
Abstract
Tissue-engineered skeletal muscle is a promising novel therapy for the treatment of volumetric muscle loss (VML). Our laboratory has developed tissue-engineered skeletal muscle units (SMUs) and engineered neural conduits (ENCs), and modularly scaled them to clinically relevant sizes for the treatment of VML in a large animal (sheep) model. In a previous study, we evaluated the effects of the SMUs and ENCs in treating a 30% VML injury in the ovine peroneus tertius muscle after a 3-month recovery period. The goal of the current study was to expand on our 3-month study and evaluate the SMUs and ENCs in restoring muscle function after a 6-month recovery period. Six months after implantation, we found that the repair groups with the SMU (VML+SMU and VML+SMU+ENC) restored muscle mass to a level that was statistically indistinguishable from the uninjured contralateral muscle. In contrast, the muscle mass in the VML-Only group was significantly less than groups repaired with an SMU. Following the 6-month recovery from VML, the maximum tetanic force was significantly lower for all VML injured groups compared with the uninjured contralateral muscle. However, we did demonstrate the ability of our ENCs to effectively regenerate nerve between the distal stump of the native nerve and the repair site in 14 of the 15 animals studied.
Impact Statement
Volumetric muscle loss (VML) is a clinically relevant problem for which current treatment options are lacking and for which tissue-engineered skeletal muscle presents a promising novel therapeutic option. However, the fabrication of tissues of clinically relevant sizes is necessary for advancement of the technology to the clinic. This study aimed to evaluate the efficacy of our scaled-up tissue-engineered skeletal muscle to treat VML in a large animal (sheep) model after a 6-month recovery.
Introduction
Volumetric muscle loss (VML) accounts for over 4.5 million reconstructive procedures performed annually and results in $800 billion dollars in health care costs in the United States alone.1–3 Furthermore, it is estimated that VML injuries affect 53% of military trauma patients.4,5 VML is a major problem in regenerative medicine and is defined as the traumatic or surgical loss of 20–30% or more of the volume of skeletal muscle resulting in sustained functional impairment, which in many cases, is accompanied by peripheral nerve injuries or physical deformity. 3 Following VML, skeletal muscle's innate capacity for regeneration is overwhelmed and fibrotic scar tissue invades in the defect site.4,6,7 The current standard of care for VML is to replace lost muscle by translocating autogenic tissue from a donor site into the wound and physical therapy.8–10 However, muscle flaps are limited by tissue availability and donor-site morbidity and physical therapy has shown limited success in restoring muscle function.10,11
These autograft techniques are often complicated by poor vascularization, denervation, and fatty infiltration. While surgical techniques are promising, the outcomes are variable and often fail to restore full functionality in patients.10,11 Engineered skeletal muscle tissue is a promising treatment for VML because it addresses the issues of tissue availability and donor-site morbidity associated with muscle flaps. Furthermore, our previous studies have shown that our laboratories engineered skeletal muscle units (SMUs) are effective at recovering muscle function following a 30% VML injury in the rat and sheep, respectively.12–17
To study the effectiveness of the SMU to repair VML in a clinically relevant large animal model, we created a 30% VML in the ovine peroneus tertius (PT) muscle and evaluated muscle function after a short 3-month recovery. 16 In this study, we fabricated constructs of clinically relevant size while avoiding the formation of a necrotic core, which is a common problem with avascular engineered tissues. We developed a modular fabrication method in which we combined multiple smaller tissue units through lateral fusion to create skeletal muscle constructs that were 15 cm long and ∼1 cm in diameter. 16 These SMUs were composed of contractile myotubes and linearly aligned extracellular matrix (ECM) proteins and were large enough to repair hindlimb VML in a sheep model. These SMUs also maintained a population of Pax7+ satellite cells that were thought to contribute to muscle repair and regeneration once implanted.
This study showed that the SMU-treated groups restored muscle mass and tetanic force production to a level that was statistically indistinguishable from the uninjured contralateral muscle after only 3 months in vivo. However, the histology showed only a partial recovery of structure in the repair site. It was hypothesized that a longer recovery time may result in further muscle regeneration in the repair site.
In clinical manifestations of VML, there is also often damage to the native nerve innervating the damaged muscle. During surgical interventions, a neurotization is occasionally performed in which a portion of a healthy native nerve is translocated to the injured or transplanted muscle to mitigate the effects of denervation. However, this is not possible when there is a lack of native nerve available to reach the repair site. To address this need, we also developed an engineered neural conduit (ENC) to bridge the spatial gap between healthy native nerve and the repair site. The ENCs are collagen rich and act as a tubular scaffold through which the native nerve can grow. To test the efficacy of ENCs in reinnervating the VML injured site, we dissected 1 cm of a rerouted native nerve and used an ENC to bridge the 1 cm gap to the repair site. Following only 3 months of recovery period, we were able to elicit a muscle contraction through direct stimulation of the rerouted nerve in most animals in the ENC-treated group. 16
Because the effects of VML injuries have been shown to progressively worsen with time, 4 we sought to better understand the effects of a longer term recovery time on the structure and function of the muscle 6 months after having received no treatment (VML-Only group), or having received treatment in the form of an SMU (VML+SMU group) or SMU and ENC (VML+SMU+ENC group).
Methods
Animal care
All animal care procedures were identical to those in our previous study 16 and followed The Guide for Care and Use of Laboratory Animals, 18 according to a protocol approved by the University's Institutional Animal Care and Use Committee. In all instances, animals were first sedated through the administration of intramuscular xylazine (0.2 mg/kg) and then anesthetized through the administration of intravenous propofol (8 mg/kg) and gaseous isoflurane at concentrations between 2–5% to maintain a deep plane of anesthesia. For survival procedures, the animals were fasted, and a fentanyl patch (75 mcg/h) was administered 24 h before surgery. Perioperatively, an intravenous dose of cefazolin (20 mg/kg) was administered. The animals received an intramuscular dose of flunixin (2.2 mg/kg) immediately after surgery with a subsequent dose administered 24 h postoperative as supplementary analgesia. The fentanyl patch was removed 48 h after surgery.
The animals were monitored daily for 10–14 days after surgery by the University of Michigan veterinary staff. As a part of this daily health monitoring, the animals' gait was monitored to see if the animals were favoring the surgical leg or exhibiting “toe-touching,” a sign of pain. Surgical staples were removed 10–14 days after surgery. For terminal procedures, all animals were euthanized through the administration of a lethal dose of sodium pentobarbital (195 mg/kg) and subsequent bilateral pneumothorax.
Muscle biopsy collection
A total of three 4-month-old female Polypay sheep weighing ∼35 kg (Oswalt Farms, Vicksburg, MI) were anesthetized and whole muscle dissections of the semimembranosus muscles were completed under aseptic conditions. After the tissue harvest was complete, animals were subsequently euthanized. The muscles were transported to the laboratory in chilled Dulbecco's phosphate-buffered saline (cat. no. 14190-250; Gibco) supplemented with 2% antibiotic–antimycotic (ABAM) (cat. no. 15240-062; Gibco).
Muscle progenitor cell isolation
Muscle progenitor cells were isolated as described previously.12–17 Briefly, 3.5 g muscle biopsies were sanitized in 70% ethanol and finely minced with a razor blade. The muscle then underwent enzymatic digestion in a solution composed of 2.3 mg/mL dispase and 0.3 mg/mL collagenase type IV at 37°C with constant agitation for 2.5 h. Following enzymatic digestion, the resultant suspension was filtered through a 100 μm mesh filter followed by a 40 μm mesh filter and centrifuged. The supernatant was discarded, and the cells were resuspended in freezing medium (70% Dulbecco's modified Eagle's medium [DMEM]; cat. no. 11995-065; Gibco), 20% horse serum, 10% dimethyl sulfoxide, supplemented with 1% ABAM), frozen to −80°C at a rate of −1°C/min and stored in liquid nitrogen until plating.
Alternatively, a subset of the cell isolates was resuspended in muscle growth media (MGM) [60% F-12 Kaighn's Modification Nutrient Mixture (cat. no. 21127-022; Gibco), 24% DMEM, 15% fetal bovine serum (FBS; cat. no. 10437-028; Gibco), 2.4 ng/mL basic fibroblast growth factor (cat. no. 100-18B; PeproTech, Rocky Hill, NJ), 1% ABAM, and supplemented with an additional 10 μL/mL 1 μM dexamethasone (DEX; cat. no. D4902; Sigma)] and seeded immediately for SMU fabrication.
Modular SMU fabrication
Cells were plated in MGM at a density of 10,000 cells/cm2 onto 500 cm2 tissue culture plates. After seeding, the cells were left undisturbed for 5 days and subsequently fed MGM every 2 days. The media were replaced with muscle differentiation media (MDM) [70% M199 (cat. no. 11150-059; Gibco), 23% DMEM, 6% FBS, 1% ABAM, 10 μL/mL 1 μM DEX, 1 μL/mL insulin–transferrin–selenium-X (cat. no. I1884; Sigma), and 0.72 μL/mL 50 mM ascorbic acid 2-phosphate (cat. no. A8960; Sigma)] when the plates were 100% confluent and elongating myotubes began to form a network (∼7 days postseeding). Light microscopy images of representative monolayers were taken 10 days after initial plating and before monolayer delamination. After 5–7 days on MDM, the monolayers delaminated off the cell culture surface and were transferred to Sylgard-coated plates, which were fabricated as described previously.12–17 Minutien pins were used to pin the monolayer into a three-dimensional (3D) cylindrical construct at lengths of 14 ± 1 cm and ∼1 cm in diameter. Henceforth, this construct is referred to as a “single scaled-up SMU.”
