
Abstract
The innate immune response, particularly the phenotype of responding macrophages, has significant clinical implications in the remodeling outcome following implantation of biomaterials and engineered tissues. In general, facilitation of an anti-inflammatory (M2-like) phenotype is associated with tissue repair and favorable outcomes, whereas pro-inflammatory (M1-like) activation can contribute to chronic inflammation and a classic foreign body response. Biologic scaffolds composed of extracellular matrix (ECM) and, more recently, matrix-bound nanovesicles (MBV) embedded within the ECM are known to direct macrophages toward an anti-inflammatory phenotype and stimulate a constructive remodeling outcome. The mechanisms of MBV-mediated macrophage activation are not fully understood, but interleukin-33 (IL-33) within the MBV appears critical for M2-like activation. Previous work has shown that IL-33 is encapsulated within the lumen of MBV and stimulates phenotypical changes in macrophages independent of its canonical surface receptor stimulation-2 (ST2). In the present study, we used next-generation RNA sequencing to determine the gene signature of macrophages following exposure to MBV with and without intraluminal IL-33. MBV-associated IL-33 instructed an anti-inflammatory phenotype in both wild-type and st2−/− macrophages by upregulating M2-like and downregulating M1-like genes. The repertoire of genes regulated by ST2-independent IL-33 signaling were broadly related to the inflammatory response and crosstalk between cells of both the innate and adaptive immune systems. These results signify the importance of the MBV intraluminal protein IL-33 in stimulating a pro-remodeling M2-like phenotype in macrophages and provides guidance for the designing of next-generation biomaterials and tissue engineering strategies.
Impact statement
The phenotype of responding macrophages is predictive of the downstream remodeling response to an implanted biomaterial. The clinical impact of macrophage phenotype has motivated studies to investigate the factors that regulate macrophage activation. Matrix-bound nanovesicles (MBV) embedded within the extracellular matrix direct macrophages toward an anti-inflammatory (M2)-like phenotype that is indicative of a favorable remodeling response. Although the mechanisms of MBV-mediated macrophage activation are not fully understood, the intraluminal protein interleukin-33 (IL-33) is clearly a contributing signaling molecule. The present study identifies those genes regulated by MBV-associated IL-33 that promote a pro-remodeling M2-like macrophage activation state and can guide future therapies in regenerative medicine.
Introduction
The innate immune system, especially macrophage activation, is recognized to play a determinant role in the structural and functional outcome of engineered tissues following implantation.1,2 Macrophages are highly plastic cells that adopt different phenotypes in response to microenvironmental stimuli. In a simplified paradigm, macrophage phenotype assumes either a pro-inflammatory (M1) state or an anti-inflammatory (M2) state. However, in reality, macrophage phenotype spans a spectrum between these two extremes determined by the many biophysical, biochemical, and molecular factors that constitute the local microenvironment. 3
In the context of tissue repair, the pro-inflammatory M1 macrophage phenotype is typically associated with either scar tissue formation or a nonhealing chronic inflammatory process and a poor downstream outcome.4,5 In contrast, the anti-inflammatory M2 phenotype is generally associated with tissue repair and a more favorable functional outcome.4,5 Biologic scaffolds composed of extracellular matrix (ECM) have been used in several million patients to facilitate a constructive remodeling response for multiple clinical applications, including rotator cuff repair, 6 volumetric muscle loss, 7 ventral hernia repair,8,9 diabetic ulcers, 10 and breast reconstruction, 11 among others.12,13 ECM bioscaffolds direct functional remodeling of site-appropriate tissue in part by inducing an appropriately timed transition from a pro-inflammatory M1-like to an anti-inflammatory M2-like macrophage response.4,5,14
The ECM consists of over 300 structural and functional proteins 15 that in concert direct the overall host immune response to the material. Although intact ECM components, including collagens, laminin, and hyaluronic acid, can interact directly with immune cell receptors to alter cell activity and phenotype,16,17 it is known that degradation of the bioscaffold and subsequent release of signaling molecules is required for anti-inflammatory immunomodulation. 18 Following implantation, matrix remodeling by endogenous enzymes produces a complex milieu of ECM-bound cytokines, growth factors, and protein fragments that mediate immune cell phenotype.15,17,19 Due to the complexity of the ECM and its byproducts, the mechanism of ECM-mediated immunomodulatory activity is only partially understood. The recently described matrix-bound nanovesicles (MBV) have provided further insight into the immunomodulatory properties of ECM. 20
MBV are nanometer-sized lipid-bound vesicles embedded within the fibrous structure of the ECM and released upon matrix degradation. Notably, these vesicles, along with their associated protein, lipid, and miRNA cargo, have been shown in vitro to recapitulate the immunomodulatory effects of the entire ECM. 21 More specifically, isolated MBV alone have been shown to direct macrophages toward an M2-like anti-inflammatory phenotype. Although the molecular mechanism of MBV-mediated macrophage activation is not fully understood, recent work has shown that interleukin-33 (IL-33) contained within the lumen of MBV is a critical contributor.22,23 Specifically, this intraluminal IL-33 is required for MBV-induced activation of macrophages toward an M2-like phenotype and this activation occurs through a noncanonical, receptor-independent mechanism. 22
IL-33 is a member of the IL-1 family that has an N-terminus nuclear localization domain with a chromatin-binding motif.24,25 IL-33 is constitutively expressed in endothelial, epithelial, neuronal, and stromal cells,26–30 and its expression can be augmented by inflammation.27,31,32 IL-33 lacks a signaling sequence and, therefore, has generally been regarded as an alarmin that is released following cell damage. Upon release, IL-33 activates the canonical stimulation-2 (ST2) receptor-signaling pathway in a variety of ST2+ immune cells to stimulate Type I, Type II, and regulatory immune responses.28,33 Full-length IL-33 has also been shown to intrinsically regulate gene expression mediated by its N-terminal domain,27,34–37 however, the nuclear function of IL-33 is specific to cell type.38,39 It has not been determined if nuclear IL-33 affects immune cell gene expression.
