell adhesion
Upon treating SDSCs with different doses (0, 100, and 400 μg/mL) of rhTG-4 and incubation time (5, 15, 30, and 60 min), the adhesion to fibronectin increased in a time- and concentration-dependent manner (Fig. 1A). At 60 min, the number of attached SDSCs to fibronectin increased ∼4-fold in the 400 μg/mL TG-4-treated group, compared to the untreated group. The crystal violet assay at 5, 15, 30, and 60 min after culturing further confirmed the adhesion of SDSCs. The crystal violet assay further revealed that rhTG-4 treatment increased the adhesion of SDSCs to fibronectin (Fig. 1B).

rhTG-4 treatment improves the adhesion of SDSCs. In total, 3 × 104 SDSCs were seeded in fibronectin-coating 96-well plates. rhTG-4 was supplemented at different concentrations (0, 100, and 400 μg/mL). The well plates were incubated for 5, 15, 30, and 60 min at 37°C in 5% CO2.
rhTG4 treatment increases integrin β1 and actin remodeling in SDSCs
The immunofluorescence assay revealed that rhTG-4 stimulation upregulated integrin β1 (Fig. 2). Furthermore, we observed dynamic actin remodeling in the rhTG-4-treated group. Pretreatment with cystamine (TG-inhibitor) markedly downregulated integrin β1 and dynamic actin remodeling. These results indicate that rhTG-4 can upregulate integrin β1 and dynamic actin fibers, enhancing the SDSCs adhesion to fibronectin.

Treatment of rhTG4 induces integrin β1 and actin remodeling in SDSCs. One hour after 400 μg/mL rhTG-4 treatment of the SDSCs with or without the TG inhibitor; cystamine and immunofluorescence staining for integrin β1 and actin fiber expression levels was performed. Scale bar: 5 μm. TG, transglutaminase; SDSCs, synovium-derived mesenchymal stem cells.
Proliferation and viability of SDSCs in HA/COL/FG gel, including rhTG-4
SDSCs proliferation was significantly increased in the HA/COL/FG gel, including rhTG-4 composite gel group (Fig. 3A); however, based on the number of SDSCs, this difference was not significant. The number of SDSCs in the HA/COL/FG gel, including rhTG-4, was relatively higher up to day 7 and thereafter this difference was not significantly increased to day 28. The Live/Dead staining assay on day 7 of culturing confirmed the viability of SDSCs encapsulated in HA/COL/FG gel, including rhTG-4 (Fig. 3B). Fluorescence staining revealed high cell viability in HA/COL/FG gel, including rhTG-4, indicating that TG-4 potentially increases the proliferation and vitality of the encapsulated SDSCs. The relative ratio of live cells/dead cells on day 7 was significantly higher in the HA/COL/FG gel, including rhTG-4 group, than in the HA/COL/FG gel (Fig. 3C).

Proliferation and appearance of encapsulated SDSCs.
Hardness of ECM
Nanoindentation analysis revealed that rhTG-4 stimulation increased the hardness of 3D micromasses. After 1 week of incubation, ECM hardness was improved in accordance with the dose of rhTG-4 (Fig. 4). Doses of 0, 25, 50, 100, and 400 μg/mL of rhTG-4 yielded a hardness of 0.13, 0.22, 0.57, 0.68, and 0.94 GPa, respectively.