To achieve the desired size to fill the VML defect, a modular approach was used in which 2–4 single scaled-up SMUs were placed side by side and allowed to fuse. These constructs are referred to as “modularly fused SMUs.” It should be noted that after delamination of the monolayer, the SMUs (both single and modularly fused) are not attached to the culture substrate and are suspended in the culture media by the pins. After approximately one additional week in vitro, the modularly fused SMU was ready for either implantation or in vitro characterization. The total fabrication time for one, large construct is approximately three weeks. Details of this fabrication process are described in detail in our previous studies.12–17 An in vitro evaluation of SMUs, including histology and force production, was also described previously.12–17
ENC fabrication
Bone marrow stromal cells (BMSCs) were harvested from an iliac crest marrow aspiration of one adult female sheep, and mononuclear BMSCs were then isolated from the bone marrow as described previously.19–22 Following an established protocol, isolated cells were induced to a tendon lineage and expanded for five passages. The cells were then trypsinized and seeded at a density of 21,000 cells/cm2 onto tissue culture dishes with embedded constraint pins. As a confluent monolayer began to spontaneously delaminate off the cell culture surface, the monolayer was wrapped around a 5-cm-long piece of super soft nylon 11 tubing, creating a lumen (outer diameter 5/32″, cat. no. 1J-262-01; Freelin–Wade). The tubing was adhered to the dish by pinning it to the embedded constraint pins.
A total of four monolayers were wrapped around the tubing and allowed to fuse. The resultant nerve conduits were ∼4 mm in diameter and 7 cm in length. Seven days after the last monolayer was added, media were removed, and ENC was frozen at −80°C to devitalize the cells in the construct and allow for the preservation of the ECM until implantation or in vitro characterization. Just before implantation, the silicone tubing was removed from the ENC. A histological evaluation of ENCs was described previously. 16
Surgical procedures
Animals used for the surgical implant procedures were 7-month-old Polypay Wethers (castrated males) weighing 45–55 kg. The animals were randomly divided into three experimental groups (Fig. 1): VML-Only (n = 15), VML+SMU (n = 15), and VML+SMU+ENC (n = 15). On the day of surgery, the animals were weighed and then placed under a deep plane of anesthesia. A 15 cm incision was made along the front of the lower left leg (surgical side) to expose the PT muscle and peroneal nerve. The experimental and control (no surgery) limbs were not randomized; the surgical leg was always the left leg. Gross measurements of the muscle were taken, and a custom algorithm using a 3D model of the PT muscle was used to calculate the muscle mass constituting 30% of the total muscle volume. Subsequently, a full-thickness longitudinal portion of the PT constituting the calculated muscle mass was dissected. The VML-Only animals (negative control) received the injury without a repair and without the transection of the distal branch of the peroneal nerve; the fascia and skin were closed with suture and the skin was stapled along the incision.
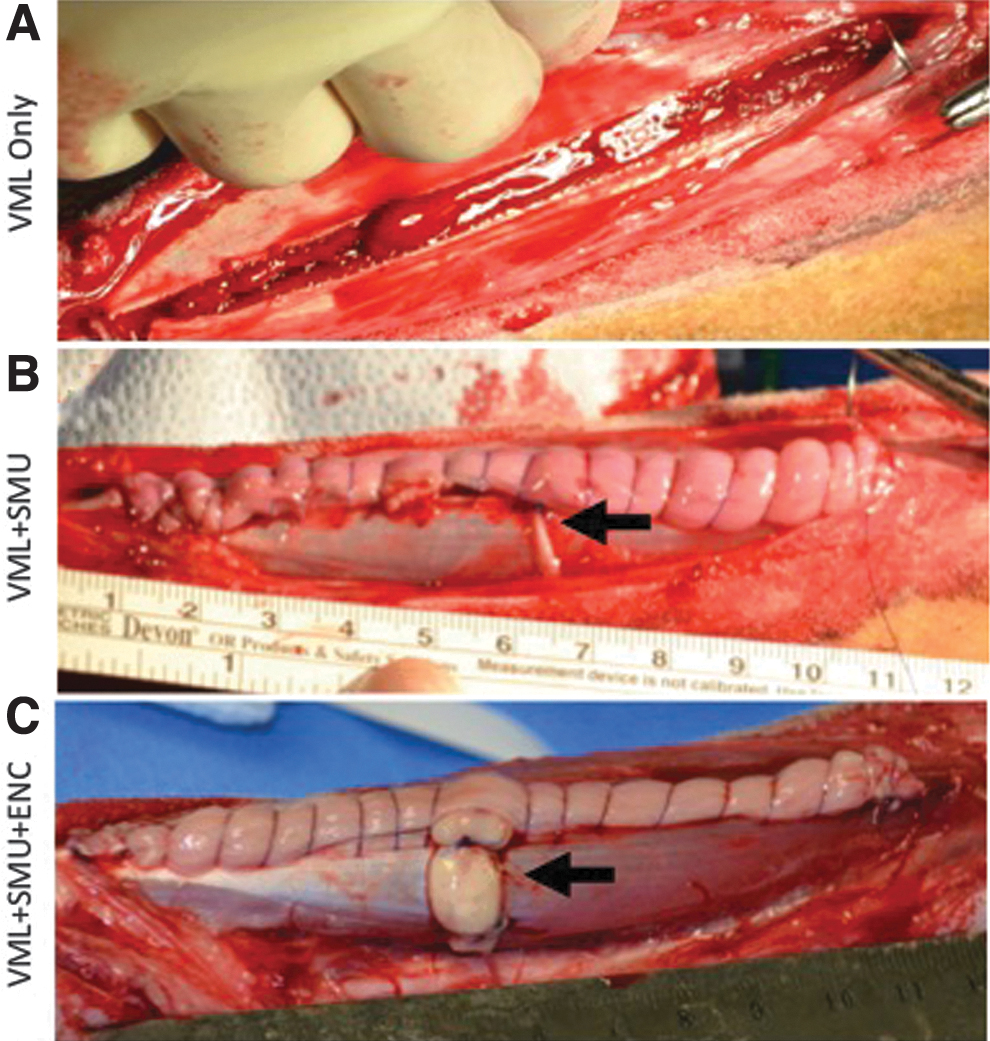
Experimental groups. In all groups, a full-thickness longitudinal portion of the PT constituting 30% of the total muscle volume was dissected.
In the VML+SMU group, the injury was immediately repaired by suturing an SMU with the approximate weight of the muscle removed within the defect. Additionally, the distal branch of the peroneal nerve that innervates the medial digit was transected and rerouted to the SMU to aid the process of reinnervation. In the VML+SMU+ENC group, the VML injury was also repaired with an SMU; however, in this group, 1 cm of the rerouted peroneal nerve was dissected to simulate a nerve injury, which often accompanies VML. The gap between the SMU and the peroneal nerve was then bridged with an ENC. A 4-0 Prolene was used to suture the rerouted nerve and ENC, whereas 4-0 PDS II was used to suture the SMU, fascia, and skin. All animals were upright and weight bearing immediately following recovery from the gas anesthesia. The repair of the VML defect was treated the same in all surgical groups with respect to wound care and closure. All animals were monitored daily for 14 days after surgery and then returned to herd housing. The animals recovered for 3 months before we assessed functional recovery and performed muscle histological analyses.
In situ biomechanical testing
Following the 6-month recovery, animals were weighed and then anesthetized. Both the contralateral and surgical PTs were dissected leaving the proximal origin intact. To secure the knee, a metal rod was inserted through the femoral epicondyles and secured to a rig mounted to the surgical table. Additionally, adjacent nerves and musculature were severed so as not to interfere with testing. The distal tendon of the PT was secured to a custom strain gauge force transducer (Vishay Precision Group, Malvern, PA) to measure the force of the muscle contractions. Contractions were elicited by stimulating the nerve innervating the PT (i.e., a branch of the deep peroneal nerve) with a bipolar platinum wire electrode. Biomechanical testing of the muscles was conducted as described previously. 16 Briefly, the muscle was placed in the slack position and single 0.1 ms pulses of increasing current amplitudes (i.e., 60, 70, 80, 90, 100, 110, and 120 mA) were delivered until peak twitch force was reached.
Maintaining the current, the muscle length was subsequently adjusted to the length at which twitch force was maximal. The length of the muscle at which twitch force was maximal was defined as the optimal length (Lo). The stimulus was then switched to a tetanus in which a 600 ms train of 0.1 ms pulses were delivered. The frequency of these pulses was increased (i.e., 60, 80, 100, and 120 Hz) until isometric tetanic force was maximal. Data were recorded using custom LabVIEW 2018 software. In the repairs with the rerouted nerve, the nerve was dissected and tested as described above for the native nerve. This process was then repeated on the contralateral (uninjured) PT muscle. Immediately after biomechanical testing, both the contralateral and surgical PTs were fully dissected, weighed, and prepared for histology. The animals were subsequently euthanized.