The objective of the present study is to establish if IL-33 delivered through MBV can modulate the gene signature of macrophages following exposure to MBV, and specifically to identify genes regulated by IL-33 that are independent of the ST2 receptor. Using transcriptomic analysis of wild-type (wt) and st2−/− macrophages exposed to MBV with (WT) or without IL-33 (IL-33−), we show that ST2 independent signaling by MBV-associated IL-33 regulates expression of anti-inflammatory genes.
Methods
Animals
The wt C57BL/6 mice were purchased from Jackson Laboratories. St2−/− mice obtained from Dr. Anne Sperling (University of Chicago) were originally generated on a BALB/c background 40 and backcrossed 10 times on a C57BL/6 background before use. C57BL/6 il-33−/− mice were a gift from S. Nakae (University of Tokyo, Tokyo, Japan). 41 Animal studies were performed in accordance with all regulations set by the National Institutes of Health and the Animal Welfare Act for use of animals in research (IACUC protocol no. 18083483).
Isolation of MBV
The small intestine was collected from wt or il-33−/− mice and washed thoroughly in phosphate-buffered saline (PBS) to remove all luminal contents. The tissue was comminuted into 1 cm pieces and enzymatically digested with 100 ng/mL Liberase TH (Roche) in buffer (50 mM Tris pH 7.5, 5 mM CaCl2, 150 mM NaCl) for 12 h at room temperature. The digested ECM was then subjected to centrifugation at 500g for 10 min, 2500g for 20 min, and 10,000g for 30 min to remove cell and ECM debris, and the resultant supernatant passed through a 0.22 μm filter. The clarified supernatant was then centrifuged at 100,000g (Beckman Coulter Optima L-90K Ultracentrifuge) at 4°C for 70 min to pellet the now free nanovesicles. The MBV pellet was resuspended in 1 × PBS.
The size and concentration of freshly isolated MBV were measured using Nanosight nanoparticle tracking analysis (Malvern Panalytical). The Brownian motion of the vesicles was used to determine the size distribution as measured across three replicates of 45 s videos for each sample. For all studies herein, vesicle dosage was based on particle count as determined by Nanosight. Following Nanosight analysis, MBV were aliquoted and stored at −20°C until further use.
Isolation and stimulation of bone marrow-derived macrophages
Bone marrow-derived macrophages (BMDMs) were isolated from the tibia and femur of B6 wt and st2−/− mice as previously described. 42 Animals were euthanized by CO2 inhalation and subsequent cervical dislocation in accordance with the guidelines of the American Veterinary Medical Association Panel of Euthanasia. Following euthanasia, the skin of the inferior legs was completely removed and the tibia and femoral bones were isolated. Under sterile conditions, the ends of each bone were then transected, and the bone marrow flushed with medium (Dulbecco's modified Eagle's medium [DMEM]) using a 30G needle. Harvested mononuclear cells were seeded at a ratio of 2 × 106 cells/mL and monocytes were differentiated into macrophages by culture at 37°C and 5% CO2 for 7 days with macrophage colony-stimulating factor-containing complete media (DMEM +10% fetal bovine serum [FBS], 10% L929 supernatant, 0.1% beta-mercaptoethanol, 100 U/mL penicillin, 100 μg/mL streptomycin, 10 mM nonessential amino acids, and 10 mM HEPES buffer). This 7-day differentiation protocol results in a confluent monolayer of naive macrophages.
