
Abstract
Glioblastoma (GBM) is the most common and lethal type of malignant brain tumor. A deeper mechanistic understanding of the invasion of heterogeneous GBM cell populations is crucial to develop therapeutic strategies. A key regulator of GBM cell invasion is interstitial flow. However, the effect of an interstitial flow on the invasion of heterogeneous GBM cell populations composed of glioma initiating cells (GICs) and relatively differentiated progeny cells remains unclear. In the present study, we investigated how GICs invade three-dimensional (3D) hydrogels in response to an interstitial flow with respect to their differentiation status. Microfluidic culture systems were used to apply an interstitial flow to the cells migrating from the cell aggregates into the 3D hydrogel. Phase-contrast microscopy revealed that the invasion and protrusion formation of the GICs in differentiated cell conditions were significantly enhanced by a forward interstitial flow, whose direction was the same as that of the cell invasion, whereas those in stem cell conditions were not enhanced by the interstitial flow. The mechanism of flow-induced invasion was further investigated by focusing on differentiated cell conditions. Immunofluorescence images revealed that the expression of cell–extracellular matrix adhesion-associated molecules, such as integrin β1, focal adhesion kinase, and phosphorylated Src, was upregulated in forward interstitial flow conditions. We then confirmed that cell invasion and protrusion formation were significantly inhibited by PP2, a Src inhibitor. Finally, we observed that the flow-induced cell invasion was preceded by nestin-positive immature GICs at the invasion front and followed by tubulin β3-positive differentiated cells. Our findings provide insights into the development of novel therapeutic strategies to inhibit flow-induced glioma invasion.
Impact statement
A mechanistic understanding of heterogeneous glioblastoma cell invasion is crucial for developing therapeutic strategies. We observed that the invasion and protrusion formation of glioma initiating cells (GICs) were significantly enhanced by forward interstitial flow in differentiated cell conditions. The expression of integrin β1, focal adhesion kinase, and phosphorylated Src was upregulated, and the flow-induced invasion was significantly inhibited by a Src inhibitor. The flow-induced heterogeneous cell invasion was preceded by nestin-positive GICs at the invasion front and followed by tubulin β3-positive differentiated cells. Our findings provide insights into the development of novel therapeutic strategies to inhibit flow-induced glioma invasion.
Introduction
Gliomas are classified into four grades depending on their malignancy. The most common and lethal type of primary malignant brain tumor in adults is glioblastoma (GBM), 1 which results in a median overall survival of less than 15 months. 2 A major cause of poor prognosis is tumor cell infiltration into surrounding normal brain tissues, which limits complete surgical resection. 3 In addition, GBM consists of glioma initiating cells (GICs)/glioma stem cells (GSCs) and progeny cells originating from the GICs/GSCs, 4 which results in the formation of heterogenous glioma cell populations. Non-GSCs are recognized to be more proliferative than GSCs. Although radiotherapy and chemotherapy are effective for proliferative non-GSCs, GSCs display low sensitivity to these therapies. 5 Consequently, therapeutic resistance occurs due to the formation of heterogeneous cell populations. In addition, it is possible that the cellular communication between GSCs and their differentiated progeny cells contributes to therapeutic resistance. A previous study showed that differentiated GBM cells promoted GSC tumor growth through a paracrine signaling. 6 Therefore, a deeper mechanistic understanding of the invasion of heterogeneous GBM cell populations is crucial to develop effective treatment strategies.
Glioma cell invasion follows preexisting routes, such as blood vessels, the subarachnoid space, and white matter tracts. 7 Perivascular spaces and the subarachnoid space have also been recognized as pathways for interstitial fluid flow and contribute to the clearance of interstitial solutes.8–10 In addition, interstitial fluid preferentially flows through specific white matter tracts, which correspond to the paths of dissemination of primary brain tumor cells. 11 These reports suggest that interstitial fluid flow plays a key role in GBM cell invasion. Recent in vitro studies of glioma cell lines or patient-derived GSCs in a microfluidic device and tissue culture insert have shown that fluid flow promotes GBM cell invasion in a three-dimensional (3D) matrix composed of hyaluronan and type I collagen by CD44-mediated mechanotransduction and autologous chemotaxis via CXCL12-CXCR4 signaling.12,13 Furthermore, these in vitro observations were confirmed via an in vivo experiment using convection-enhanced delivery, which is an experimental clinical technique used to increase drug distribution via local infusion. In mice with convection-enhanced delivery, interstitial flow enhanced the GBM cell invasion in a CXCR4 signaling-dependent manner. 14 However, the effect of an interstitial flow on the invasion of heterogeneous GBM cell populations composed of GICs/GSCs and relatively differentiated progeny cells remains unclear.
We utilized microfluidic culture systems that allow us to apply an interstitial flow to cells migrating from cell aggregates into a 3D hydrogel to investigate the effect of an interstitial flow on the invasion of heterogeneous GBM cell populations. 15 Our previous study using a microfluidic device showed that an interstitial flow and its direction affected the migration and proliferation of HepG2 cells, a hepatic cancer cell line, 16 confirming the suitability of the microfluidic device to study the effect of an interstitial flow on the invasion of tumor cell populations. In addition, to recapitulate heterogeneous GBM cell populations, we used a genetically induced GIC murine model, which faithfully resembles the histopathologic features of human GBM on orthotopic transplantation and shows a differentiation ability in response to serum exposure. 17 Recently, we reported that GICs formed heterogeneous glioma cell populations in a microfluidic device, and their invasion was significantly induced by a coculture with endothelial cells. 18
In the present study, we modulated the differentiation status of GICs by changing the culture media and investigated their invasion process in 3D hydrogels composed of type I collagen and Matrigel under an interstitial flow in a microfluidic device. This study aimed to investigate how GICs invade the 3D hydrogel in response to an interstitial flow with respect to their differentiation status. Furthermore, we investigated the expression of cell–extracellular matrix (ECM) adhesion-associated molecules, such as integrin β1, focal adhesion kinase (FAK), and phosphorylated Src, focusing on the flow-induced invasion of the GICs in differentiated cell conditions. Finally, we investigated the spatial distribution of the heterogeneous GBM cell populations during the invasion process. Our findings indicate that the invasion of GICs in differentiated cell conditions was significantly enhanced by the interstitial flow in a specific direction, whereas it was not enhanced by the interstitial flow in stem cell conditions. Furthermore, the flow-induced GIC invasion was preceded by nestin-positive immature cells at the invasion front and followed by tubulin β3 (TUBB3)-positive differentiated cells. These results provide insights into the development of novel therapeutic strategies to inhibit flow-induced GBM cell invasion.
