
Abstract
Cardiac tissue engineering has been working to alleviate the immense burden of cardiovascular disease for several decades. To improve cardiac tissue homogeneity and cardiomyocyte (CM) maturation, in this study, we investigated altering initial encapsulation geometry in a three-dimensional (3D) direct cardiac differentiation platform. Traditional engineered cardiac tissue production utilizes predifferentiated CMs to produce 3D cardiac tissue and often involves various cell selection and exogenous stimulation methods to promote CM maturation. Starting tissue formation directly with human induced pluripotent stem cells (hiPSCs), rather than predifferentiated CMs, simplifies the engineered cardiac tissue formation process, making it more applicable for widespread implementation and scale-up. In this study, hiPSCs were encapsulated in poly (ethylene glycol)-fibrinogen in three tissue geometries (disc-shaped microislands, squares, and rectangles) and subjected to established cardiac differentiation protocols. Resulting 3D engineered cardiac tissues (3D-ECTs) from each geometry displayed similar CM populations (∼65%) and gene expression over time. Notably, rectangular tissues displayed less tissue heterogeneity and suggested more advanced features of maturing CMs, including myofibrillar alignment and Z-line formation. In addition, rectangular tissue showed significantly higher anisotropic contractile properties compared to square and microisland tissues (MI 0.28 ± 0.03, SQ 0.35 ± 0.05, RT 0.79 ± 0.04). This study demonstrates a straightforward method for simplifying and improving 3D-ECT production without the use of exogenous mechanical or electrical pacing and has the potential to be utilized in bioprinting and drug testing applications.
Impact statement
Current methods for improving cardiac maturation postdifferentiation remain tedious and complex. In this study, we examined the impact of initial encapsulation geometry on improvement of three-dimensional engineered cardiac tissue (3D-ECT) production and postdifferentiation maturation for three tissue geometries, including disc-shaped microislands, squares, and rectangles. Notably, rectangular 3D-ECTs displayed less tissue heterogeneity and more advanced features of maturing cardiomyocytes, including myofibrillar alignment, Z-line formation, and anisotropic contractile properties, compared to microisland and square tissues. This study demonstrates an initial human induced pluripotent stem cell-encapsulated rectangular tissue geometry can improve cardiac maturation, rather than implementing cell selection or tedious postdifferentiation manipulation, including exogenous mechanical and/or electrical pacing.
Introduction
Heart disease is the leading cause of death worldwide in both men and women of nearly all ethnicities. 1 With limited regenerative capacity, diseased cardiac tissue is not able to fulfill its electrical and mechanical roles, including electrical signal propagation and blood circulation, often leading to arrhythmias and sudden cardiac death. Heart transplants remain the standard of care for treating heart failure, but as a result of the highly limited supply of organs for transplant, this costly and invasive option is not always available. 2 Cardiac tissue engineering provides the potential to create functional replacement tissues for use in cardiac regenerative medicine and as an in vitro platform for testing new therapeutics.
With the discovery of human induced pluripotent stem cells (hiPSCs) in 2007, 3 employing hiPSC-derived cardiomyocytes (hiPSC-CMs) has become customary in the field of cardiac tissue engineering. HiPSCs are differentiated down the cardiac lineage using a series of small molecules and growth factors, often in the form of embryoid bodies4,5 or monolayers.6,7 Extensive research is being conducted using hiPSC-CMs to recapitulate the native tissue functionality, but there are major challenges in producing cells that mature to the adult CM phenotype, in particular, with respect to cell morphology and alignment, electrophysiological function, calcium handling, and gap junction distribution.8,9
One of the key components of cardiac maturation in vivo is the alignment of CMs to form highly aligned myofibers, which are important for contraction 10 and electrophysiological function. 11 In the native myocardium, the CMs orient themselves longitudinally to allow for electric coupling through intercalated discs at the ends of the rod-shaped cells, where most gap junctions reside, resulting in anisotropic action potential propagation. 12 The organization and distribution of gap junctions in neighboring cells determine the current resistance, and therefore electrical function of the tissue. 13 In vivo, anisotropic action potential results in faster propagation in the longitudinal direction than in the transverse direction. 14
This orientation allows for the individual CMs to function together as a single contractile tissue for proper cardiac function. In vitro, engineered cardiac tissues often contain fetal-like CMs with randomly dispersed gap junctions, resulting in isotropic action potential propagation. 8 Improving in vitro CM morphology and alignment, thereby promoting CM maturation, is critical for applicability in therapeutic testing and disease modeling.