Naive macrophages isolated from wt or st2−/− mice were exposed to one of the following treatments in DMEM +10% exosome-depleted FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin for 24 h: (1) 20 ng/mL recombinant IL-33 (rIL-33), (2) 4.5 × 109 particles/mL WT MBV, or (3) 4.5 × 109 particles/mL IL-33− MBV. After 24 h, RNA was collected with TRIzol.
RNA isolation
Total RNA was extracted from stimulated macrophages with 800 μL TRIzol reagent using a cell scraper. The solution was mixed with 200 μL chloroform, vortexed for 15 s, and centrifuged at 12,000g for 10 min. The aqueous phase was transferred to a new tube and the RNA precipitated with 3M sodium acetate (1/10 of the volume) and isopropanol (1 volume), followed by centrifugation at 18,000g for 20 min. RNA was purified by washing the RNA pellet in 75% ethanol with an additional centrifugation at 18,000g for 15 min. The RNA pellet was air dried and resuspended in nuclease-free water. The concentration and purity were measured by NanoDrop and Agilent Bioanalyzer 2100, respectively.
RNA-seq and bioinformatic analysis
Library preparation and next-generation RNA sequencing was completed by the Genomics Research Core at the University of Pittsburgh. Bioinformatic analysis was performed by Genevia Technologies (Tampere, Finland). A pipeline in TrimGalore was used to automate quality control by FastQC and adapter trimming by Cutadapt. Reads with a minimum length of 35 bp after trimming were used for downstream analysis. Trimmed reads were aligned to the mouse reference genome (GRCm38) and gene-level counts were obtained simultaneously. The DESeq2 package in R 43 was used to normalize read counts and perform downstream analysis. The Core Analysis function of Ingenuity Pathway Analysis (IPA) software was used to determine the functional relevance of genes differentially regulated by intraluminal IL-33.
Quantitative polymerase chain reaction validation
To validate 16 of the top genes significantly regulated by MBV intraluminal IL-33, 1000 ng of isolated RNA was converted to cDNA using the SuperScript IV First-Strand Synthesis System (Invitrogen). Using a QuantStudio system machine, the quantitative polymerase chain reaction (qPCR) was performed with the TaqMan Fast Advanced Master Mix and TaqMan Advanced Assays for mouse GPR34, Retnlg, Slco2b1, CD28, INPP5J, Klkb11, IL6, SHISA3, Nrxn2, MARCO, SYT7, IL12A, IL19, CD69, CCL4, and CSF3. Mouse hprt1 was used as the housekeeping gene, and the fold change in gene expression following stimulation with WT MBV normalized to IL-33− MBV for each macrophage source was calculated using the ΔΔCt method. A two-way analysis of variance with Sidak's correction was used to determine statistical significance of the log2(fold change) of WT MBV versus IL-33− MBV treatment in each macrophage genotype.
Results
MBV alter gene expression in macrophages independently of ST2 receptor
Next-generation RNA sequencing was used to determine the transcriptomic signature of macrophages following exposure to MBV. Macrophages with (wt) and without (st2−/−) the ST2 receptor were exposed to MBV with (WT) or without IL-33 (IL-33−) to determine the genes specifically regulated by ST2-independent signaling by MBV-associated IL-33. Treatment with rIL-33 was also included to identify MBV-specific effects. The number of differentially expressed genes (DEGs), defined as ±twofold change and p < 0.05, determined by RNA sequencing analysis for each treatment group are shown in Table 1. Exposure of macrophages to MBV had a prominent effect in directing macrophage activation, resulting in over 2000 DEGs relative to naive untreated macrophages regardless of the macrophage genotype or the IL-33 content of the MBV. Visualization by both principal component analysis (Fig. 1A) and Pearson's correlation analysis (Fig. 1B) showed separation of clusters according to group. The first principal component represented 61% of the variance between samples and showed strong separation based on treatment.