Materials and Methods
Cell culture
GICs were established as previously described. 17 In brief, primary Ink4a/Arf-null neural stem cells and neural progenitor cells isolated from the subventricular zones of adult C57BL/6 mice were transfected with H-RasV12 and green fluorescent protein (GFP). All the animals used in the experiments received humane care, and the experimental protocol was approved by the Committee of Laboratory Animals following Keio University guidelines. The tumors formed by the orthotopic implantation of GICs were shown to display histopathologic features of human GBM, such as hemorrhage, necrosis, pronounced cell proliferation, pseudopalisading, and marked infiltration of the surrounding brain. 17 GICs were used to form the heterogeneous cell populations, which mimic tumor microenvironments better than conventional glioma cell lines. We have distinguished GICs and GSCs in our previous studies. GSCs are the transgenic cells isolated from primary tumors formed on the orthotopic implantation of GICs, and they are maintained in neurosphere culture. 19
The GICs were expanded in Dulbecco's modified Eagle's medium (DMEM)/F-12 (Sigma, St Louis, MO) supplemented with 20 ng/mL of recombinant human epidermal growth factor (PeproTech, Rocky Hill, NJ), 20 ng/mL of recombinant human basic fibroblast growth factor (PeproTech), 200 ng/mL of heparan sulfate (Sigma), 2% B27 supplement minus vitamin A (Invitrogen, Carlsbad, CA), and 1% antibiotic-antimycotic (Gibco, Gaithersburg, MD), which is called the stem cell medium. The GICs were maintained in a humidified 5% CO2 incubator at 37°C.
Fabrication of microfluidic devices
The microfluidic device consisted of micropatterned polydimethylsiloxane (PDMS, Sylgard 184; Dow Chemical, Midland, MI) and a coverslip to form two parallel channels for the culture medium and a channel for a 3D gel scaffold (Fig. 1A). The devices were fabricated as previously described.
15
In brief, SU-8 3050 photoresist (MicroChem, Newton, MA) was spun onto a 4-inch silicon wafer to a height of ∼135 μm. The wafer was baked and exposed to UV light through a photomask. The wafer was then baked again and developed in the SU-8 developer, resulting in SU-8 molds with defined channel patterns. A PDMS prepolymer was poured on the SU-8 molds and placed in an oven at 65°C for 6 h to cure the PDMS. The cured PDMS was peeled off from the wafers, cut into pieces, and punched to form inlets/outlets for the culture medium and gel. The PDMS pieces and coverslips were treated with air plasma and were bonded to each other to form microfluidic channels. The microfluidic channels were coated with poly-

Culture of GICs in a microfluidic device.
Three-dimensional invasion assay using a microfluidic platform
For a 3D culture in a microfluidic device, a hydrogel solution composed of 2.4 mg/mL type I collagen (pH 7.4; BD Biosciences, San Jose, CA) and 0.6 mg/mL Matrigel (Corning, Corning, NY) was injected into the gel channel and polymerized in a 5% CO2 incubator at 37°C for 30 min. The microfluidic channels were then filled with the stem cell medium, and subsequently, 1 × 104 cells were seeded into one of the channels. This seeding concentration was adjusted to form cell aggregates on the surface of the gel scaffold because we aimed to recapitulate the invasion behavior of the GICs departing from a tumor mass. The devices were incubated for 2 h to allow the cells to attach to the channel surface. After incubation, plastic tubes, which were used as medium reservoirs, were connected to the microchannels, and an interstitial flow across the gel scaffold was generated by a pressure difference of 5 mmH2O between the two microchannels (Fig. 1A). The interstitial flow velocity was calculated based on the displacement of the medium level in the reservoirs during 24 h and Darcy's law as previously described. 20 In this study, the flow velocity induced by 5 mmH2O was estimated to be 0.5 μm/s between the trapezoid PDMS posts, whereas the average velocity in the entire gel scaffold was 0.2 μm/s. The GICs were cultured in two types of interstitial flow conditions: forward flow, whose direction was the same as that of the cell invasion (arrows, Fig. 1A), and reverse flow, whose direction was opposite to that of the cell invasion. The cells were also cultured in static conditions as control experiments.
In stem cell conditions, the stem cell medium described above and an endothelial growth medium (EGM2; Lonza) without fetal bovine serum (FBS) were mixed at a 1:1 ratio. In differentiated cell conditions, DMEM/F-12 supplemented with 10% FBS and antibiotic-antimycotic and EGM2 were mixed at a 1:1 ratio. The culture medium was replaced every day. For inhibition experiments, 100 μM genistein (Merck, Darmstadt, Germany) and 10 μM PP2 (Merck) were added to the culture medium.
Immunofluorescence staining
The cells were fixed with 4% paraformaldehyde for 15 min and permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 5 min. Furthermore, the cells were treated with Block Ace (Sumitomo Dainippon Pharma, Tokyo, Japan) for 1 h to prevent nonspecific staining. The cells were then incubated with primary antibodies at 4°C overnight. Primary antibodies against nestin (1:300, mouse, Rat-401; Cell Signaling, Danvers, MA), TUBB3 (1:500, mouse, TUJ1; Covance, Princeton, NJ), integrin β1 (1:100, goat, AF2405; RSD, Minneapolis, MN), phosphorylated Src (Tyr419) (1:100, rabbit, 44-660G; Invitrogen), and FAK (1:100, rabbit, EP695; Abcam, Cambridge, UK) were used. The cells were then incubated for 2 h with secondary antibodies, namely Alexa Fluor 555-conjugated anti-mouse IgG (1:100, A31570; Invitrogen), Alexa Fluor 594-conjugated anti-rabbit IgG (1:100, A11037; Invitrogen), Alexa Fluor 647-conjugated anti-rabbit IgG (1:100, A31573; Invitrogen), and Alexa Fluor 555-conjugated anti-goat IgG (1:100, A21432; Invitrogen). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI, D1306; Invitrogen). Fluorescence images were obtained using a confocal laser-scanning microscope (LSM700; Carl Zeiss, Hallbergmoos, Germany).
Statistical analysis
Cell culture experiments were repeated at least twice to confirm the repeatability of the results. Data are presented as the mean ± standard error. Student's t-test was used to test for the differences between two groups, which were considered statistically significant at p < 0.01. Tukey's multiple comparison test was also used to test for the differences among three groups. Statistical analyses were performed using SPSS (IBM, Armonk, NY).
Results
Differentiation status modulated by the culture medium
First, we assessed the differentiation status of the GICs cultured in a microfluidic device. The cells were cultured under either stem cell or differentiated cell conditions. In stem cell conditions, the cells were cultured in a serum-free medium supplemented with growth factors, whereas in differentiated cell conditions, the cells were cultured in a serum-containing medium. The GICs used in the present study have the potential to differentiate into TUBB3-positive neurons, glial fibrillary acidic protein-positive astrocytes, and O4-positive oligodendrocytes. 17 However, the GICs preferentially differentiated into TUBB3-positive neurons in the present culture conditions. Therefore, we confirmed the formation of the heterogeneous cell populations focusing on nestin-positive stem cells and TUBB3-positive differentiated cells. The immunofluorescence staining images of the GICs on day 3 in stem cell conditions showed that the cells both in the gel scaffold and in the microchannel expressed nestin (Fig. 1B, stem cell condition). In contrast, few cells expressed TUBB3. On the contrary, cells in differentiated cell conditions were observed to express TUBB3 both in the gel scaffold and in the microchannel (Fig. 1B, differentiated cell condition). Nestin-positive cells were also observed in the differentiated cell conditions. These results demonstrated that the differentiation status of the GICs can be controlled by the culture medium, and that the GICs form heterogeneous cell populations composed of stem cells and relatively differentiated cells in differentiated cell conditions.