Several techniques have been employed to improve cell organization in vitro, including micropatterning and nanopatterning.15,16 However, these experiments often required specialized equipment and are performed in two-dimensional monolayers rather than 3D tissues. In addition, many of these platforms require predifferentiated CMs, which prevent us from studying CM alignment throughout development. The platform employed in this study provides a simplified, less labor-intensive approach to producing 3D engineered cardiac tissue that does not require predifferentiated CMs or specialized equipment for driving maturation. While other systems have to initiate tissue production and differentiation before implementing the maturation strategy, our method incorporates the maturation strategy into the initial tissue production step.
We have previously shown the ability to encapsulate hiPSCs in poly (ethylene glycol) fibrinogen (PEG-fibrinogen) to form 3D human engineered cardiac tissues (3D-ECTs) in a “microisland” disc-shaped geometry. While this study was highly successful, resulting in 3D-ECTs that presented with T-tubules adjacent to Z-lines, a key component of functional maturation, 17 contraction often propagated in a circular direction along the edges of the microisland, which is not typical of a contracting human myocardium.
In this study, we investigated the promotion of cardiac tissue maturation on the macroscale by changing the initial hiPSC encapsulation geometry. To do this, we compared three different tissue geometries: a rectangular tissue with a 5:1 length to width ratio, a square tissue with a 1:1 length to width ratio, and our established microisland geometry. The square geometry was selected to mimic the microislands, while providing a straight outer edge for alignment. The rectangular geometry was designed to simulate a linear representation of the dense tissue ring that we saw in previous studies. In this study, we show that the rectangular geometry resulted in less tissue heterogeneity and improved cardiac tissue maturation, resulting in the tissues more closely mimicking the native myocardium. The knowledge gained in this study will impact future production of engineered cardiac tissue for studying disease mechanisms and cardiac developmental biology.
Materials and Methods
All materials were purchased from Sigma-Aldrich, unless otherwise stated.
3D-ECT formation
HiPSCs (IMR90 clone 1 (WiCell); IRB determined project to be exempt from oversight) were dissociated using Versene for 4.5 min at 37°C. HiPSCs were resuspended in mTeSR medium and centrifuged for 5 min at 200 g. The supernatant was removed leaving a cell pellet, which was resuspended in polymer precursor solution at a concentration of 60 million hiPSCs/mL of precursor solution using a wide orifice pipet. Eight microliters of cell suspension was pipetted into each tissue geometry mold on acrylated glass coverslips. The solution was crosslinked using visible light for 30 s.
The polydimethylsiloxane (PDMS) mold was carefully removed, and the attached tissues were placed in a six-well plate with 3 mL of mTeSR medium supplemented with 10 μM Y-27632 (ROCK inhibitor, RI) (day −3). On the following 2 days (day −2 and day −1), the spent medium was replaced with 3 mL of fresh mTeSR medium. On day 0, cardiac differentiation was initiated using established protocols 6 where 12 μM CHIR99021 was added to RPMI B27 minus insulin (Thermo Fisher Scientific). Exactly 24 h later (day 1), the media were replaced with fresh RPMI B27 minus insulin. Exactly 48 h later (day 3), half of the spent media was combined with an equal amount of fresh RPMI B27 minus insulin supplemented with 5 μM IWP2. Exactly 48 h later (day 5), spent media were replaced with fresh RPMI B27 minus insulin. On day 7 and every 3 days thereafter, 3D-ECTs were fed with RMPI B27.
All methods for glass acrylation, PDMS mold preparation, PEG-fibrinogen synthesis, hiPSC culture, and tissue characterization are listed in the Supplementary Methods.