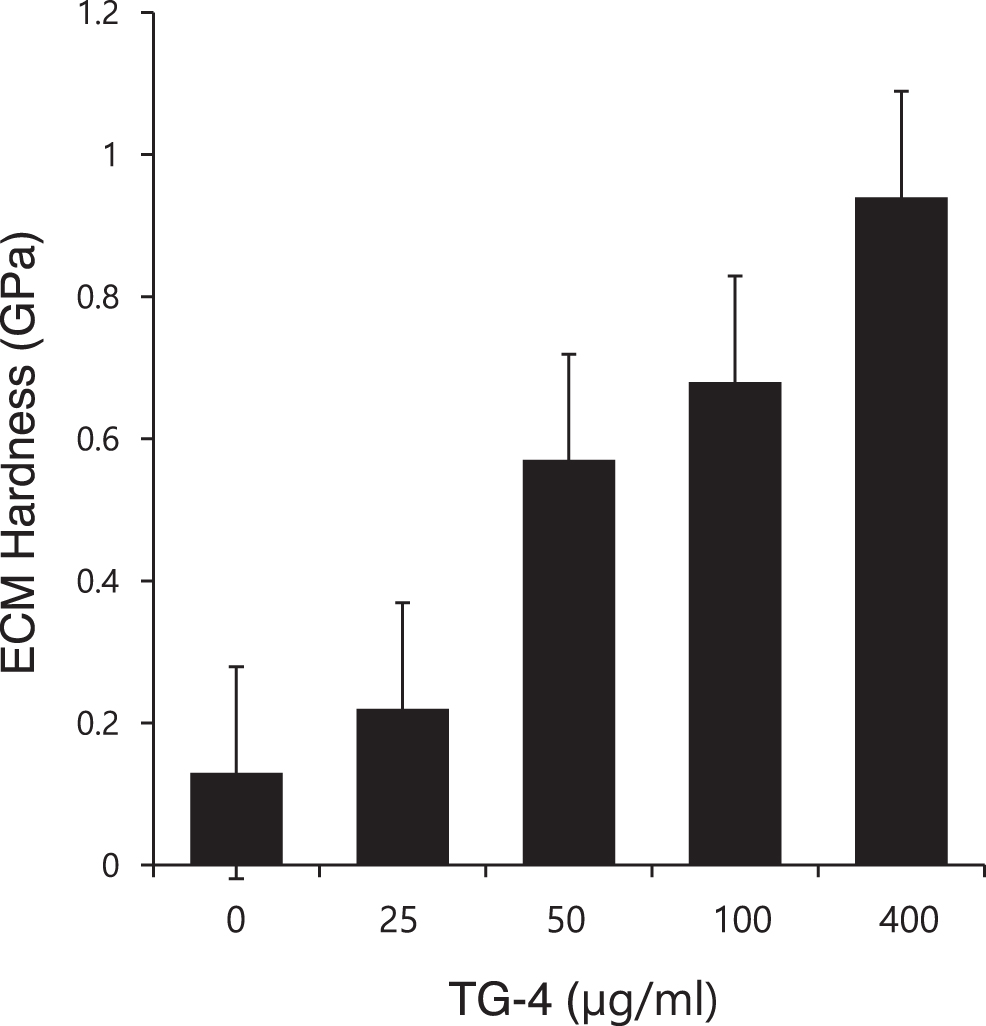
Hardness of ECM. Nanoindentation revealed that rhTG-4 stimulation increased the hardness of 3D micromasses. ECM, extracellular matrix; TG, transglutaminase.
The chondrogenic effects of rhTG4-hydrogel
At 28 days after culturing in the supplement of rhTG-4 in HA/COL/FG gel group, the relative expression levels of type II COL (rhTG-4 100 μg: 4.00-fold, rhTG-4 400 μg: 4.5-fold) and aggrecan (rhTG-4 100 μg: 4.1-fold, rhTG-4 400 μg: 4.7-fold) mRNA were significantly increased, compared to the HA/COL/FG composite gel group; however, expression levels of type X COL, Sox9, and type I COL were not significantly altered compared to those in the HA/COL/FG composite gel group (Fig. 5A, B).

Chondrogenic effects of rhTG4-hydrogel.
Expression of genes associated with the intracellular signal transduction pathway
Immunofluorescence analysis revealed that pretreatment with integrin β1 siRNA markedly decreased rhTG4-induced actin remodeling (Fig. 6A). MAPK was activated in an integrin β1 siRNA-dependent manner. ERK was not phosphorylated, and JNK and p-38 did not depend on integrin β1 siRNA treatment (Fig. 6B). Furthermore, chondrogenesis-related genes, including aggrecan and type II collagen, were downregulated (Fig. 6C). These data indicate that rhTG-4 potentially regulates SDSCs adhesion and expression of chondrogenesis-related genes via the activation of intracellular signal transduction pathways.

Expression of intracellular signal transduction pathway.