Invasion of GICs in stem cell conditions
The cells were cultured in interstitial flow conditions to investigate the effect of an interstitial flow on GIC invasion. Specifically, the GICs were cultured in stem cell conditions for 3 days under three different conditions: forward flow, reverse flow, and static conditions (Fig. 2A). The corresponding phase-contrast images showed that, regardless of the interstitial flow conditions, the GICs began to invade the gel on day 1 and proceeded across the gel scaffold until day 3 (Fig. 2A). Quantitative analysis revealed that no significant differences were detected between the invasion distance in the forward/reverse flow conditions and that in the static conditions on days 2 and 3; however, the invasion distance in the forward flow conditions was significantly longer than that in the reverse flow conditions on days 2 and 3 (Fig. 2B).
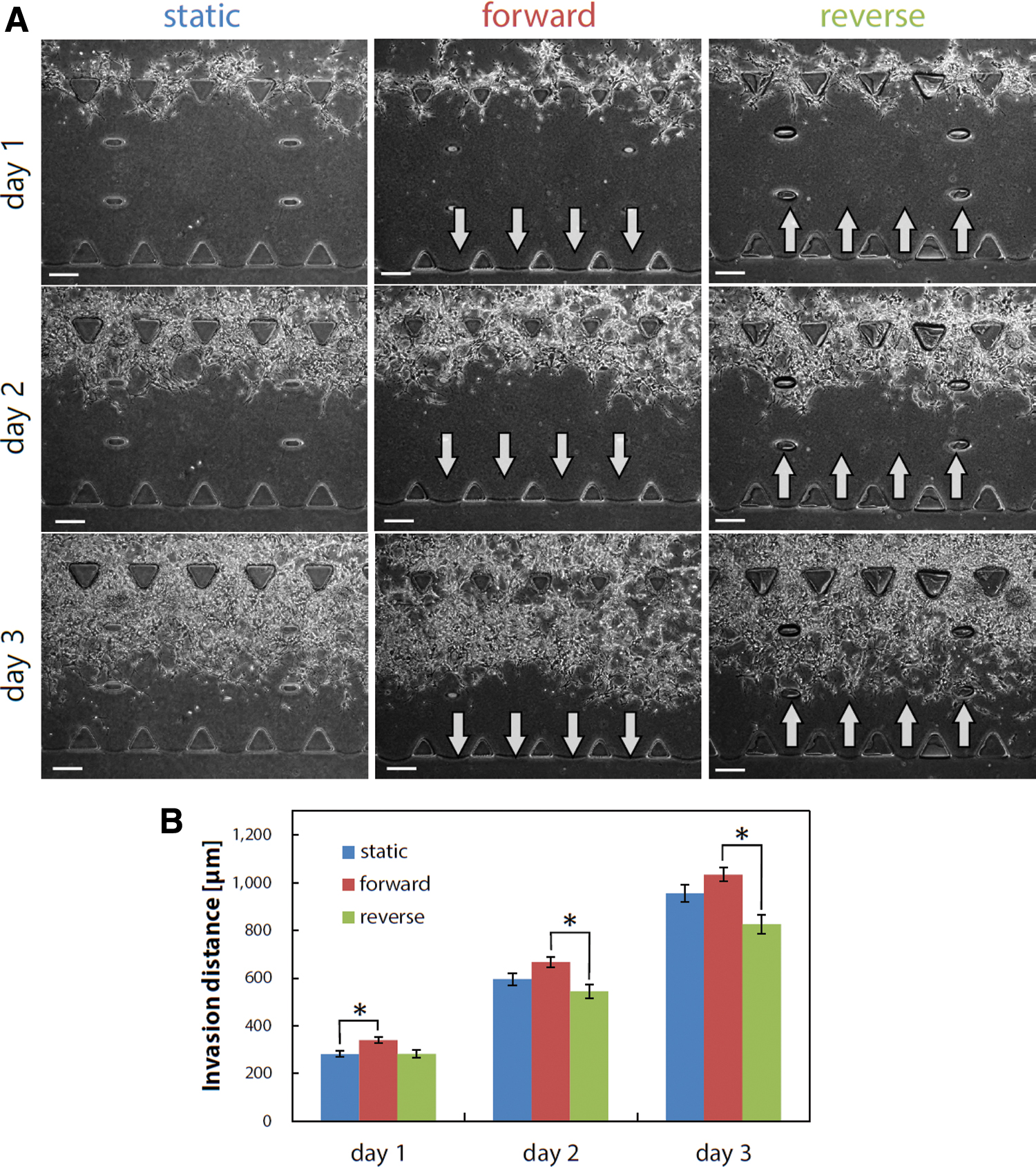
Invasion process of GICs in stem cell conditions during days 1–3.
During the invasion process, the GICs formed protrusions regardless of the interstitial flow conditions (arrowheads, Fig. 3A). In addition, some cells at the invasion front became detached from the follower cells and invaded individually. Quantitative analysis was performed to investigate the protrusions formed on day 3 further because the protrusion formation appeared to be a priming step for invasion. Consequently, it was observed that the protrusion length of 20–29 μm was most abundant, regardless of the interstitial flow conditions. In addition, no remarkable change in the total protrusion number was observed among the static, forward flow, and reverse flow conditions (Fig. 3B). Thus, no remarkable response to the interstitial flow was observed in the invasion process of the GICs in stem cell conditions.
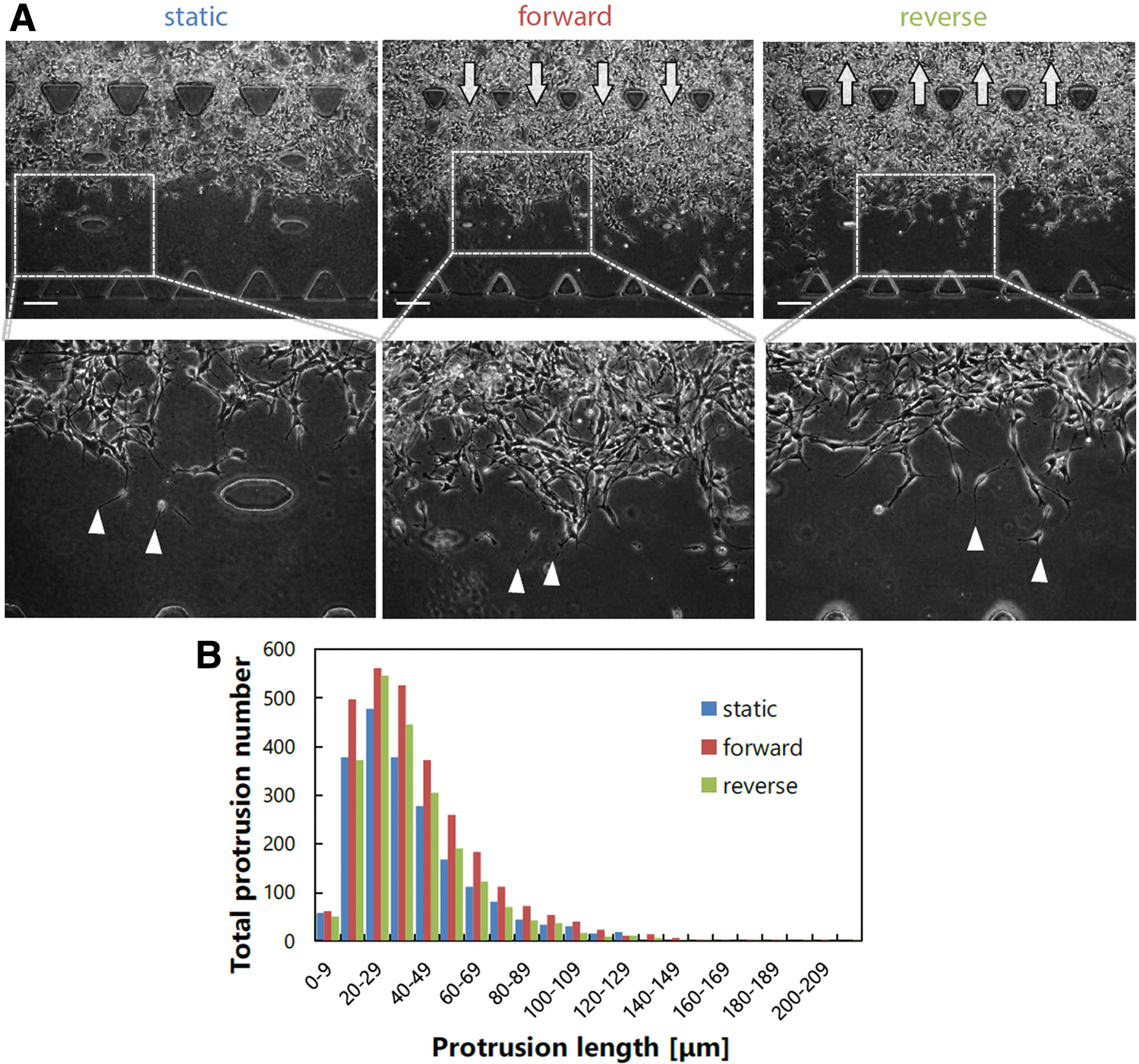
Protrusion formation of GICs in stem cell conditions.