Histology, fiber typing, and collagen content
After dissection, the muscles were weighed, and gross measurements were taken. The muscles were then divided into segments and prepared for histology in one of two ways; the sample was either coated in Tissue Freezing Medium and frozen in dry ice-chilled isopentane or fixed in 10% formalin for 24 h and then embedded in paraffin. Frozen samples were cryosectioned at 10 μm and then immunohistochemically stained to identify myosin heavy chain (5 μg/mL dilution, cat. no. MF-20c; DSHB), laminin (5 μg/mL dilution, cat. no. ab7463; Abcam), fast myosin isoform (1:200 dilution, cat. no. ab91506; Abcam), and slow myosin isoform (1:200 dilution, cat. no. ab11083; Abcam) using a protocol described previously. 16 Longitudinal samples were sectioned at 25 μm and immunohistochemically stained for acetylcholine receptors (α-bungarotoxin, 1:2000 dilution, cat. no. B1601; Life Technologies), synaptic vesicle protein-2 (2 μg/mL dilution, cat. no. SV2c; DSHB), and neurofilament (0.5 μg/mL dilution, cat. no. 837904; BioLegend) to identify the presence of neuromuscular junctions.
To evaluate collagen organization, sections stained with Picrosirius Red were imaged under monochromatic polarized light. The birefringence of collagen fibers was semiquantitatively analyzed, based on brightness, to determine differences in collagen deposition and maturation at the site of repair.22,23 Two representative longitudinal sections across the full width of the muscle through the mid-belly of the VML repair sites were analyzed for N = 5 animals per group. The polarization plane was rotated until maximum brightness was obtained to account for variations in specimen orientation on the slide. Digital images were taken of each tissue section at X10 magnification during a single imaging session under identical imaging parameters. The images were imported into ImageJ software and underwent 8-bit digitalization.
Noncollagenous material was shown as black (zero) and collagenous material was assigned a gray scale value from 1 to 255. Higher gray scale values signify more organized and mature collagen. Mean gray scale values for each image were averaged for each animal that obtained a brightness value. Due to the increased level of collagen in repairs compared with control muscle, we normalized the gray scale signal to the total amount of collagen present to more accurately the gray scale signal due primarily to collagen alignment.
To evaluate the fiber types in the muscle, a mid-belly cross-section of each PT was immunohistochemically stained for fast myosin isoform to identify the type II muscle fibers and slow myosin isoform to identify type I fibers. To evaluate the percentage of the total muscle cross-sectional area that was type I fibers, the amount of area positively stained for slow myosin isoform was measured using ImageJ/Fiji. These data were presented as the total number of slow fibers as a percentage of the total number of fibers enumerated.
To evaluate the degree of fiber type grouping, we also enumerated the number of grouped fibers in the entire muscle cross-section. A “grouped fiber” is defined as a type I muscle fiber that is completely surrounded by other type I muscle fibers. These data were correlated to the percentage of force recovery for each animal. Due to the novel findings in the 6-month group, we went back and analyzed the 3-month animals. Thus, the analysis includes data from the 3- and 6-month recovery group. While some of the data collected on the 3-month animals have already been published, 16 new slides were produced from frozen sections collected from the 3-month recovery animals for this analysis.
Paraffin-embedded cross-sectional samples were sectioned at 5 μm and stained with hematoxylin and eosin (H&E), Picrosirius Red, and Masson's Trichrome to visualize morphology of the repair site, monitor collagen alignment, and content. To quantitatively evaluate the collagen content of the muscles, a cross-section from the mid-belly at the widest portion of the contralateral and surgical PT muscles was stained with Picrosirius Red. These samples were imaged and the percentage of positive red staining relative to the total cross-sectional area (percent collagen) was measured using ImageJ/Fiji (NIH).
Statistical analyses
For the collagen alignment, statistical analysis of specimen brightness was performed by two-way analysis of variance (ANOVA) with p-value <0.05 considered significant. Data are presented as mean ± standard error. Statistical analyses were performed using GraphPad Prism 7 software. Statistical differences between groups in which the contralateral muscle was compared with the surgical PT muscle were assessed with a two-way ANOVA and Sidak's multiple comparison (Sidak's MC) tests. Statistical differences between groups in which the values were normalized to the contralateral side were assessed with a one-way ANOVA with Tukey's multiple comparison (Tukey's MC) test. Results were significant at p < 0.05. Bars on graphs indicate mean ± standard deviation.
Results
Surgical procedures
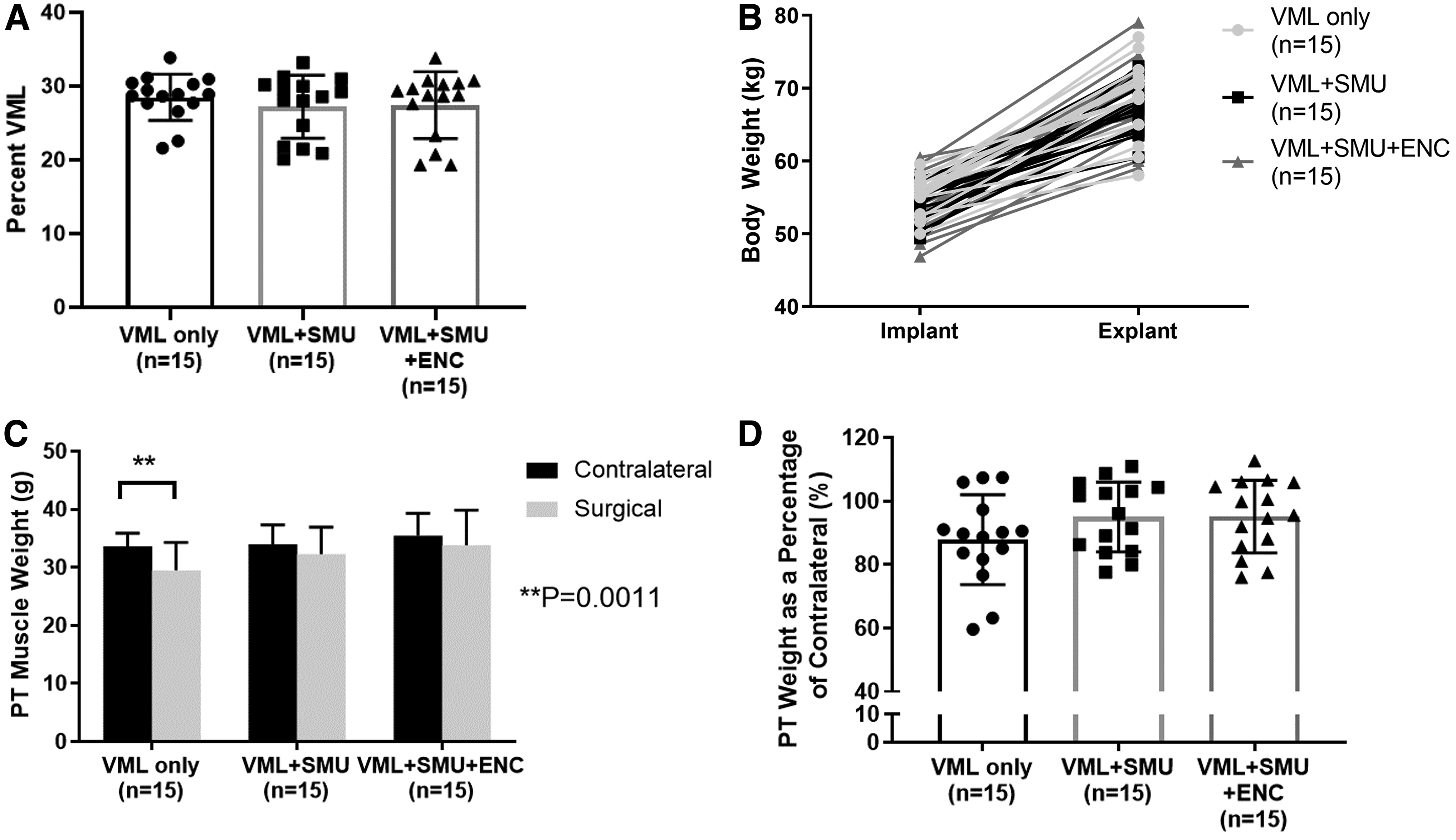
Across all VML injury groups, the calculated average percentage of muscle removed was 27.7 ± 3.97% and the actual mass removed was 5.32 ± 0.69 g. The variability in the weights removed is due to differences in size of the PT muscles between individuals. There was no significant difference in the magnitude of the VML injury between groups (one-way ANOVA: p = 0.6502, n = 15 per group). Specifically, the VML injury was calculated to be equal to 28.5 ± 3.1% in the VML-Only group, 27.2 ± 4.3% in the VML+SMU group, and 27.4 ± 4.5% in the VML+SMU+ENC group (Fig. 2A).
VML injury and mass recovery.