Results
HiPSCs were encapsulated in three tissue geometries before cardiac differentiation
In this study, we demonstrate that initial encapsulation geometry of hiPSCs impacts the functionality of resulting 3D-ECTs. Building on prior work, molds were designed to ascertain whether a straight edge and shorter edge to middle distance would promote CM alignment and maturation. The microisland geometry served as the control, with an edge to middle distance of 2 mm and tissue thickness of 700–800 μm. The square geometry design incorporated straight edges, while maintaining the 2 mm edge to middle distance. The rectangular geometry design aimed to mimic a straightened version of the outer ring that formed throughout the microisland differentiation, with an edge to middle distance of 0.75–1 mm at any given point along the perimeter of the rectangle.
Custom PDMS molds in microisland, square, and rectangular geometries were produced using 3D printed positive casts (Supplementary Fig. S1), such that all geometries would have the same initial tissue volume and thickness. HiPSCs were encapsulated at 60 million cells/mL in PEG-fibrinogen (Fig. 1) in each geometry similar to previously established methods17,18 through free radical polymerization using the photoinitiator Eosin Y and visible light. On day −2 (24 h postencapsulation), hiPSC colonies appeared evenly distributed throughout in all tissue geometries (Fig. 1).
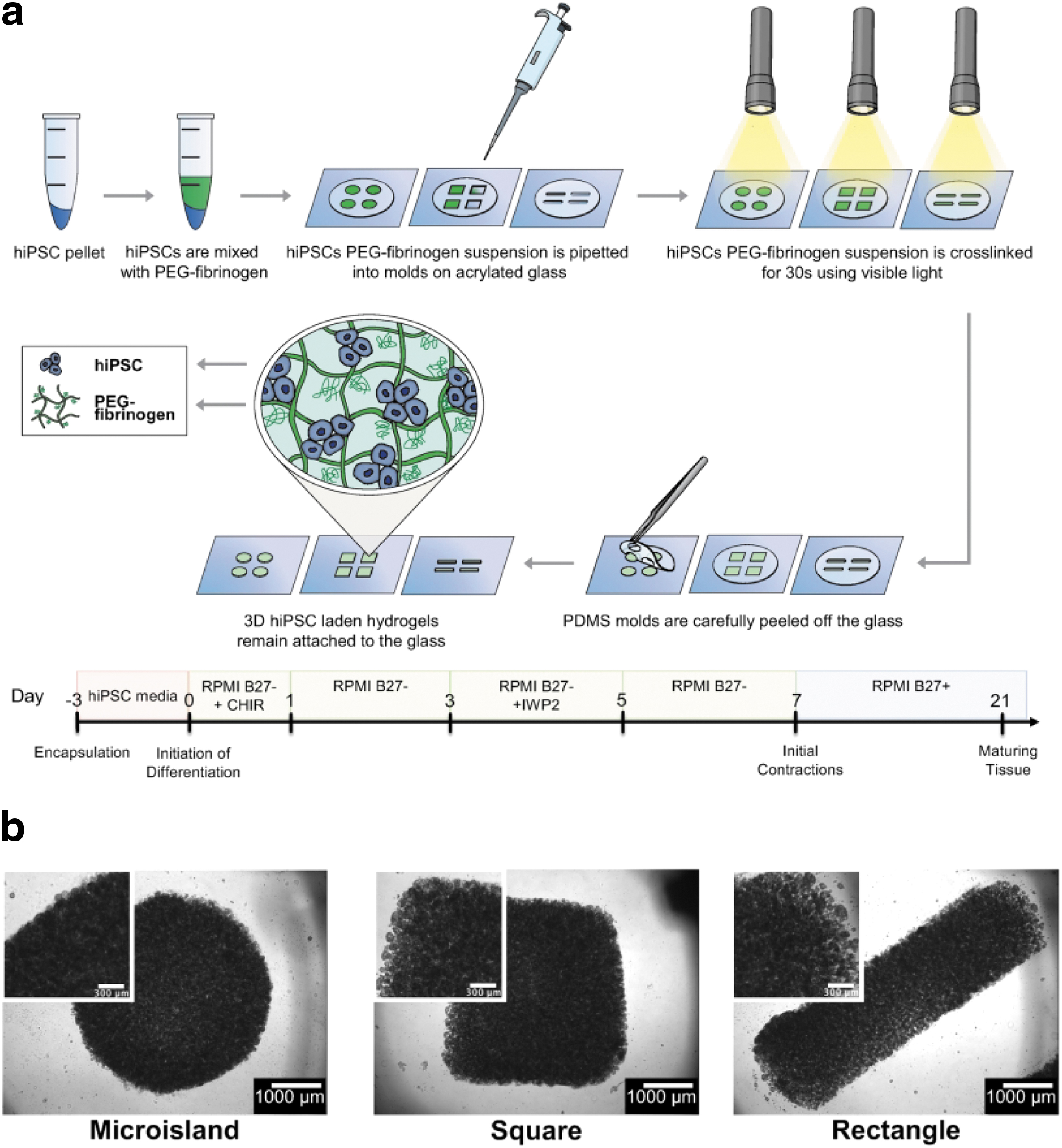
HiPSCs were encapsulated and differentiated in microisland, square, and rectangular geometries.