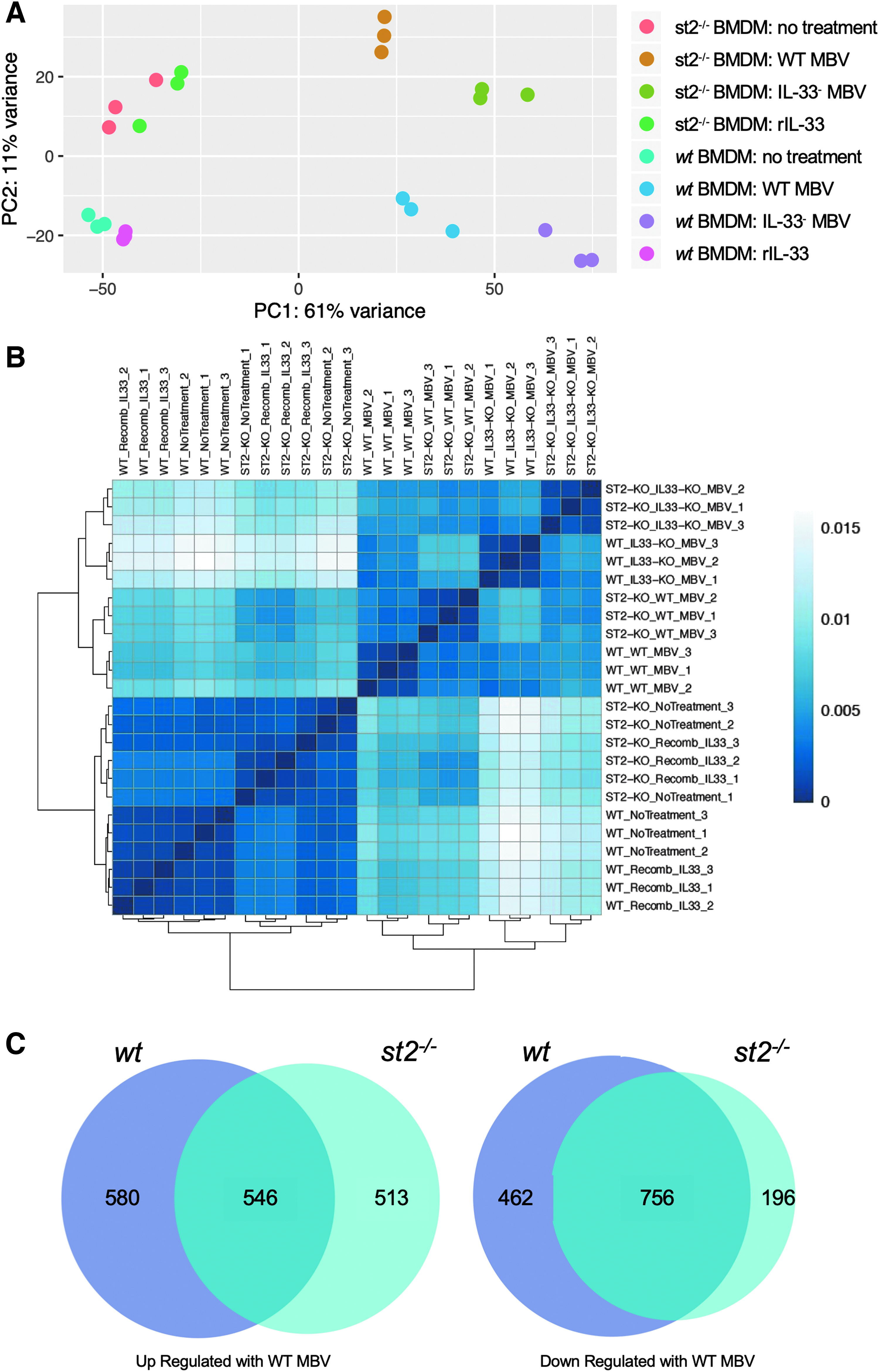
MBV IL-33-induced gene expression in macrophages is independent of ST2 receptor.
Number of Differentially Expressed Genes with ±Twofold Change and p < 0.05 Between Each Treatment Group
BMDM, bone marrow-derived macrophage; DEGs, differentially expressed genes; IL-33, interleukin-33; MBV, matrix-bound nanovesicles; rIL-33, recombinant interleukin-33; wt, wild-type.
The second principal component separated samples according to the genotype of the macrophages (wt vs. st2−/−), but only represented 11% of the variance of the samples indicating minor contribution of the ST2 receptor to the overall macrophage response. Along the first principal axis, there is a significant effect of MBV treatment compared with naive untreated macrophages as well as a distinct contribution of MBV intraluminal IL-33. In st2−/− macrophages the nontreated and rIL-33-treated samples formed two mixed clusters indicating no significant differences between these groups, and the effect of rIL-33 on wt macrophages at the concentration evaluated was limited compared with MBV. A Venn diagram was used to compare the genes up- and downregulated by WT MBV in wt and st2−/− macrophages (Fig. 1C).