Invasion of heterogeneous glioma cell populations in differentiated cell conditions
Subsequently, we investigated the effect of an interstitial flow on the invasion of heterogeneous GBM cell populations in differentiated cell conditions. The corresponding phase-contrast images during days 1–3 showed that the heterogeneous glioma cell populations actively invaded the hydrogel in the forward flow conditions, in contrast to the static and reverse flow conditions (Fig. 4A). Quantitative analysis revealed that the invasion distance significantly increased in the forward flow conditions throughout the culture period. For example, the invasion distance in the forward flow conditions on day 3 was 1.6 times greater than that in the static conditions and 2.6 times greater than that in the reverse flow conditions (Fig. 4B). Interestingly, the invasion distance in the reverse flow conditions was shorter than that in the static conditions (Fig. 4B).
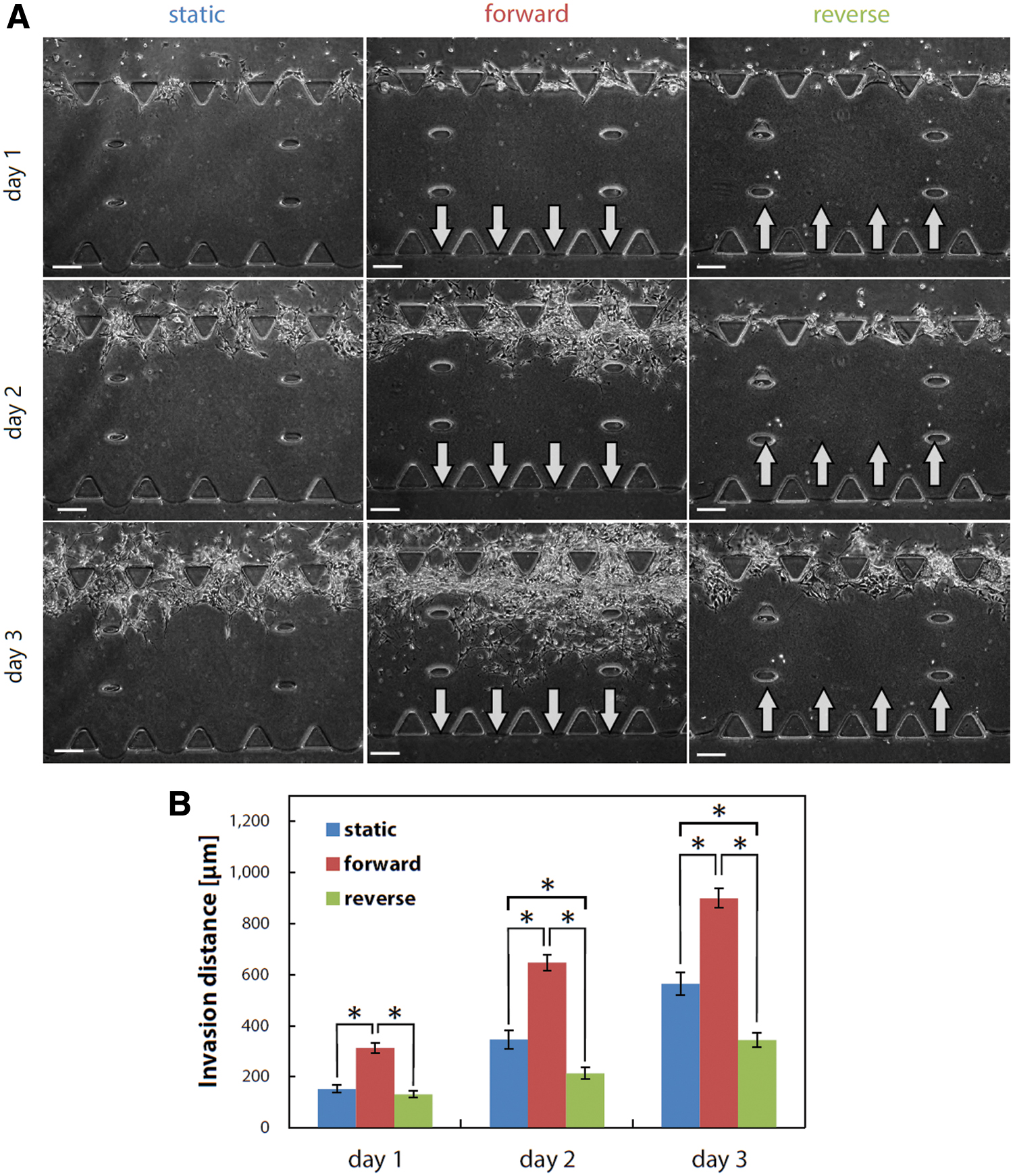
Invasion process of GICs in differentiated cell conditions during days 1–3.
The invasion process of the cells in differentiated cell conditions was further investigated by focusing on the cells at the invasion front. We observed that only the cells in the forward flow conditions formed extensive protrusions (arrowheads, Fig. 5A). In addition, the cells in differentiated cell conditions invaded the gel scaffold collectively in contrast to the invasion of individual cells in stem cell conditions. Quantitative analysis revealed that the total protrusion number in the forward flow conditions was the largest, whereas that in the reverse flow conditions was the smallest (Fig. 5B).