Body mass and muscle mass recovery
Animals were awake, alert, and weight bearing within 2 h of completing surgery. None of the animals was observed to have abnormal gait for extended periods of time after surgery; however, about one-third of animals exhibited occasional “toe-touching,” a sign of pain, while n = 4 animals were noted to have a slight limp for 1 day during the recovery period. Additionally, animals experienced mild-to-moderate swelling at the injury site for up to 2 weeks postoperatively. One hundred percent of animals gained weight normally during the 6-month recovery period and there was no significant difference in the animals' body weight between experimental groups (two-way repeated measures [RM] ANOVA: experimental group: p = 0.5461, n = 45) (Fig. 2B). This indicates that the VML injury did not significantly impact the health of the animals during this 6-month recovery period. Additionally, no signs of rejection were observed in the sheep. Bloodwork taken at the time of explant revealed no signs of a chronic immune response (i.e., white blood cell counts did not exceed normal limits).
Gross observations at the time of explant revealed larger amounts of connective tissue in the VML injured PTs of all animals relative to what was observed in the uninjured contralateral. We did not note muscle tethering to surrounding tissues. At the time of explant, we compared the weights of the VML injured PTs with the contralateral sides. Notably, there were no significant differences in muscle mass between the contralateral and VML repaired PTs in the VML+SMU and the VML+SMU+ENC groups (Sidak's MC: p = 0.3064 and p = 0.3616, respectively, n = 15 per group) (Fig. 2C). In contrast, there was a significant muscle mass deficit in the VML-Only group (Sidak's MC: p = 0.0011, n = 15) indicating a lack of muscle mass regeneration.
The significant mass deficit experienced by the VML-Only group was expected, as the defect in the VML-Only group was not repaired with a construct or any other filler. We also compared groups after normalizing the weight of each VML injured PT muscle to the contralateral muscle. Represented as a percentage of the contralateral, there were no significant differences in PT weight between groups (one-way ANOVA: p = 0.1929) (Fig. 2D). On average, the VML-Only, VML+SMU, and VML+SMU+ENC groups had an 87.87 ± 14.16%, 95.01 ± 11.04%, and 95.13 ± 11.46% muscle mass recovery, respectively.
Force recovery
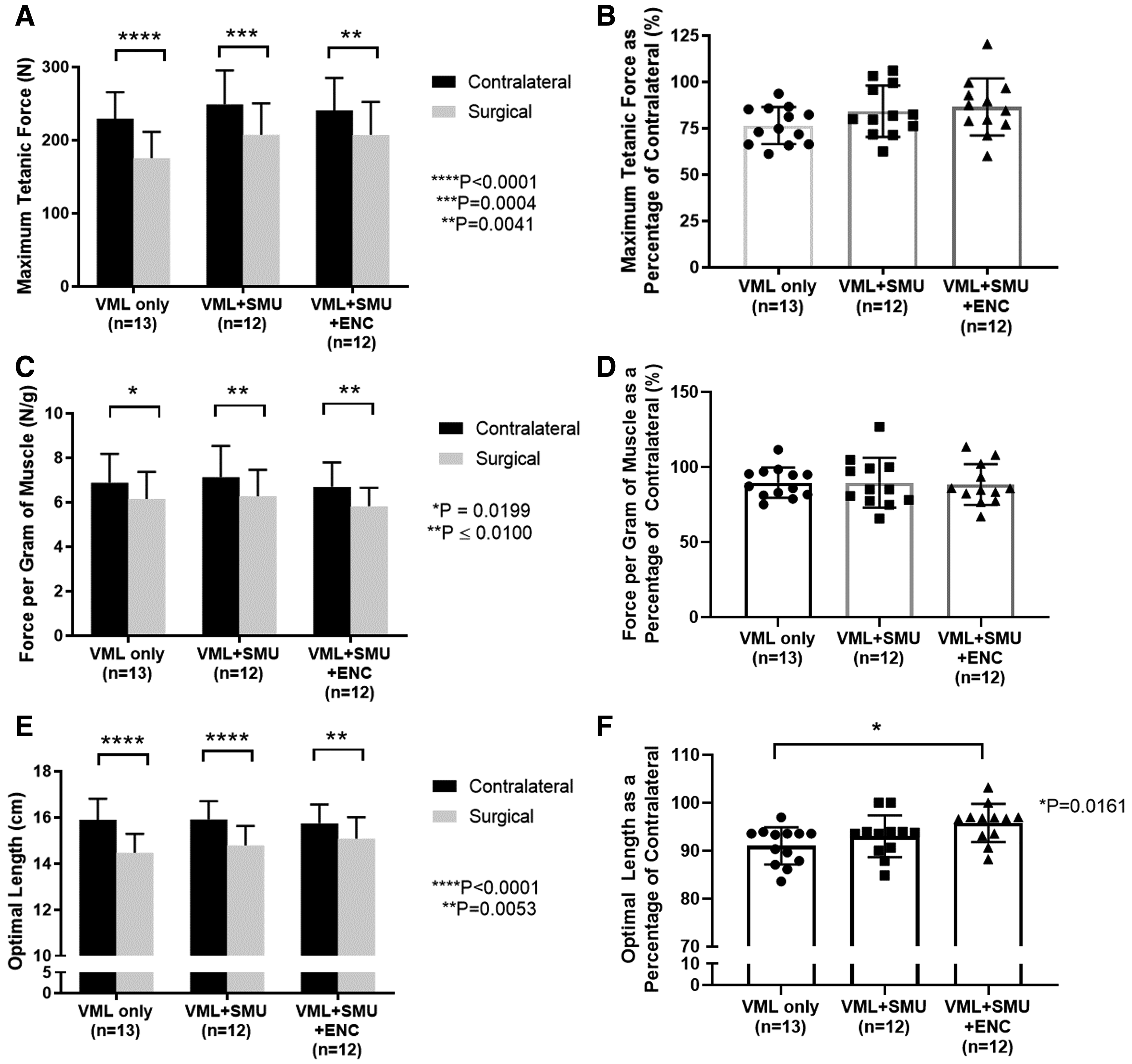
To assess force recovery, we compared the maximum tetanic force of the VML injured PT with that of the contralateral PT (Fig. 3A). All VML injured muscles experienced a significant decrease in maximum force production between the injured and contralateral PTs [Sidak's MC: p < 0.0001 for VML-Only (n = 13), p = 0.0004 for VML+SMU (n = 12), and p = 0.0041 for VML+SMU+ENC (n = 12)]. Muscle weight (g), absolute values for maximum (N), and specific force (N/g) are presented in Table 1. Specifically, the mean difference in maximum tetanic force between the contralateral and VML injured PTs in the VML-Only, VML+SMU, and VML+SMU+ENC groups was 54.2 ± 9.2 N, 41.4 ± 9.6 N, and 33.4 ± 9.6 N, respectively. Although the differences between groups were not significant (two-way RM ANOVA: p = 0.2138), the force recovery in the VML+SMU+ENC group was the greatest on average. Represented as a percentage of the contralateral, there were no significant differences between VML groups (one-way ANOVA: p = 0.1497) (Fig. 3B).
Biomechanical evaluations. Tetanic isometric forces elicited by an electrical stimulus were measured from both the surgical VML injured and contralateral PT muscles.
Muscle Weight and Muscle Force Characteristics
Values are means ± SD.
Significantly different from Control p < 0.05.
Significantly different from Control p < 0.001.
Significantly different from Control p < 0.0001.
ENC, engineered neural conduit; SD, standard deviation; SMU, skeletal muscle unit; VML, volumetric muscle loss.
On average, the VML-Only, VML+SMU, and VML+SMU+ENC groups experienced a 76.5 ± 10.0%, 84.2 ± 13.9%, and 86.6 ± 15.4% maximum tetanus force recovery, respectively. Additionally, data show no significant difference in the frequency required to produce a maximum tetanus between groups. For the uninjured contralateral, maximum tetanus was achieved at 79.5 ± 14.1 Hz and the frequency was 86.9 ± 18.0 Hz, 91.7 ± 16.4 Hz, and 87.5 ± 23.0 Hz for the VML-Only, VML+SMU, and VML+SMU+ENC groups, respectively. Statistically, there is no significant difference between these experimental groups (one-way ANOVA: p = 0.1178).
Because of the complex muscle architecture of the PT muscle, we were not able to calculate specific force in the traditional sense, by dividing the maximum force by the physiological cross-sectional area. Instead, we normalized the maximum force to the muscle weight (Fig. 3C, D). All VML injured groups experienced a significant decrease in maximum force production normalized to muscle weight between the injured and contralateral PTs [Sidak's MC: p = 0.0199 for VML-Only (n = 13), p = 0.0081 for VML+SMU (n = 12), and p = 0.0077 for VML+SMU+ENC (n = 12)] (Fig. 3C). Specifically, the mean difference in force per gram of muscle between the contralateral and VML injured PTs in the VML-Only, VML+SMU, and VML+SMU+ENC groups was 0.74 ± 0.26 N/g, 0.86 ± 0.27 N/g, and 0.87 ± 0.27 N/g, respectively. Represented as a percentage of the contralateral, there were no significant differences in the force per gram of muscle between groups (one-way ANOVA: p = 0.9695) (Fig. 3D). On average, the VML-Only, VML+SMU, and VML+SMU+ENC groups experienced an 89.6 ± 10.10%, 89.6 ± 16.7%, and 88.4 ± 13.6% normalized specific force recovery, respectively.