Intraluminal IL-33 within the MBV differentially regulates gene expression in macrophages
Exposure of macrophages to WT MBV and IL-33− MBV enabled identification of genes specifically regulated by MBV-associated IL-33. In total, WT MBV versus IL-33− MBV treatment resulted in 1202 and 1514 DEGs in wt and st2−/− macrophages, respectively, suggesting the significance of intraluminal IL-33 in regulating macrophage activation (Table 1). Volcano plots of WT versus IL-33− MBV in wt and st2−/− macrophages show the genes that are specifically up- or downregulated by intraluminal IL-33 within the MBV (Fig. 2A, Supplementary Tables S1–S6). Validation by qPCR on independently generated biologic replicates for 6 of the top upregulated and 10 of the top downregulated DEGs of interest confirmed the directionality of change of results of the sequencing study, although log2(fold change) did not reach statistical significance for all genes by this method.
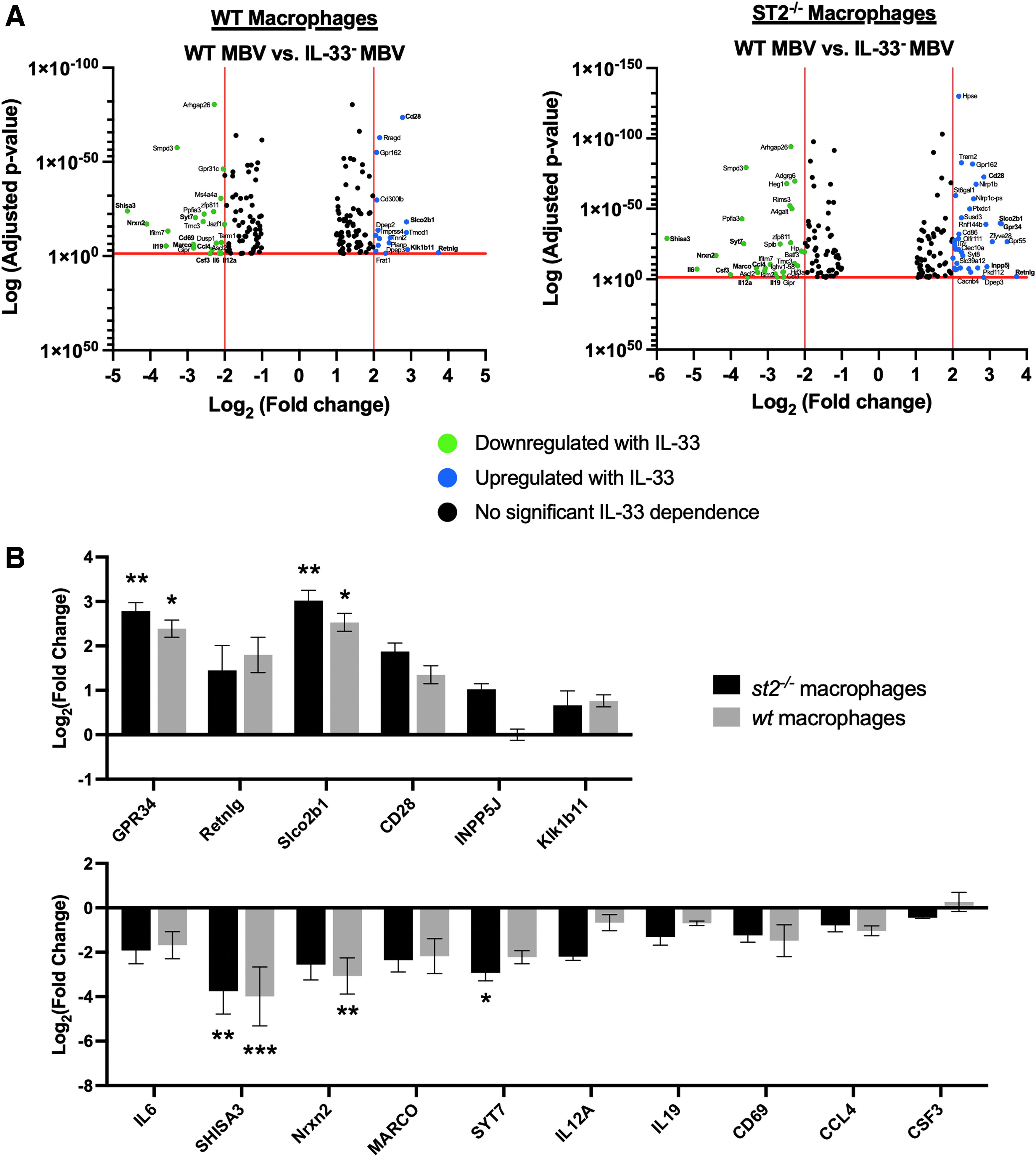
IL-33 content differentially regulates gene expression in macrophages.
The qPCR analysis also demonstrated a similar response to intraluminal IL-33 in macrophages with and without the ST2 receptor (Fig. 2B). Genes upregulated in the presence of intraluminal IL-33 are generally associated with pro-remodeling (M2-like) activity in macrophages while those downregulated are associated with pro-inflammatory (M1-like) effects (Table 2). These results suggest the importance of intraluminal IL-33 in MBV-mediated promotion of an anti-inflammatory M2-like phenotype in macrophages.