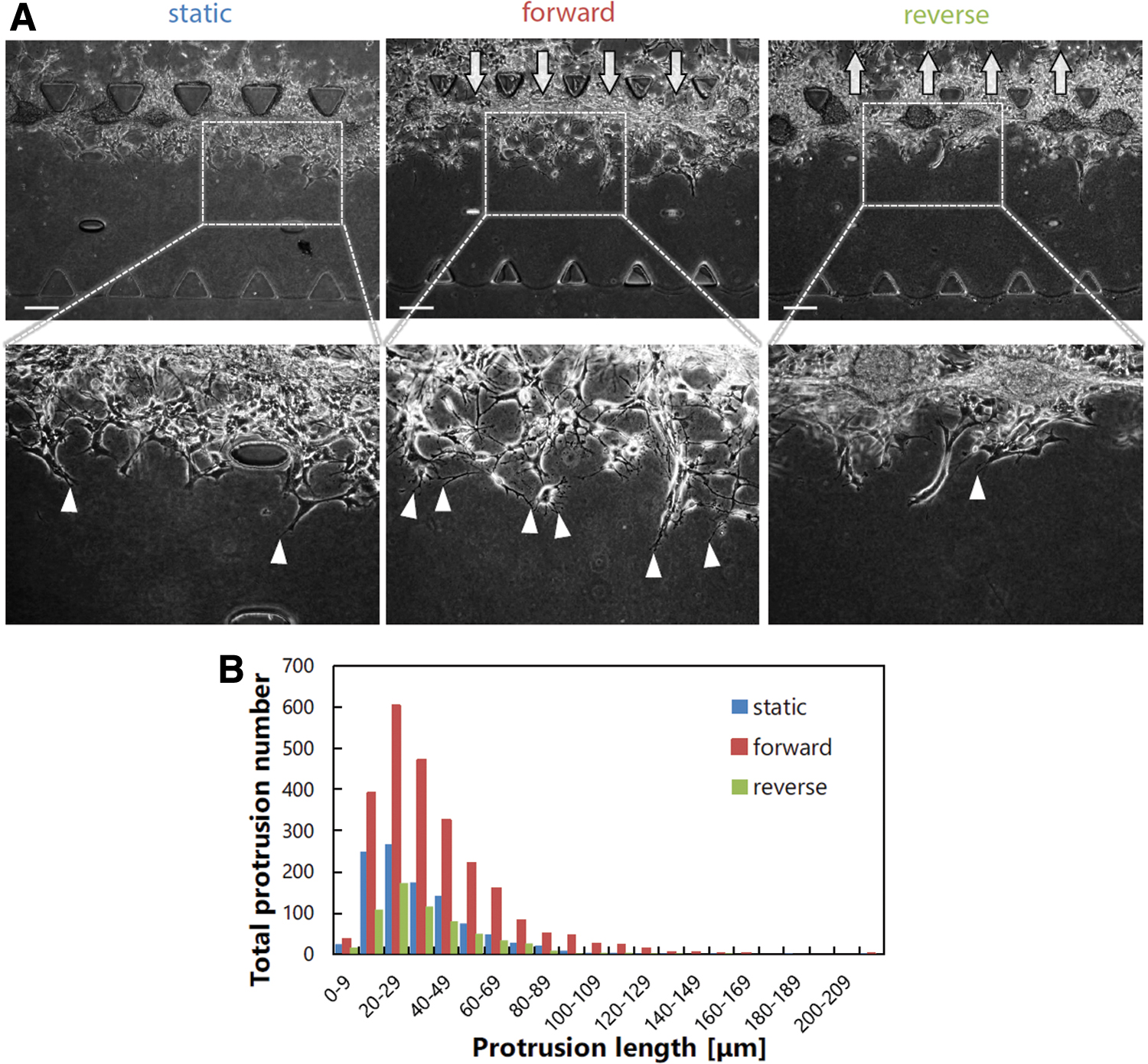
Protrusion formation of GICs in differentiated cell conditions.
The results of invasion distance and total protrusion number suggest that the invasion of heterogeneous GBM cell populations in differentiated cell conditions is flow responsive. Specifically, the forward interstitial flow promotes invasion, whereas the reverse interstitial flow suppresses invasion.
Promotion of the invasion of heterogeneous glioma cell populations by forward interstitial flow via adhesion-associated proteins
An interstitial flow was shown to induce the phosphorylation of FAK and the upstream migration of MDA-MB-231, a human breast cancer cell line. 21 FAK-mediated signaling is also involved in flow-induced endothelial cell migration and angiogenesis. 22 Therefore, we focused on cell–ECM adhesion-associated molecules to investigate the underlying mechanism regulating the flow-responsive invasion of heterogeneous glioma cell populations in differentiated cell conditions. The immunofluorescence staining images showed that the cells in the forward flow conditions exhibited a stronger expression of integrin β1, FAK, and phosphorylated Src compared with the cells in the static conditions (Fig. 6A–C).
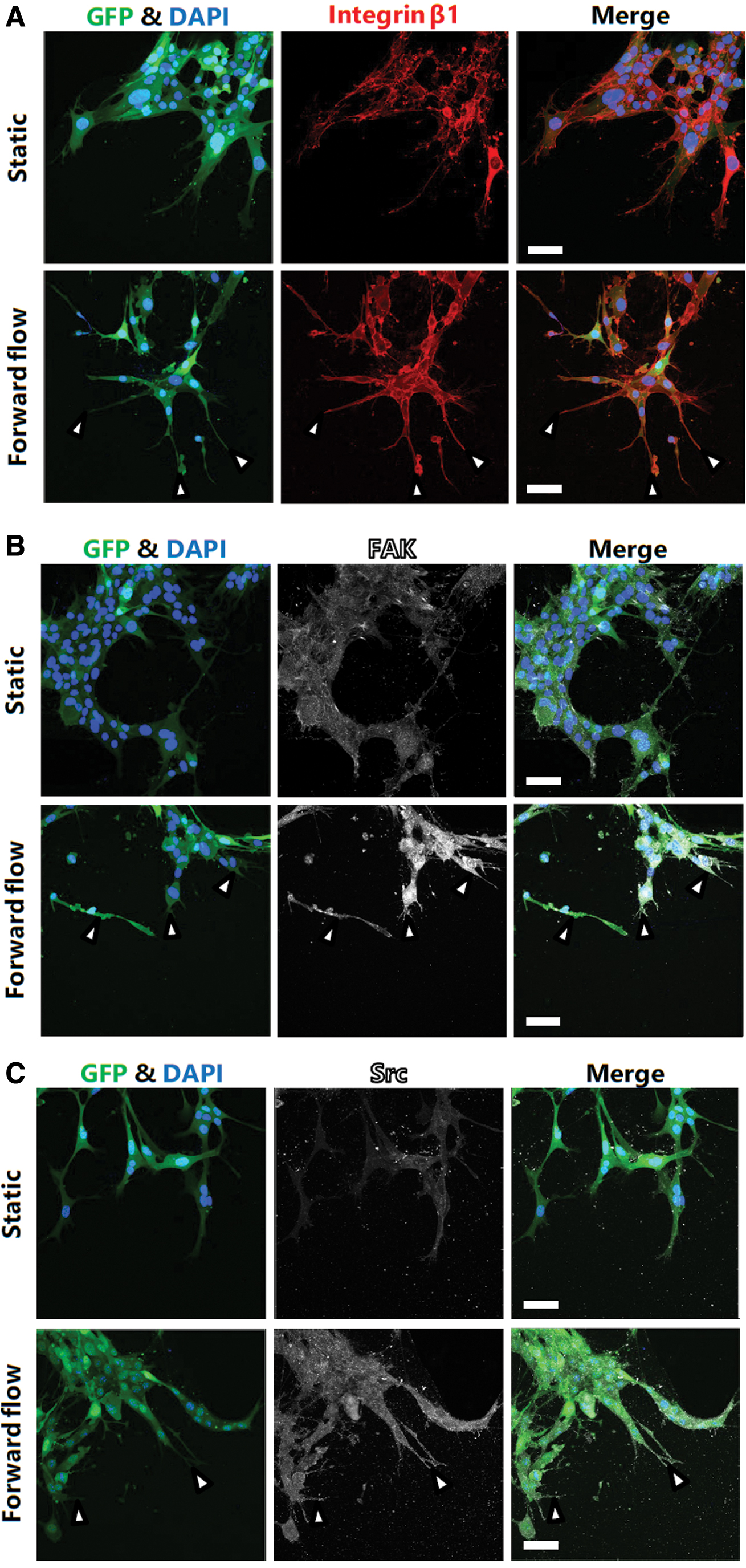
Immunofluorescence staining images of GICs in differentiated cell conditions. Cells cultured in both static and forward interstitial flow conditions were fixed on day 3 and stained for integrin β1
We treated the cells with genistein, a tyrosine kinase inhibitor, or PP2, which is a Src inhibitor, to confirm the role of these molecules in mediating the flow-responsive invasion further. Treatments with genistein and PP2 significantly reduced the invasion distance of heterogeneous glioma cell populations in the forward flow conditions on day 3 (Fig. 7A). In addition, both treatments reduced the number of protrusions formed during days 1–3 (Fig. 7B). These results indicate that, in differentiated cell conditions, cell–ECM adhesion-associated molecules, including Src kinase, play a critical role in mediating the flow-responsive invasion and protrusion formation of heterogeneous glioma cell populations.