HiPSCs remained viable throughout encapsulation and differentiation
Cell viability was monitored throughout differentiation in regions on the edge and middle of each tissue. On day −2 (24 h post-encapsulation), viable cell density appeared uniform both within and between each tissue geometry (Fig. 2b and Supplementary Fig. S2 and S3). At the onset of differentiation on day 0, viable cell colonies both on the edge and in the middle increased in size for all tissue geometries as hiPSCs proliferated in the 3D PEG-fibrinogen microenvironment; PEG-fibrinogen can be degraded and remodeled by encapsulated cells. 19
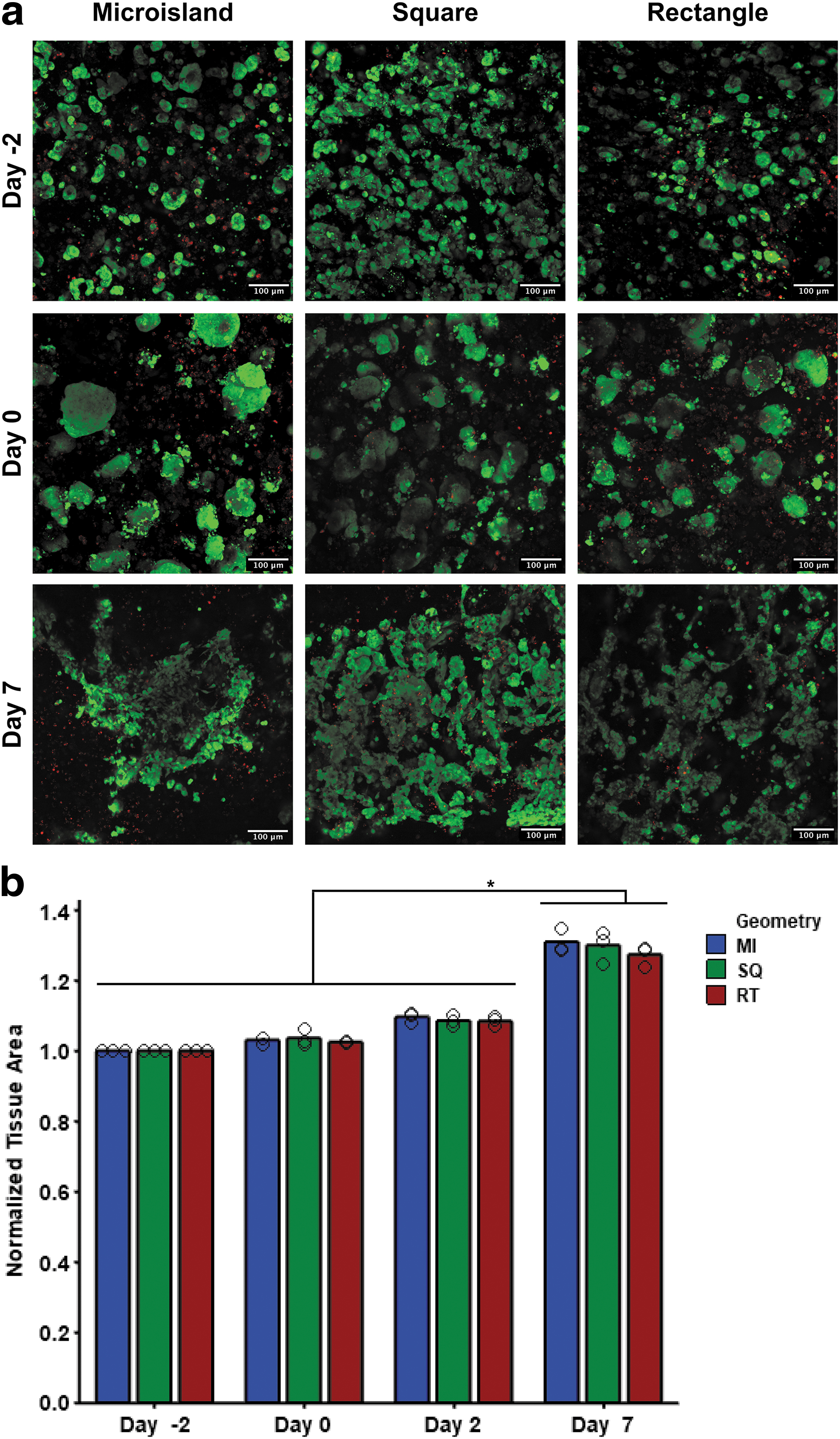
HiPSC colonies remained viable and grew in the 3D PEG-fibrinogen microenvironment postencapsulation.
By day 7 of differentiation, viable cell density was similarly uniformly distributed around the edges of each tissue geometry; however, this uniform distribution of viable cells was not maintained in the middle regions of microisland and square tissues (Supplementary Fig. S3). Rectangular tissues showed a more homogenous regional cell viability throughout differentiation. Tissue area for each geometry increased comparably (MI 31.0 ± 3.3%, SQ 28.4 ± 4.5%, RT 27.5 ± 2.9%) throughout differentiation from day −2 to 7 (Fig. 2b).