Although we did not notice any tissue tethering, we did observe changes in the Lo of the muscle when comparing the VML injured to the contralateral PT. A Sidak's multiple comparisons test revealed that there was a significant decrease in the Lo in all VML injured muscles compared with contralateral muscles (p < 0.005, Fig. 3E). The Lo represented as a percentage of the contralateral was significantly lower in the VML-Only group compared with the VML+SMU+ENC group (TMC: p = 0.0161, n = 13 for VML-Only, n = 12 for VML+SMU+ENC) (Fig. 3F). Specifically, average percent Lo was 91.0 ± 3.9%, 93.0 ± 4.4%, and 95.8 ± 4.0% for the VML-Only, VML+SMU, and VML+SMU+ENC groups, respectively. In addition to stimulating the native nerve innervating the PT, we also sought to elicit a muscle contraction by stimulating the nerve that was transected and rerouted to the surgical site. Direct stimulation of the rerouted nerve resulted in a muscle contraction in 73% (11 out of 15) of the animals in the VML+SMU group and 93% (14 out of 15) of the animals in the VML+SMU+ENC group.
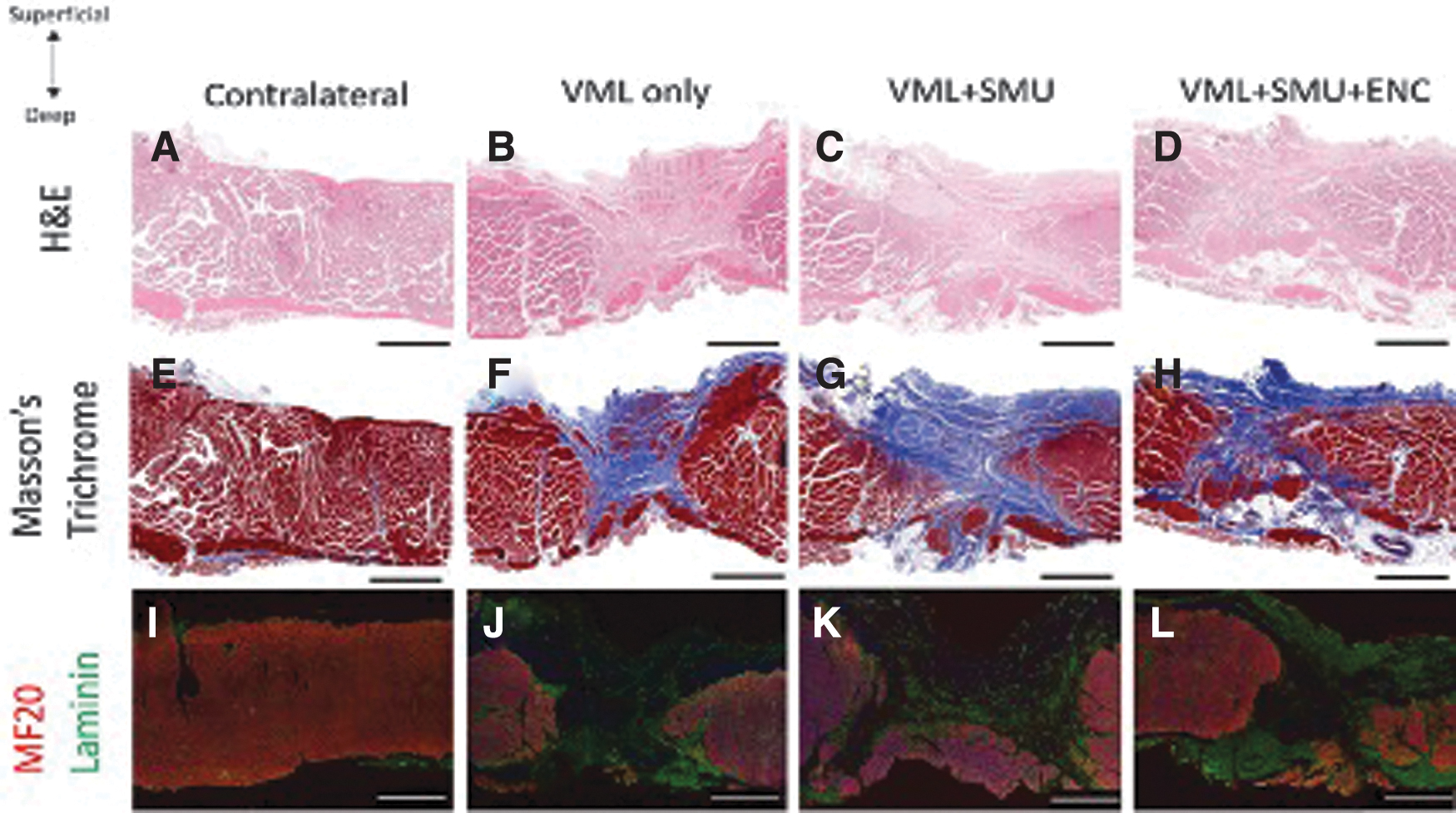
Histological analyses of muscles
We performed qualitative and quantitative histological analyses on cross-sections of the mid-belly of the PT in both contralateral and VML injured PT muscles. In all surgical groups, the repair site was characterized by a fibrotic region, as evidenced by the hematoxylin and eosin staining (Fig. 4A–D) and Masson's Trichrome staining (Fig. 4E–H). The blue regions in the sections stained with Masson's Trichrome demonstrate the collagen deposition in the repair site. Immunostaining for myosin heavy chain and laminin (Fig. 4I–L) showed that there was also laminin in the repair site.
Gross morphology of explanted muscle. Cross-sections of explants from contralateral
At higher magnifications, we noticed the presence of vasculature in the repair site of the hematoxylin and eosin-stained sections of all VML injured groups (Fig. 5A–D). We also observed the presence of intramuscular fat near the repair site of the VML injured groups, but not in the uninjured contralateral muscle (Fig. 5A–D). Immunohistochemical staining for myosin heavy chain (MF20) and laminin revealed the presence of small muscle fibers within the repair site in the VML-Only, VML+SMU, and VML+SMU+ENC groups (Fig. 5E–H). The presence of these small muscle fibers suggests that following VML, muscle regeneration within the wound site is occurring; however, the origin of these fibers has not been determined. We also noted the presence of neuromuscular junction formation in all VML injured groups as evidenced by positive staining for acetylcholine receptors, synaptic vesicle protein-2, and neurofilament in longitudinal sections of the VML injury site (Fig. 5I–L).

Microstructures of Explanted Muscle. H&E-stained cross-sections of the repair site demonstrated the presence of fat (red circles) in all VML injured groups
To quantitatively evaluate the collagen content of the muscles, we measured the percentage of positive Picrosirius Red staining (percent collagen) in mid-belly cross-sections of the PT muscles (Fig. 6). We first verified this method by evaluating the collagen content of the uninjured contralateral muscles and found that they have very little variability in the percent collagen. Specifically, the average percent collagen in the uninjured contralateral PTs was 17.63 ± 2.76% (n = 13). In contrast, the average percent collagen in the VML injured PTs was 46.0 ± 8.5%, 36.5 ± 4.5%, and 39.6 ± 9.2% for the VML-Only (n = 13), VML+SMU (n = 11), and VML+SMU+ENC (n = 12) groups, respectively. The percent collagen of the VML injured PT of the VML-Only group was significantly higher than the VML+SMU+ENC group (Sidak's MC: p < 0.0377), which indicates that there was significantly more connective tissue deposition in the VML-Only group compared with the VML+SMU+ENC groups (Fig. 6E).