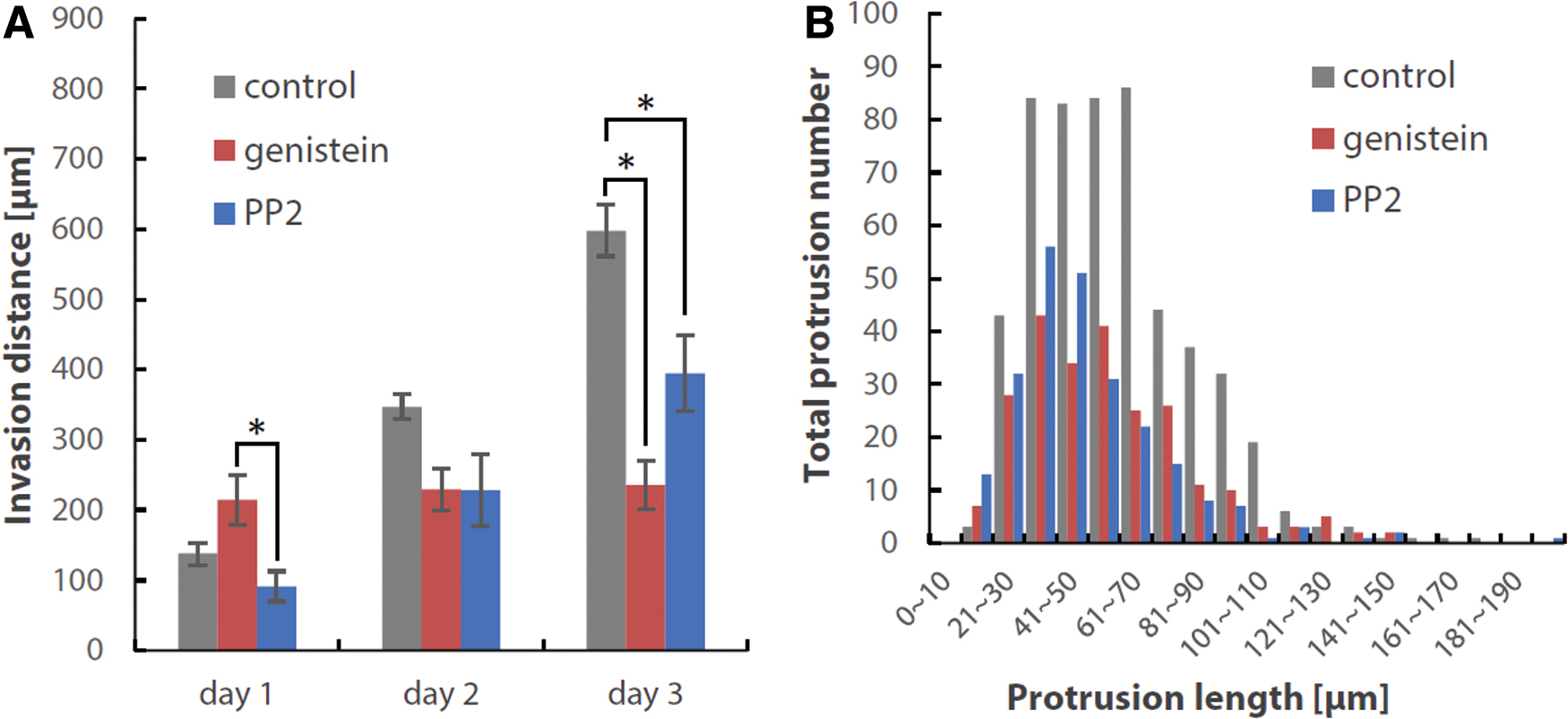
Inhibition experiments using genistein (tyrosine kinase inhibitor) and PP2 (Src inhibitor).
Nestin-positive GICs preceding the invasion of heterogeneous glioma cell populations under forward interstitial flow
Finally, we investigated the spatial distribution of heterogeneous glioma cell populations during the invasion process. In particular, we focused on the differentiation status of the cells at the invasion front in differentiated cell conditions under forward interstitial flow. The immunofluorescence staining images revealed that the cells at the invasion front were positive for nestin but negative for TUBB3, whereas the follower cells were positive for both nestin and TUBB3 (Fig. 8A). This indicates that the invasion was preceded by nestin-positive immature GICs and followed by TUBB3-positive relatively differentiated cells. The ratio of the nestin/TUBB3 projection area to the GFP projection area was quantified both in the medium channel and in the gel region to confirm this tendency further (Fig. 8B). The results showed that the nestin-positive area was 3.1 times and 1.8 times larger than the TUBB3-positive area in the gel region and in the medium channel, respectively (Fig. 8C). In particular, the nestin-positive area was significantly larger than the TUBB3-positive area in the gel region. These results suggest that the invasion of heterogeneous glioma cell populations in differentiated cell conditions under forward interstitial flow was preceded by nestin-positive GICs.