Top Regulated Genes Are Associated with Anti-Inflammatory Activation of Macrophages
IFNy, interferon-y; LPS, lipopolysaccharide; TNFα, tumor necrosis factor alpha.
MBV-associated IL-33 regulates pathways of the inflammatory response
The core analysis function in IPA software (Qiagen) was used to identify the top diseases, canonical pathways, and signaling networks that are associated with DEGs induced by intraluminal MBV IL-33 (Fig. 3). The top diseases and pathways showed a strong association of IL-33 with the inflammatory response and cell-to-cell signaling. The full list of the top canonical pathways regulated by MBV-associated IL-33 is shown in Figure 3B. Of note, many of the genes regulated by IL-33+ MBV have an established role in pathways of T cell differentiation, cellular communication of both the innate and adaptive immune systems, and involvement in autoimmune diseases. Analysis of predicted upstream regulators identified activation of the anti-inflammatory IL10RA pathway and downregulation of pro-inflammatory regulators such as nuclear factor kappa B (NF-κB) and tumor necrosis factor alpha by IL-33+ MBV-regulated genes (Fig. 3C). The repeated association of DEGs stimulated by intraluminal IL-33 signaling to the inflammatory response highlights the importance of this intraluminal protein in directing the immune reaction to MBV.
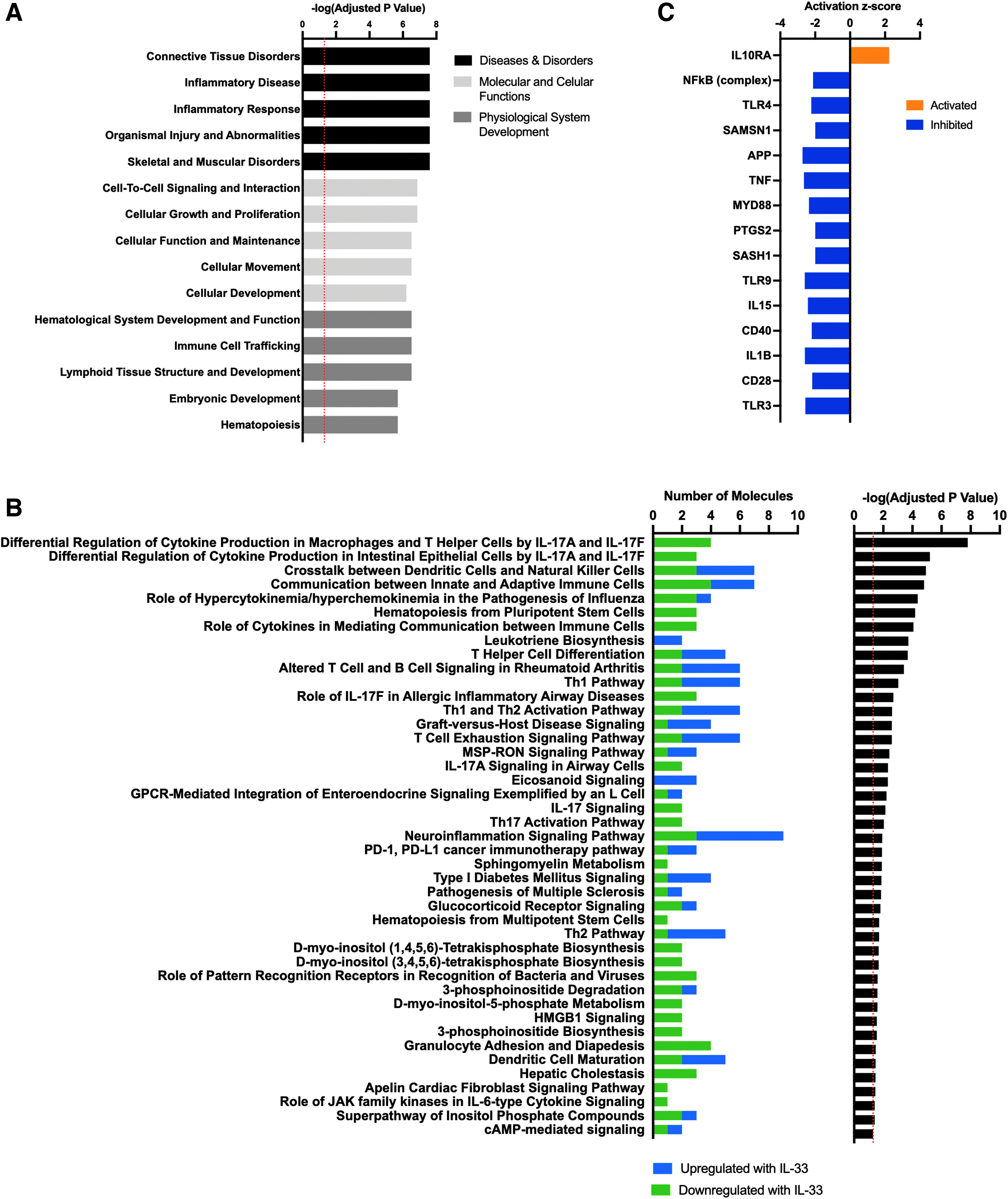
Top pathways regulated by MBV-associated IL-33 are related to the inflammatory response.