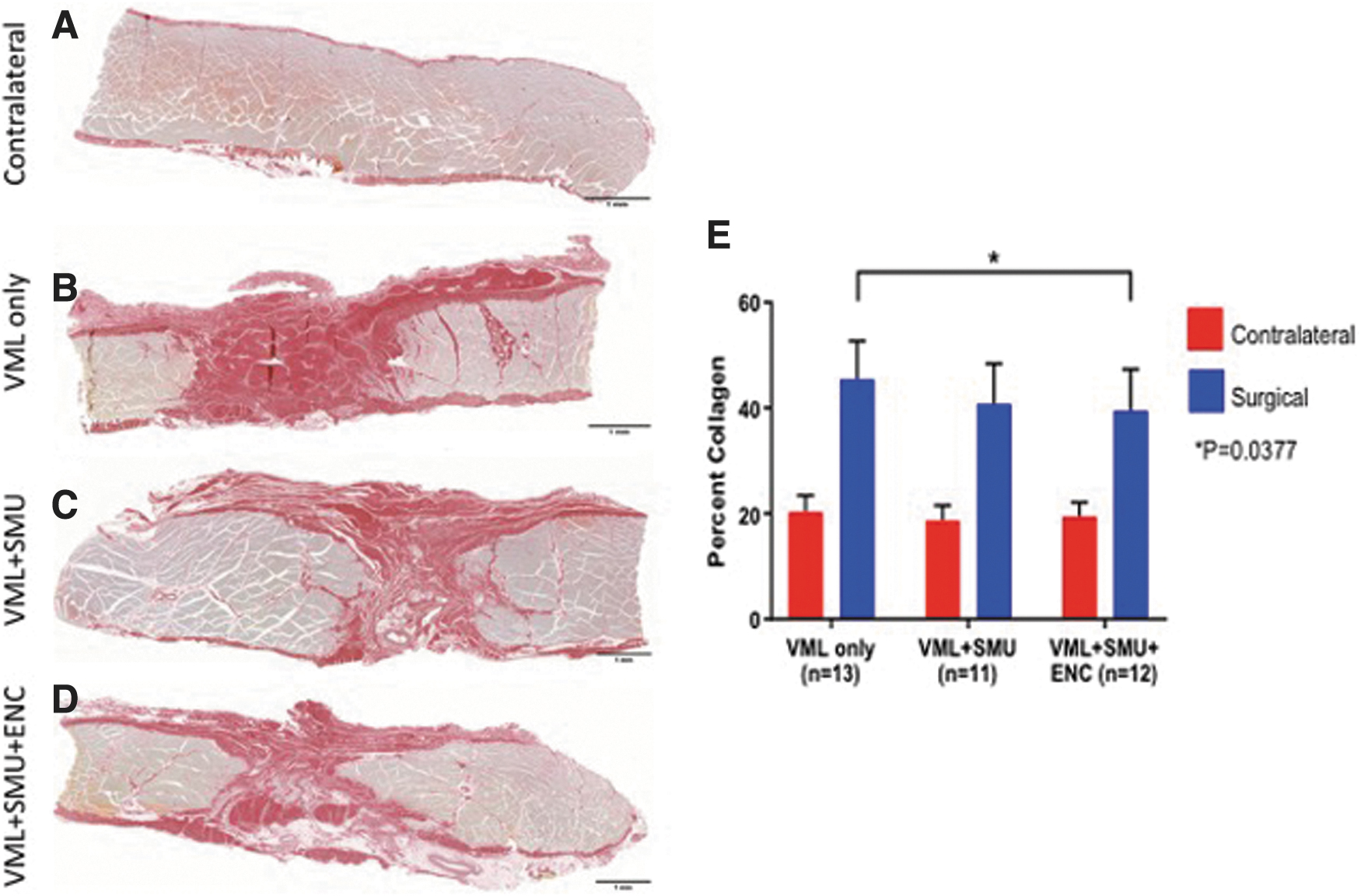
Analysis of collagen content in explanted muscle. Mid-belly PT cross-sections of the VML injury sites of muscles in the
We also performed quantitative histological analyses of the collagen content and collagen alignment on longitudinal sections of the mid-belly of the PT in both contralateral and VML injured PT muscles (Fig. 7). In all surgical groups, the repair site was characterized by an increase in collagen content compared with control, as evidenced by the Picrosirius staining and polarized light analysis (Fig. 7A–D). Grayscale normalized to the total amount of collagen indicated that the control muscle had significantly less collagen, but this collagen was highly aligned (Fig. 7E). The VML-Only group had the greatest collagen content and was the least aligned (Fig. 7E). Normalization of the collagen alignment by the percentage of collagen content indicated that the VML+SMU group was not significantly different from control or the VML+SMU+ENC groups and was significantly different (p < 0.01) from the VML-Only group. The VML+SMU+ENC was significantly different from both the control and VML-Only groups, p > 0.01 and 0.05,
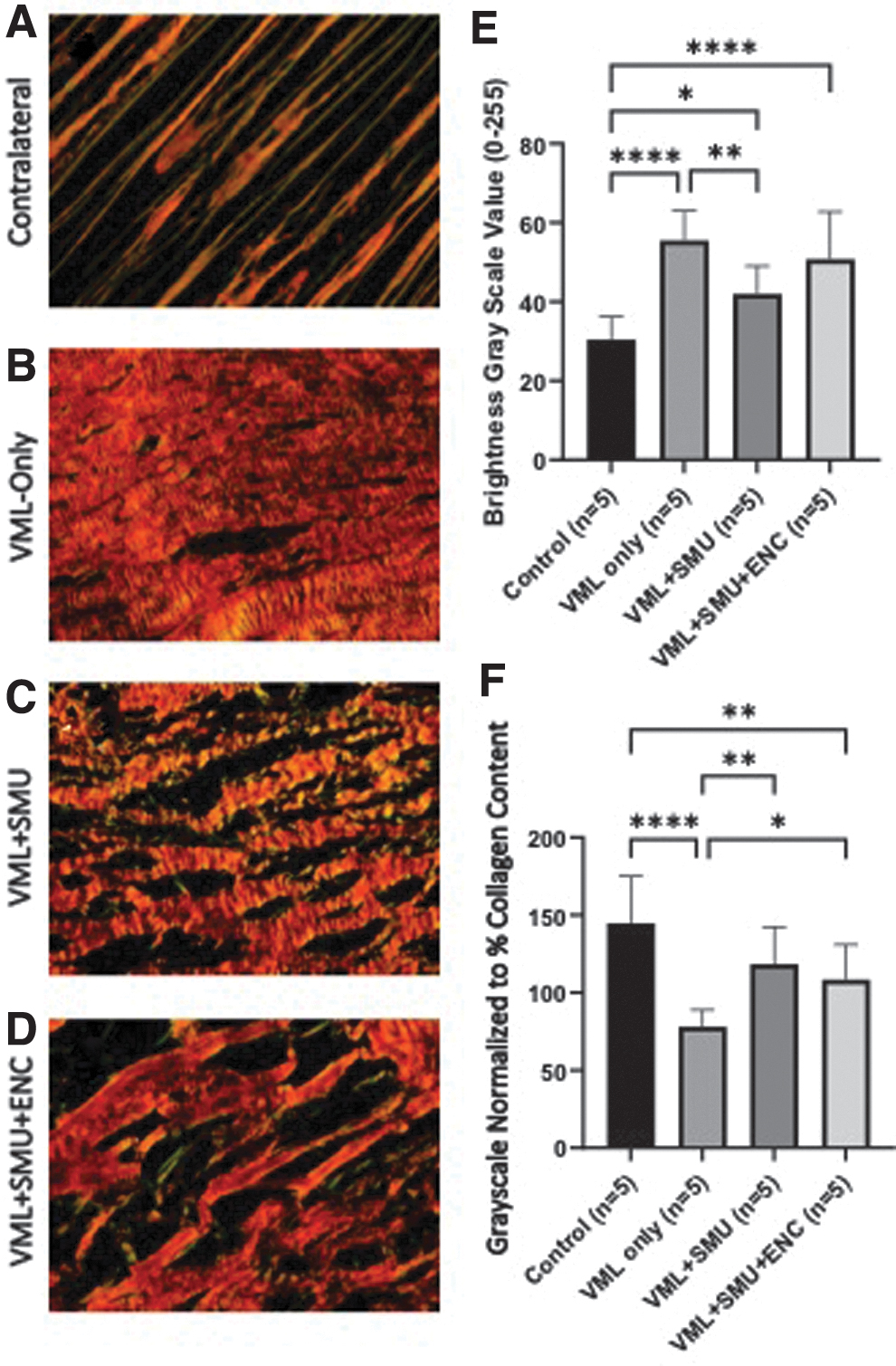
Analysis of collagen alignment and content in longitudinal sections of explanted muscle. Mid-belly PT longitudinal sections of the VML injury sites of muscles in the
Due to the findings of increased fiber remodeling in the 6-month muscle repair sites, we did further histological analyses of the 3-month recovery groups. The remaining paragraphs in this section describe the differences in reinnervation and muscle remodeling between the 3- and 6-month recovery time points. We determined the percentage of slow fibers constituting the total area of the muscle to give an indication of fiber type change over time (3 vs. 6 months). Images presented in Figure 8 are representative images from the 6-month recovery time point for (A) contralateral, (B) VML-Only, (C) VML+SMU groups, and (D) VML+SMU groups. The percentage of slow fibers was not significantly different between time points or VML injured groups (p = 0.6196, n = 5 per group) (Fig. 8E).

Fiber type analysis at the 3- and 6-month recovery time point. To note any changes to the fiber type of the muscle at the 3- and 6-month recovery time point, we performed immunostaining for fast myosin isoform (type II fibers, green) and slow myosin isoform (type I fibers, red). Presented are representative images from the 6-month recovery time point for:
In the 3-month recovery group, the cross-sectional area represented by slow fibers was 25.25 ± 12.17% in the VML-Only group, 23.39 ± 5.26% in the VML+SMU group, and 18.6 3 ± 7.07% in the VML+SMU+ENC group, compared with the contralateral, which was 18.08 ± 4.49%. Following the 6-month recovery the cross-sectional area represented by slow fibers was 17.53 ± 8.48% in the VML-Only group, 18.50 ± 10.19% in the VML+SMU group, and 19.28 ± 7.92% in the VML+SMU+ENC group, compared with contralateral, 17.76 ± 5.45%.