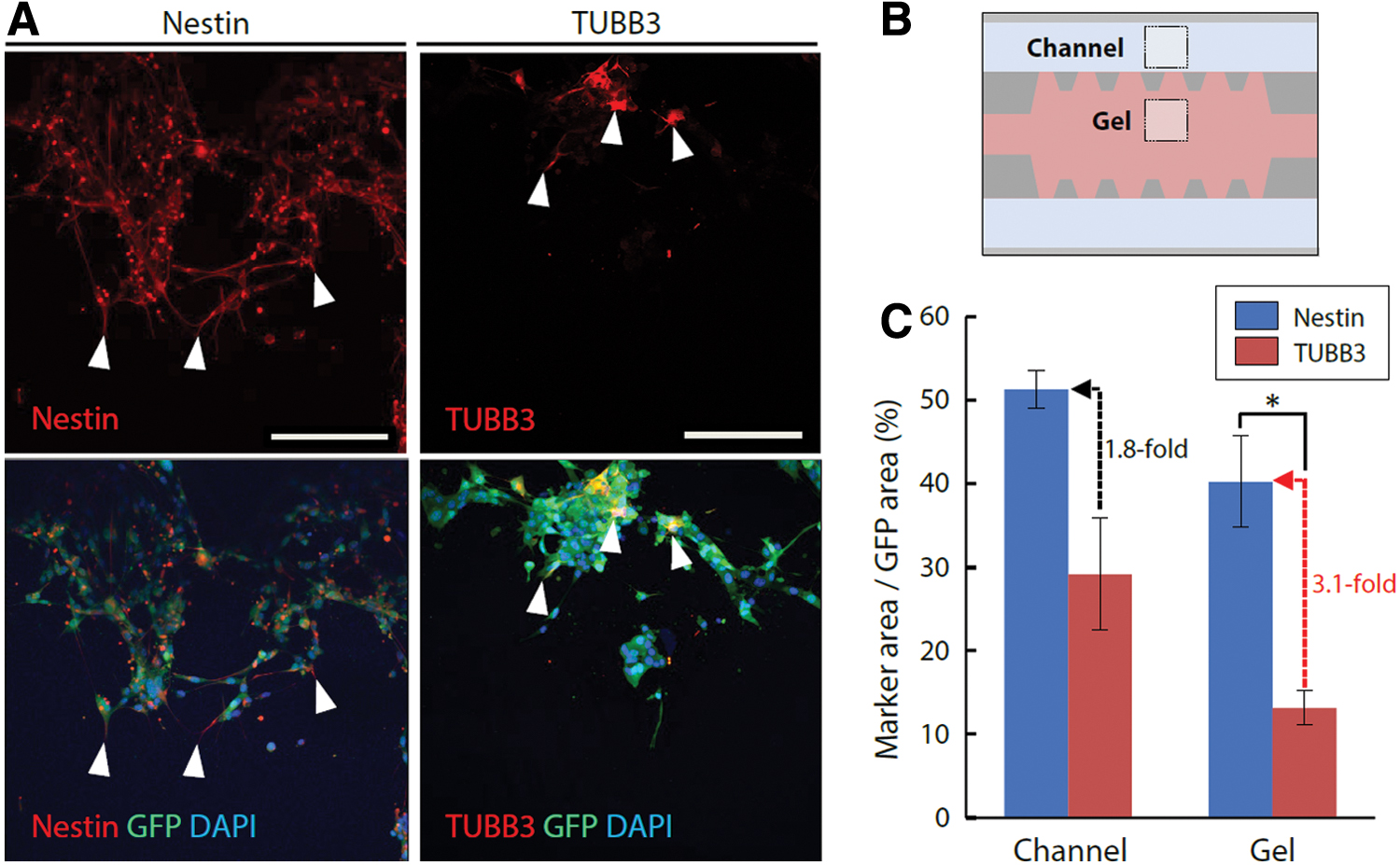
Spatial distribution of immature GICs and relatively differentiated cells at the invasion front.
Discussion
Validity of interstitial flow velocity in microfluidic device
In the present study, we used a microfluidic device to investigate the effect of an interstitial flow on GIC invasion in 3D gels. The direction and velocity of the interstitial flow can be controlled using the microfluidic device. Specifically, the interstitial flow velocity was generated by a pressure difference of 5 mmH2O across the gel scaffold. The velocity induced by 5 mmH2O in the present study was estimated to be 0.5 μm/s around the trapezoid PDMS posts placed to hold the gel scaffold, whereas the average velocity in the entire gel scaffold was 0.2 μm/s. This velocity range of 0.2–0.5 μm/s is comparable to the values reported previously. 23 In addition, a previous study using magnetic resonance imaging showed that the interstitial flow velocity in human GBM cell line xenograft models ranged from 0 to 2.0 μm/s. 24 Based on these previous studies, we confirmed that the interstitial flow velocity generated in the present study mimics the interstitial flow velocity in GBM. In the present study, we focused on the presence of the interstitial flow induced by 5 mmH2O. However, the invasion response may be altered by the magnitude of interstitial flow. Further investigations will be required to clarify the effect of interstitial flow magnitude on the invasive ability of the GICs.
Response of GBM cell populations to interstitial flow depending on their differentiation status
One of the key findings in the present study is that the response of GBM cell populations to the interstitial flow varied depending on their differentiation status. The invasion distance of GICs was significantly increased by the forward interstitial flow in differentiated cell conditions (Fig. 4), whereas it was not increased by the interstitial flow in stem cell conditions (Fig. 2). Similarly, the protrusion formation of the cells was remarkably increased by the forward interstitial flow in differentiated cell conditions (Fig. 5), whereas it was not increased by the interstitial flow in stem cell conditions (Fig. 3). These results suggest the possibility to attenuate the invasion of GBM cells by physically modulating the interstitial flow. It is also possible to develop new therapeutics focusing on the inhibition of signaling pathways upregulated in the flow-induced invasion process although further investigations will be required to clarify the signaling molecules. In addition, these results suggested that the GICs in differentiated cell conditions acquired the ability to respond to the interstitial flow, whereas those in stem cell conditions did not respond to the interstitial flow. This can be explained by two factors that the medium composition or cellular communication between heterogeneous cell populations alters the sensitivity of the GICs to the interstitial flow, which enhances the invasive ability of the GICs. Important differences in the medium composition are the presence of FBS and growth factors, such as EGF and bFGF. The GICs in stem cell conditions were exposed to EGF and bFGF, while those in differentiated cell conditions were exposed to FBS. The addition of FBS or the exclusion of EGF and bFGF were not essential to acquire the ability to respond to the interstitial flow in the present study because the GICs in stem cell conditions responded to the interstitial flow although the response was less significant compared to the cells in differentiated cell conditions. However, the addition of FBS or the exclusion of EGF and bFGF appeared to enhance the ability of the cells to respond to the interstitial flow. The possible mechanisms of the GIC response to the interstitial flow are discussed below.
Our results also suggested that the protrusion formation might be correlated with the cell invasion. In the present study, the GICs in differentiated cell conditions extended the protrusions at the downstream side of the cells in the forward interstitial flow, which corresponded to the maximum invasion distance. In our previous study using hepatic cancer (HepG2) cells, the cells extended protrusions in response to an interstitial flow. 16 However, the HepG2 cells extended the protrusions at the upstream side of the cells in the interstitial flow. This direction of protrusion formation in HepG2 cells is opposite to that of the GICs in the present study. Another previous study using human breast cancer cells (MDA-MB-231) demonstrated that the cells also extended protrusions at the upstream side of the cells in an interstitial flow, resulting in migration against the flow.21,25 These previous studies are inconsistent with the present study in terms of the direction of protrusion formation, suggesting that the cellular response to the interstitial flow is different depending on the cell type. It is possible that the responses of HepG2 and MDA-MB-231 cells were similar because they are both epithelial cells, whereas GICs are nonepithelial cells.