Discussion
In the present study, transcriptomic analysis was used to determine the effect of MBV-associated IL-33 on macrophage activation. Results show significant contribution of ST2 receptor-independent IL-33 signaling to the macrophage response to MBV. Specifically, MBV intraluminal IL-33 plays an important role in promoting an anti-inflammatory M2-like macrophage phenotype by upregulating M2-associated and downregulating M1-associated genes. The repertoire of IL-33-mediated genes in macrophages are broadly related to the inflammatory response and crosstalk between cells of the innate and adaptive immune systems.
Implantation of an ECM bioscaffold is typically associated with a constructive remodeling outcome characterized by site-appropriate tissue deposition and absence of foreign body reaction in multiple anatomical locations.7,65–69 An early transition of responding immune cells toward a type 2, anti-inflammatory phenotype is now recognized as a critical determining factor of the positive downstream outcome to ECM scaffolds. 4 Although numerous constituents of the ECM contribute to the overall cellular response to bioscaffold implantation, MBV have emerged as an important, perhaps necessary, factor in directing the anti-inflammatory M2-like activation of macrophages.20,70 In the present study, the potent effect of MBV on macrophage activation is exemplified by the differential expression of over 2000 genes in macrophages following exposure to MBV regardless of the genotype of the cells or IL-33 content of the MBV.
Comprehensive characterization of MBV composition and bioactivity remains an active area of investigation, but lipid, protein, and nucleic acid components likely all contribute to their immunomodulatory effects. Previous work has shown that perturbation of the microRNA content of MBV alters their effects on macrophages. 70 MBV also contain lipid mediators, including resolvin D1 and lipoxin A4, 71 and other cytokines, including ICAM-1, endostatin, and resistin, 22 with established effects on macrophages.72–76
While all MBV components collectively contribute toward an overall anti-inflammatory effect on macrophages, IL-33 in particular is of interest in the present study due to its determinant role in the MBV-mediated macrophage response. Specifically, MBV containing IL-33 produced the expected M2-like activation of BMDM, and elimination of IL-33 within the MBV instead induced a pro-inflammatory phenotype. 22 Owing to the potential clinical importance of IL-33 to MBV effects, the secreted products of macrophages exposed to IL-33+ MBV, but not IL-33− MBV, promoted myogenesis of skeletal muscle progenitor cells in vitro. 22 In a mouse heart transplant model, intraluminal IL-33 was required for MBV to reduce the presence of pro-inflammatory macrophages and mitigate immune-driven chronic rejection of the graft. 23
Consideration of the top signaling pathways predicted by IPA to be regulated by MBV-associated IL-33 may suggest additional or alternative contributions of IL-33 to the biologic and remodeling effects broadly associated with ECM bioscaffolds. Wolf et al. showed that ECM produced from urinary bladder matrix (UBM-ECM), known to contain IL-33+ MBV, 22 synergized with PD-1 and PD-L1 checkpoint blockade to inhibit tumor formation by a macrophage-dependent mechanism. 77 “PD-1/PD-L1 cancer immunotherapy” is among the top pathways regulated by MBV-associated IL-33 in macrophages. Exposure of lipopolysaccharide/interferon-y-activated microglia to either UBM-ECM or UBM-MBV inhibits production of pro-inflammatory cytokines, including IL-33-regulated IL6, consistent with a role of IL-33+ MBV in the “neuroinflammation signaling pathway”. 78
Genes regulated by MBV-associated IL-33 direct pathways of T cell activation and immune cell phenotype, including “communication between innate and adaptive immune cells,” “cytokines mediating communication between immune cells,” “T helper cell differentiation,” and “Th1 and Th2 activation pathway.” Sadtler et al. showed that the pro-regenerative, anti-inflammatory response to ECM scaffolds requires crosstalk between innate and adaptive cells, specifically T helper 2 cells.14,79 Overall, the connection of IL-33 signaling by MBV to known effects of ECM and MBV suggest that intraluminal IL-33 may be a significant, but previously uncharacterized, contributor.