Upon closer inspection of fiber type distributions, we noted an increase in the number of muscles with slow (type I) fiber grouping at 6 months when compared with the 3-month recovery time point. Images presented in Figure 9 are representative images from the 6-month recovery time point for (A) contralateral, (B) VML-Only, (C) VML+SMU groups, and (D) VML+SMU groups. With the exception of one muscle from the 6-month VML-Only group, there were fewer than 49 grouped fibers in all control and VML-Only at the 3- and 6-month recovery time points, respectively. Similarly, 4 out of 6 VML+SMU and the VML+SMU+ENC muscles had fewer than 49 grouped fibers at the 3-month recovery time point. However, in the 6-month group, the VML+SMU and VML+SMU+ENC groups, which received a rerouted donor nerve into the repair site, showed an increase in slow (type 1) fiber type grouping compared with the control muscles (Fig. 9E).
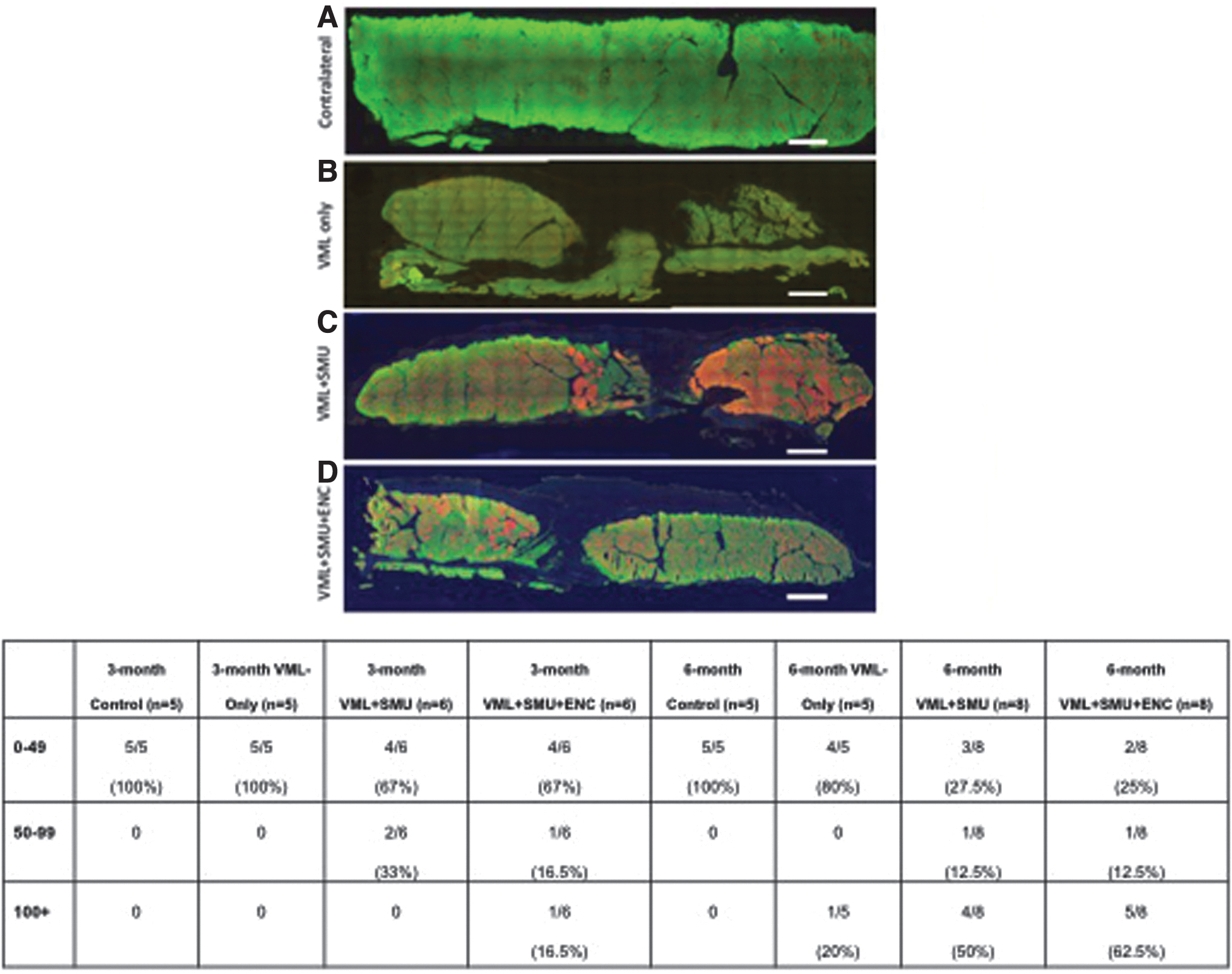
Percentage of animals with slow (type 1) fiber grouping at 3 and 6 months. To note any changes to the fiber type of the muscle at the 3- and 6-month recovery time point, we performed immunostaining for fast myosin isoform (type II fibers, green) and slow myosin isoform (type I fibers, red). Presented are representative images from the 6-month recovery time point for:
Specifically, there were more than 100 grouped fibers in 4 out of 8 of the VML+SMU and 5 out of 8 of the VML+SMU+ENC muscles at the 6-month recovery time point (Fig. 9E). The greater number of animals exhibiting fiber type grouping in the VML+SMU+ENC group compared with the VML+SMU or VML-Only group, suggests the rerouted nerve in combination with the ENC enhanced the remodeling/reinnervation of the muscle.
Due to the increase in the number of animals with slow (type 1) fiber grouping and the effect that this could have on force production, we evaluated the correlation between slow-type fiber grouping and percentage of maximum tetanic force recovery at 3- and 6 months. Notably, there was no significant correlation between the number of grouped slow fibers and the percentage of maximum tetanic force recovery in either the 3- or 6-month groups (Fig. 10).
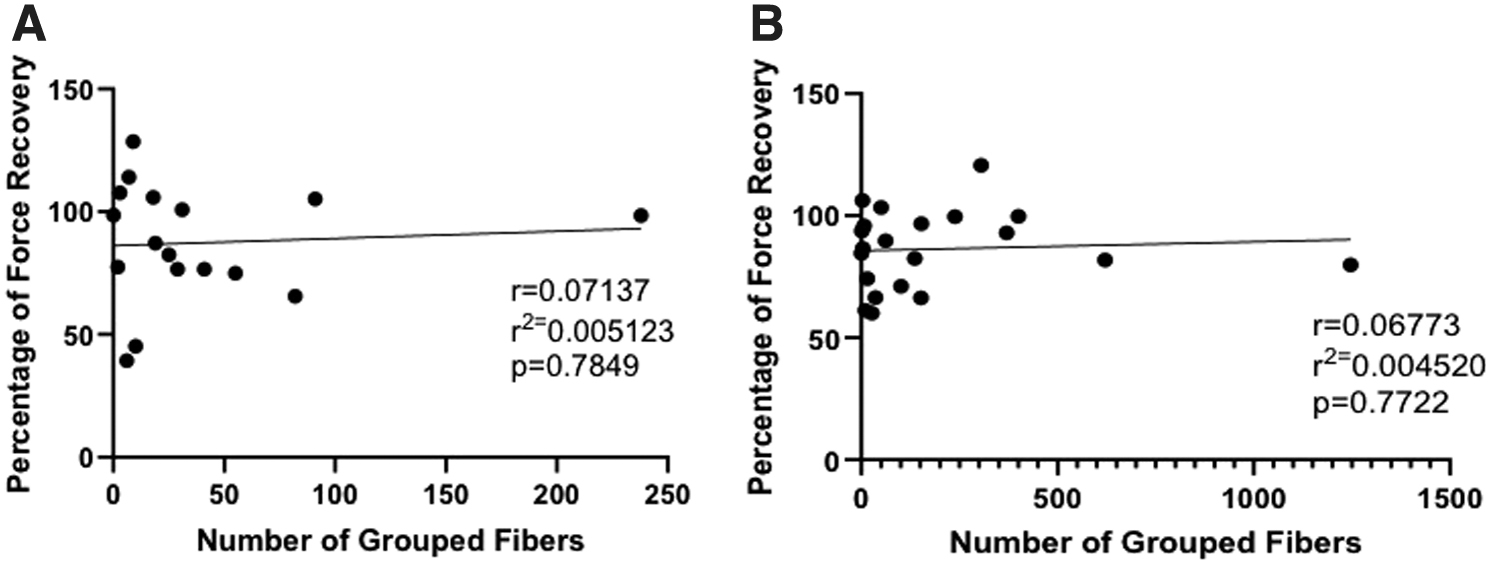
Fiber grouping and force in the 3- and 6-month recovery group. There was no significant correlation between the number of grouped fibers and the percentage of force recovery in either
Discussion
VML injuries are defined by a traumatic permanent loss of skeletal muscle, extensive fibrosis, and functional impairment, which can often lead to chronic disability.4,11,24 The current standard of care for repairing VML injury is to replace lost muscle by translocating autogenic tissue from a donor site into the wound. 10 Other treatment techniques use regenerative medicine strategies to address the limitations of current treatment options and promote functional healing.15,16,25 These strategies include the delivery of myogenic cells to the wound site or the implantation of acellular scaffold approaches.8,26–29 While these strategies show some regeneration of muscle function, no intervention to date restores muscle function to preinjury levels following VML. The permanent loss of muscle function observed following VML injury can be partially explained by an acute fibrogenic and inflammatory response, which invades the injury site and impedes regeneration of functional muscle.30,31
The acute fibrotic infiltration into the wound site not only diminishes reparative myogenesis, but also interferes with innervation and vascularization of the remaining healthy muscle and newly regenerating muscle fibers and contributes to failure in restoring muscle function following treatment.32,33 In fact, patients suffering from VML often have complex traumatic injuries that include bone and muscle damage. The goal of the surgeon is first to repair bones and avoid infections. The repair of muscle loss often occurs weeks or months after the injury. 34 In this study, we used an acute VML with immediate repair with an SMU graft. This acute model allows the study of novel technologies in a more controlled, best-case scenario. Technologies showing success in an acute model, it will have to be vetted in a chronic model of VML. A technology that successfully promotes muscle regeneration in the face of acute inflammation and fibrogenesis would be a significant advancement in the repair of acute VML injuries. Our SMUs offer a novel skeletal muscle engineering approach to repair VML injury.