The in vivo potential of HA/COL/FG gel, including rhTG-4, to regenerate and repair osteochondral defects
As shown in Figure 7A, the defect lesion in the control group was partially covered with dark brown tissue at 1 week after transplantation. The defect lesion in the Gel/SDSC and the Gel/SDSC+TG-4 groups were partially filled with white tissue similar to intact cartilage. At 12 weeks, the surface of the control group displayed irregular white tissue, with a depression in the center of the defects. In both the Gel/SDSC and Gel/SDSC+TG-4 gel groups, the transplanted area was mostly repaired by a cartilage-like tissue. In the Gel/SDSC group, the defect lesion was almost repaired with newly formed cartilage tissues except for a slight depression at the center. In the Gel/SDSC+TG-4 group, the defect lesion was more filled with white tissue and displayed substantial integration between the newly formed cartilage and the surrounding original cartilage. Moreover, the newly formed cartilage tissue on the defects had more hyaline cartilage, which was densely stained with safranin-O in the Gel/SDSC+TG-4 group (Fig. 7B). At higher magnification, the cells had a columnar alignment and had a similar morphology to normal chondrocytes in the Gel/SDSC+TG-4 group (Fig. 7C). The macroscopic scores at 12 weeks for the control group, Gel/SDSC, and Gel/SDSC+TG-4 group were 3.40 ± 1.20, 5.20 ± 1.44, and 6.40 ± 1.36, respectively. The total macroscopic and histological scores at 12 weeks were significantly higher in Gel/SDSC+TG-4, than in the control group and Gel/SDSC group. The O'Driscoll histological scores at 12 weeks for the control group, Gel/SDSC group, and Gel/SDSC+TG-4 group were 11.00 ± 1.72, 15.00 ± 2.84, and 18.00 ± 2.24, respectively. The macroscopic scores at 12 weeks for the control group, Gel/SDSC, and Gel/SDSC+TG-4 group were 3.40 ± 1.20, 5.20 ± 1.44, and 6.40 ± 1.36, respectively (Fig. 7D). Dil-labeled SDSCs distribution was markedly increased at the center of defect in the Gel/SDSC+TG-4 group (Fig. 7E). These results indicate that the reconstructed tissue derived from the SDSC encapsulated in the supplement of rhTG-4 on HA/COL/FG composite gel resembles hyaline cartilage.

The in vivo potential of rhTG-4 supplementation in the HA/COL/FG to regenerate and repair osteochondral defects.
Discussion
This study aimed to investigate the effect of rhTG-4 supplement in a HA/COL/FG composite gel encapsulated with SDSCs in vitro and in vivo for the repair of osteochondral defect.
Hydrogel-type scaffolds are reportedly an effective system for chondrogenesis of MSCs, including SDSCs, because these scaffolds have mechanical properties similar to those of articular cartilage.4–6 Collagen, hyaluronic acid, and fibrin are major components of the cartilage ECM and are expected to provide a favorable environment for chondrogenesis of MSCs.21–23 Furthermore, we previously developed an injectable HA/COL/FG composite gel that encapsulated SDSCs. 17 The combination of HA/COL/FG may mimic the cartilage ECM, providing a chondrogenic environment for the encapsulated SDSCs. Moreover, SDSCs encapsulated in HA/COL/FG composite gel led to the formation of hyaline cartilage in damaged articular cartilage in vivo. However, scaffold-based cartilage tissue engineering is associated with potential issues, including poor implanted cell adhesion, phenotypic alteration of cells, overly soft mechanical properties, and engraftment rates after implantation.6,7 To achieve effective transplantation of SDSCs, we assessed SDSCs encapsulated in HA/COL/FG composite gels by supplementing rhTG-4, thus enhancing SDSCs adhesion to the matrix.