The modality of cell invasion was also different depending on the differentiation status of GICs. The cells exhibited individual cell invasion in stem cell conditions, whereas they exhibited collective cell invasion in differentiated cell conditions. A previous study investigated how N-cadherin regulates cancer cell invasion using prostate cancer cells (PC3) as a model system and demonstrated N-cadherin-dependent collective cell invasion. 26 Furthermore, a recent study showed that the collective invasion of human GBM cells depended on the cell–cell junctions stabilized by N-cadherin, β-catenin, and p120-catenin, and the downregulation of these proteins led to cell individualization during migration on a substrate coated with a reconstituted basement membrane. 27 As reported in these previous studies, cell–cell adhesion is important in regulating cancer cell invasion. In the present study, the expression of cell–cell adhesion proteins might have changed depending on their differentiation status, which led to the different modalities of cell invasion. The difference between individual cell invasion and collective cell invasion is important in the context of surgical treatment because individual cell invasion may result in the disseminated infiltration of glioma cells to the surrounding brain tissues.
The modality of cell invasion was also affected by the mechanical property of the gel used in the present study. The Young's modulus of nontumor gliotic tissues ranges from 10 to 180 Pa, while that of GBMs ranges from 70 to 13,500 Pa. 28 In addition, the stiffness of 1.7 mg/mL rat tail collagen hydrogel was measured by atomic force microscopy, resulting in the Young's modulus of 51 Pa. 29 These results suggest that the stiffness of the hydrogel used in this study, which was composed of 2.4 mg/mL type I collagen and 0.6 mg/mL Matrigel, might range in the value of the brain tissue in vivo.
Characteristic spatial distribution of heterogeneous GBM cell populations during the invasion process
Another key finding is the characteristic spatial distribution of heterogeneous GBM cell populations in differentiated cell conditions during the invasion process in the forward interstitial flow. As confirmed by the immunofluorescence staining images, the cells in differentiated cell conditions formed heterogeneous cell populations composed of nestin-positive immature cells and TUBB3-positive differentiated cells (Fig. 1B). We then demonstrated that the cells at the invasion front expressed nestin, whereas few cells expressed TUBB3 at the invasion front (Fig. 8). Instead, the follower cells expressed TUBB3. These results suggested that the flow-induced cell invasion was preceded by nestin-positive cells and followed by TUBB3-positive cells. This characteristic spatial distribution of heterogeneous GBM cells is consistent with our previous report demonstrating that the endothelium-induced invasion of heterogeneous GBM cells was preceded by nestin-positive GICs. 18 Although the cell invasion was induced by different factors, either the forward interstitial flow (a biomechanical factor) or the coculture with endothelial cells (a biochemical factor), similar characteristic spatial distributions were observed during the invasion process. These results suggest that the GICs are potentially invasive compared with relatively differentiated GBM cells. Indeed, the invasion distance of the GICs in stem cell conditions was ∼900 μm on day 3 in static conditions (Fig. 2) while that of the GICs in differentiated cell conditions was ∼600 μm on day 3 in static conditions (Fig. 4). These results suggest that the GICs in stem cell conditions are more invasive than those in differentiated cell conditions. In addition, previous studies reported that GSCs are more invasive than nonstem tumor cells.30,31 Further investigation will be necessary to clarify whether the preceding GICs induce the invasion of the following differentiated GBM cells. In addition, we assumed that the interstitial flow did not alter the differentiation of GICs. Although there are no previous studies demonstrating the induction of GIC differentiation by the interstitial flow, this possibility cannot be ruled out.
Possible mechanisms of flow-induced invasion involving focal adhesion proteins
Our findings indicate that the phosphorylation of Src mediates the flow-induced protrusion formation and the invasion of heterogeneous GBM cells. It is known that Src binds to FAK and the resulting form of active Src/FAK complex promotes the phosphorylation of Src substrates, such as p130Cas and paxillin, which subsequently stimulates the Crk/Dock180/Rac pathway, resulting in actin polymerization, protrusion formation, and migration.32,33 Previous studies using a human breast cancer cell line (MDA-MB-231) in a 3D collagen gel suggest that an interstitial flow applies tension in integrin β1 and activates focal adhesion complexes, including FAK phosphorylation, at the upstream side of the cell, which results in upstream migration.21,25 However, our results showed that the forward interstitial flow promoted the invasion of heterogeneous GBM cells, whereas the reverse interstitial flow suppressed the invasion, suggesting that the flow-induced increase in tension across integrins is an unlikely mechanism for the flow-responsive phosphorylation of Src and the invasion of heterogeneous GBM cell populations in the present study. An alternative mechanism for the flow-induced activation of focal adhesion proteins was reported in a study showing that rat vascular smooth muscle cells in type I collagen gel displayed the flow-induced phosphorylation of FAK via heparan sulfate proteoglycans. 34 Glycocalyx components, namely heparan sulfate and hyaluronan, were also shown to mediate the flow-enhanced migration of renal cell carcinoma (SN12L1) in a 3D collagen gel. 35 Consequently, the forward interstitial flow might activate the FAK/Src complex via the glycocalyx, which results in actin polymerization, protrusion formation, and invasion.
We demonstrated that the enhanced invasion of the GICs in differentiated cell conditions under the forward interstitial flow was significantly attenuated by PP2, a Src inhibitor. This suggests the possibility to attenuate the invasion of glioma cells by a Src inhibitor. Several Src inhibitors have been approved for the treatments of diseases. 36 For example, Bosutinib and Dasatinib are currently approved for the treatment of chronic myelogenous leukemia and acute lymphoblastic leukemia. Vandetanib, an inhibitor of EGFR, VEGFR, and Src family kinase, has been also approved for the treatment of medullary thyroid carcinoma. Similarly, our results implicated the potential of these Src inhibitors for the treatment of GBM. Indeed, clinical trials have been set up to evaluate the GBM treatment with the Src inhibitors although their results are not conclusive yet. 37
We also observed that the forward interstitial flow induced the expression of integrin β1 and FAK in heterogeneous GBM cell populations. The expression of proteins involved in focal adhesion assembly was shown to be promoted by the transcriptional regulator Yes-associated protein (YAP), 38 a key mediator translating mechanical cues, such as substrate rigidity and interstitial shear stress, into transcriptional programs. 39 Several mechanisms for the activation of YAP by fluid flow have been proposed in previous studies. In HEK293A cells, flow-induced YAP activation was shown to be mediated by caveolin-1, 40 a component of caveolae. Caveolin-1 was also reported to be enriched in GBM cells. 41 In prostate cancer cell lines, fluid shear stress stimulates ROCK/LIMK/cofilin signaling, which stabilizes F-actin cytoskeletons and activates YAP1, resulting in enhanced cell motility. 42 It is possible that caveolae or ROCK/LIMK signaling mediates the interstitial-flow-induced activation of YAP, which upregulates the expression of focal adhesion molecules and promotes focal adhesion assembly and invasion.
The mechanism by which GICs sense interstitial flow is currently unknown. Further investigation will be required to elucidate the mechanism, including a mechanosensor and related signaling molecules, which contributes to develop new therapeutics by inhibiting the flow-induced GBM invasion.
Conclusion
In conclusion, we showed that the invasion of heterogeneous GBM cell populations was significantly promoted by a forward interstitial flow via focal adhesion proteins and preceded by nestin-positive immature GICs. Our findings indicate that the forward interstitial flow enhances GIC progression by promoting their invasive ability depending on their differentiation status. This provides deeper mechanistic insights into flow-induced invasion to develop an effective therapy for GBM.
Footnotes
Acknowledgments
We thank Editage for English language editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by AMED under grant number JP20gm5810026 (R.S.).