In a previous study, our laboratory was able to scale-up and fabricate engineered muscles (SMUs) that were 14–15 cm long and
We hypothesized that: (1) a longer recovery time would result in improved muscle regeneration; (2) with increased time, we would observe remodeling and removal of amorphous fibrotic tissue present at 3 months and that we would also observe increases in sarcomere organization, revascularization, and reinnervation of the wound site; and (3) that ENCs would enhance nerve regeneration and reinnervation in the wound site and that would contribute to the recovery of muscle force production. Thus, the purpose of this study was to evaluate the potential of our scaffold-less tissue-engineered SMU constructs to treat a volumetric muscle injury in a sheep model following a 6-month recovery. To some degree each of the above hypothesis were supported by the data presented.
One of the limitations to many of the current interventions for VML injury is the potential for immune rejection of the implanted tissue. As previously described in the 3-month study, 16 despite the allogeneic nature of the SMU and ENC grafts, we did not observe an immune response in the animals of this study. In concurrence with the 3-month study, we observed extensive fibrosis in the wound site of the VML injured muscle. Additionally, amorphous collagen fibers surrounded the regenerating muscle fibers, although, to a lesser extent than at 3 months, which suggests continued remodeling and repair of the injury site between 3 and 6 months of recovery. At 6 months, the VML-Only group had a significantly greater percentage of collagen within VML injury site than the SMU repaired groups. The collagen fibers in the repair site were significantly more aligned in the SMU repaired groups than the VML-Only (no repairs) and look qualitatively more like control muscle following repair with an SMU.
Additionally, we observed an increase in muscle fiber content in the VML injury site following a 6-month recovery in the SMU repaired groups. These data suggest that the muscle regeneration and wound site remodeling was improved by the addition of the SMU when compared with the VML-Only group compared with the SMU-repaired groups and that the additional 3 months of recovery allowed for further muscle regeneration in the groups repaired with an SMU.
In accordance with our 3-month study, the results of the 6-month time point demonstrated the ability of the SMUs to restore muscle mass in the VML injury site to a level that was statistically indistinguishable from the uninjured contralateral muscle. However, unlike the 3-month study, the maximum and specific force production was significantly lower in the VML injured muscle at 6 months. Following a 6-month recovery, the VML-Only, VML+SMU, and VML+SMU+ENC groups sustained deficit in maximum tetanus force generation of a 23.5%, 15.8%, and 13.4% sustained deficit in maximum tetanus force generation, respectively. The reduction in Lo suggests a change in the tissue's mechanical properties and the gross architecture of the muscle. 35 As a reduction in Lo is expected to occur in VML, 9 the fact that the VML+SMU+ENC group's Lo as a percentage of the contralateral was significantly higher compared with the VML-Only group, suggests that our constructs may have mitigated the effects of the injury by attenuating changes to the tissue's mechanical properties. While the VML injuries repaired with an SMU appeared to have greater regeneration of muscle force than the VML-Only group, this did not reach statistical significance.
There are several explanations for the lack of continued recovery in the SMU repaired muscles over time, including lack of continued reinnervation to the VML site and/or maladaptive changes to the remaining healthy muscle because of a transient denervation. Fiber-type shifts occur in the muscles during recovery from VML. As such, we analyzed the fiber type and fiber grouping in the repaired muscles following the VML injury. A shift in fiber type from fast to slow and an increase in the grouping of slow fibers is an indication of denervation and reinnervation of the muscle tissue. This shift to higher percentage of slow fibers could lead to a decrease in maximum force production. 36 One study suggests that donor-specific conversions to a slow muscle fiber type and an increase in slow fiber grouping occurs following denervation and reinnervation with a grafted donor nerve. 16 While we did not see a significant increase in slow fibers, there was a trend toward an increase in slow fibers that could have a contributing factor to the observed decrease in force capabilities following VML on our model.
The analysis of fiber type showed that greater number of VML repaired muscles exhibited fiber grouping at the 6-month time point compared with the 3-month time point, which suggests that muscle remodeling continued to occur during the additional 3 months of recovery. Specifically, there was an increase in slow fiber-type grouping in the VML+SMU+ENC groups compared with the VML-Only group. Thus, the possibility exists that the remaining healthy muscle was also remodeled to have a greater percentage of slow fibers either due to denervation and increased reinnervation by slow motor neurons originating from either the native nerve or the rerouted nerve. However, this does not explain the observed decrease in specific force. It may be that there is a decreased ability to recruit fibers to produce force because of the denervation/reinnervation remodeling in the VML injured muscles.
Notably, our results also demonstrate the ability of the ENCs to effectively treat a concomitant peripheral nerve injury by bridging the gap between the rerouted nerve and the repair site, as evidenced by direct stimulation of the rerouted nerve, which elicited an action potential to the injury site and generation of muscle force production in most animals in the VML+SMU+ENC group. We were able to directly stimulate the rerouted nerve in a greater percentage of animals in the 6-month repair groups (73% VML+SMU and 93% in the VML+SMU+ENC) versus the 3-month repair groups (25% VML+SMU and 75% in the VML+SMU+ENC). This direct stimulation produced a contraction through the center of the muscle at the site of the VML injury. This suggests that the rerouted nerve was able to successfully form neuromuscular junctions in the injury site, and notably, the ENC was able to effectively bridge the gap between the rerouted nerve and the surgical site in most of the animals. These data suggest that the VML+SMU+ENC animals experienced greater reinnervation through the rerouted nerve.
The fact that it took ∼10–14 days for the nerve to travel through the ENC and reach the VML site, may have given the VML site time to clear the acute inflammatory response. Thus, the ENC routed nerve would not have been exposed and protected from the acute inflammatory environment of the VML injury compared with the nerves' direct reinnervation in the VML+SMU group. This may have led to the increased reinnervation and stimulation potential in the ENC routed repairs. The ability to stimulate the muscle through the ENC in the VML+SMU+ENC group indicates the ENC can serve as a conduit for redirected nerve to promote innervation to a severely damaged VML injury site.
In conclusion, while we demonstrated promise for the SMU graft for partially repairing a VML injury, we also observed issues commonly faced by tissue engineers, significant fibrosis that may play a role decreasing the true potential of the SMU for muscle regeneration. Thus, understanding the acute as well as chronic response of the host to the implanted engineered tissues will be critical for the advancement of this technology. Controlling the fibrotic response during muscle regeneration and remodeling of our SMU will be critical to the translation of our SMU technology to the clinic. Future experiments by our laboratory and others in the VML injury space are actively investigating the acute use of antifibrotic drugs to decrease the fibrotic response for a short period of time to allow implanted tissue to engraft and regenerate muscle without the interference with increased fibrosis in the repair site.37–39
We are also actively investigating the use of protective hydrogels in combination with the SMU technology to acutely protect the implanted tissue from fibrosis. Without the use of a significant intervention like antifibrotic drugs or protective hydrogels, the SMU technology in its current form will have limited success in fully repairing a VML injury site.
Footnotes
Authors' Contributions
The authors confirm contribution to the article as follows: study conception and design: B.L.R. and L.M.L.; data collection: B.L.R., S.S.N., E.E.V.-S., G.P.N., P.C.D.M., and L.M.L.; analysis and interpretation of results: B.L.R., S.S.N., E.E.V.-S., G.P.N., P.C.D.M., and L.M.L.; draft article preparation: B.L.R., S.S.N., and L.M.L. All authors reviewed the results and approved the final version of the article.
Disclosure Statement
The authors have nothing to disclose.
Funding Information
The authors would like to acknowledge their funding source (W81XWH-16-1-0752). Furthermore, the authors would like to acknowledge Charles Roehm (Engineering Design and Fabrication Core, University of Michigan Orthopedic Research Laboratory) and the NIH/NIAMS P30 AR069620 support provided by the Michigan Integrative Musculoskeletal Health Core Center, as well as the Michigan Medicine Department of Pathology.