TG is present in several forms in tissue, blood, ECM, and on cell surfaces. All proteins in the TG family have transamidase activity and catalyze the formation of N-ɛ-(γ-glutamyl)lysine protein crosslinks. 8 Members of the TG family are expressed in numerous tissues and have numerous functions in processes, including posttranslational modification of proteins, cell adhesion and migration, ECM remodeling, and blood clotting.8,9,24,25 TG-4 in the prostate gland has a relatively restricted distribution pattern. 26 TG-4 levels in prostate tumor cells may be associated with their aggressiveness.13,14 For example, TG-4 overexpression in prostate tumor cells increases their invasiveness and migration. 13 TG-4 can induce the phosphorylation of FAK and paxillin, enhancing the cell-matrix adhesion. 14 Furthermore, TG-4 interacts with integrin and initiates cell-matrix adhesion sequences. A neutralizing antibody to integrin-β1 can abolish TG-4 induced cell-matrix adhesion. Matrix adhesion activities of TG-4 are observed in the TG-4 core domain because all constructs encoding core domains promote matrix adhesion, whereas the deletion of the core domain from TG-4 eliminates these activities. 14 Because the interaction between cells and the ECM not only mechanistically supports cell adhesion to the matrix but also mediates cellular signaling, we hypothesize that recombinant TG-4 can enhance the cartilage regenerative potential of SDSCs by enhancing adhesion to matrix. As shown in Figures 1 and 2, rhTG-4 increased the adhesion to fibronectin in a time- and concentration-dependent manner. Furthermore, rhTG-4 can induce the expression of integrin β1 and dynamic actin fibers, thus enhancing SDSCs adhesion to fibronectin and improving proliferation (Fig. 3). Finally, the ECM becomes harder in accordance with the dose of rhTG-4, thus providing a stable environment for SDSCs survival and differentiation (Fig. 4).
Adhesion to the matrix is a key step in promoting proliferation, cell differentiation, and migration. Cellular adhesion is primarily mediated by adhesion-related receptors, including integrin. Specific receptor-ligand binding is important in cartilage tissue engineering, since these interactions result in an intracellular signaling, thus altering the status of the cells. Adhesion prevents apoptosis, induces proliferation, and regulates protein production by chondrocytes, as in the case of ECM proteins and growth factors.27–29 The rhTG4 increased the proliferation of SDSCs and the expression of aggrecans and type II collagen mRNA (Fig. 5). The present results suggest that rhTG-4 supplementation in the HA/COL/FG composite gel further increases the chondrogenic effects in vitro, compared to the HA/COL/FG composite gel group. Furthermore, the underlying mechanisms modulated by rhTG-4 are revealed herein, indicating that activation of the intracellular signal transduction pathway is mediated by integrin β1. Binding with rhTG4 and integrin β1 in the cell wall induced actin remodeling and MAPK activation, eventually inducing chondrogenic mRNA expression (Fig. 6).
This study shows that rhTG-4 can enhance cartilage regeneration of SDSC encapsulated in hydrogels. As shown in Figure 7, the O'Driscoll histological scores and macroscopic scores at 12 weeks were significantly higher in the Gel/SDSC+TG-4 group than in the control group and Gel/SDSC group. The Dil-labeled SDSCs distribution was markedly increased at the center of defect in the Gel/SDSC+TG-4 gel group. Furthermore, host cell (Dil-unlabeled cells/DAPI positive cells) recruitment was further increased at the center of the defect in the Gel/SDSC+TG-4 gel group. Hence, rhTG-4 treatment might have recruited host cells by modulating the stiffness of the ECM. TG regulates ECM stiffness through posttranslational modifications of ECM components, including crosslinking.30–32
Conclusion
This study hypothesized that TG-4 supplementation potentially increases host cell recruitment, including bone marrow cells for cartilage regeneration. However, further studies are required to experimentally verify this hypothesis. The present results show that rhTG-4 supplementation in the HA/COL/FG composite gel mediates successful cartilage regeneration herein. Together, this study is the first, to our knowledge, to show that rhTG-4 can enhance cartilage regeneration of SDSCs encapsulated in hydrogels in rabbits.
Footnotes
Acknowledgments
Conception and experiment design: J.K.K., H.C.B., D.H.R., S.H.L., M.C.L., and H.S.H.; analysis and interpretation of the data: J.K.K, H.C.B., D.H.R., and H.S.H.; drafting of the article: J.K.K, H.C.B., and H.S.H.; critical revision of the article for important intellectual content: J.K.K., H.C.B., D.H.R., S.H.L., M.C.L., and H.S.H.; and final approval of the article: H.S.H.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Seoul National University Hospital Research Fund (04-2013-0560, 800-20130035, and 34-2014-0040), and the Korea Health Technology R&D Project through the Korea Health Industry Development Institute, funded by the Ministry of Health & Welfare, Republic of Korea (HI14C3339).