IL-33 is produced as a ∼32 kDa protein that lacks a signaling sequence for secretion but has an N-terminal domain containing a nuclear localization sequence and chromatin-binding motif. Due to the nuclear domain, IL-33 is predominantly found within the nucleus and is constitutively expressed by endothelial, epithelial, and fibroblast cells with little to no basal presence in hematopoietic cells, including macrophages.25,80,81
Following cell death, damage, or mechanical stress, free IL-33 is released into the extracellular space and can stimulate the canonical ST2 signaling pathway in ST2 receptor-positive cells. 28 Of importance, the genes significantly regulated by MBV-associated IL-33 in macrophages are unique from previously established effects of IL-33/ST2 signaling in other cell types. For example, ST2 signaling by IL-33 increased CSF3 expression in dendritic cells 82 and adipocytes, 83 CD69 expression in CD8+ T cells and NK cells,84,85 CCL4 in mast cells,86,87 and IL6 expression in intestinal immune cells, 88 bone marrow mast cells, 86 adipocytes, 83 and dendritic cells 82 : a stark contrast to the downregulation of all these genes by MBV-associated IL-33 signaling in macrophages.
Canonical IL-33/ST2 signaling also decreases CD28 expression in mast cells, 89 whereas noncanonical signaling by MBV increased the expression of this anti-inflammatory marker in macrophages. Activation of MyD88 and NF-kB signaling cascades are also stimulated by IL-33/ST2 signaling in macrophages and other cell types,28,90 but are predicted to be inhibited by MBV-associated IL-33 signaling.
IL-33 has previously been shown to be packaged within membranous vesicles in the cytoplasm of fibroblasts 91 and within exosomes produced by Epstein–Barr virus-infected cells, 92 but the bioactive effects of vesicle-associated IL-33, specifically MBV, has not been previously described. Previous work showed that encapsulation of IL-33 within the MBV lumen enables bypassing of the ST2 surface receptor to stimulate changes in macrophage phenotype. 22 Herein, it is further shown that intraluminal IL-33 delivered by MBV induces transcriptomic changes independent of the ST2 receptor. In other cell types, nuclear IL-33 binds chromatin through protein–protein interactions and acts as a transcriptional repressor.25,34 The present study showed a near doubling of significantly regulated genes following MBV treatment versus no treatment in the absence of IL-33 (4289 vs. 2344 in wt and 2011 vs. 3862 in st2−/− macrophage) potentially supporting IL-33 as a transcriptional repressor in macrophages as well. Full-length IL-33 can bind directly to the p65 and p50 subunits of NF-kB in HEK293 cells to dampen pro-inflammatory signaling 34 (1,31-33).
In endothelial cells, IL-33 has also been shown to suppress transcription of the pro-inflammatory cytokine IL6 by binding to promoter-bound proteins. 93 In the present work, IL-33 within MBV significantly decreased IL6 gene expression, and IPA analysis identified NF-kB as a predicted upstream regulator that is downregulated by the presence of IL-33 within the MBV. The present results suggest a similar mechanism of NF-kB inhibition following IL-33 delivery to macrophages.
The parallels between the role of nuclear IL-33 in other cells and the response of macrophages to MBV informs a hypothesis that following uptake, IL-33 may translocate to the nucleus and interact with chromatin to diminish pro-inflammatory activities in macrophages. In this hypothesis, MBV may act as a vector for nucleus-to-nucleus transport of proteins to direct cell activation in a previously uncharacterized paradigm of signaling. Although results shown in this study apply specifically to IL-33, this model of receptor-independent delivery of intraluminal MBV proteins may also apply to other nuclear alarmins and cytokines lacking a signaling sequence. Future studies will explore this novel hypothesis and confirm that nuclear localization of IL-33 occurs following MBV uptake in macrophages and is at least partially responsible for transcriptomic regulation.
Conclusion
The finding that receptor-independent delivery of MBV-associated IL-33 is sufficient for directing an anti-inflammatory macrophage response highlights the importance of this cytokine in determining the host response and remodeling outcome to MBV and ECM. Furthermore, understanding the importance of IL-33 signaling is relevant for intelligent design of the next generation of ECM-based therapies.
Footnotes
Acknowledgments
The authors thank Anna Lucas for her animal husbandry support, Deborah Hollingshead for conducting the RNA-seq, and Jane Pulman from Genevia for the bioinformatic analysis. This project used the University of Pittsburgh HSCRF Genomics Research Core.
Disclaimer
This work is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the following grants: NIH R01AR073527 to S.F.B. and H.R.T.; R01HL122489 to H.R.T.; and F31HL151083 to M.C.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.