Engineered cardiac tissue geometry does not impact cardiac differentiation
On day 10 of differentiation, cell population was analyzed by dissociating 3D-ECTs and performing flow cytometry using a fixable viability dye and cardiac markers, cardiac troponin T (cTnT) and MF20. All tissue geometries showed similar percentages of cTnT+ cells (cTnT: MI 59.5 ± 10.1%, SQ 61.4 ± 5.8%, RT 66.7 ± 4.6%; MF20: MI 57.9 ± 10.2%, SQ 61.7 ± 6.1%, RT 67.0 ± 5.3%; Fig. 3a–d, f), as expected for this differentiation protocol without the use of CM selection.17,18
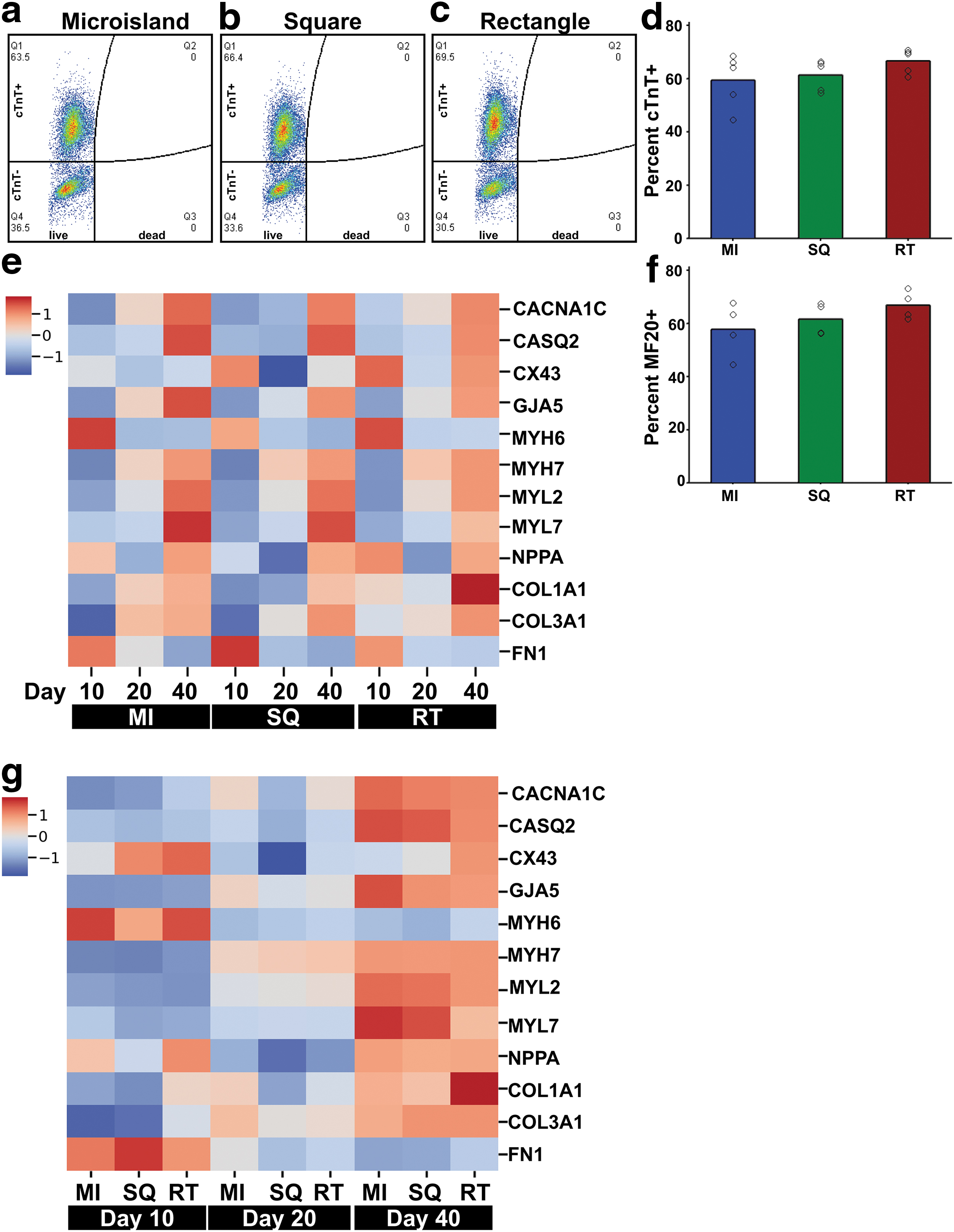
HiPSCs differentiated in all tissue geometries and showed temporally appropriate changes in gene expression.
Cardiac maturation was also assessed by reverse transcription quantitative polymerase chain reaction using a variety of genes that influence electrophysiology, calcium handling, contractility, and extracellular matrix (ECM) production. 3D-ECTs from each geometry showed temporally appropriate changes in gene expression (Fig. 3e, g, Supplementary Fig. S4). No significant difference was observed between tissue geometries at each time point. In the microisland and square tissues, the majority of the cells are on the edges. Since these areas of high cell density in the microisland and square tissues develop and mature similarly to the cells throughout the rectangular tissues, the similar gene expression profiles were somewhat expected.
Cardiac gene MYH7 (β-myosin heavy chain [β-MHC]) expression increased over time with a subsequent temporal decrease of MYH6 (α-myosin heavy chain [α-MHC]) expression, revealing a transition from fetal isoform α-MHC to the adult isoform β-MHC (Supplementary Fig. S4e, f). In addition, MYL2 (ventricular myosin light chain 2) expression increased over culture time, as expected based on previous studies (Supplementary Fig. S4g). 17 All geometries showed increased expression of GJA5 (connexin 40) and COL3A1 (Collagen Type III Alpha 1 Chain) over time (Supplementary Fig. S4d, k). Microisland and square geometries showed an upregulation of MYL7 (atrial myosin light chain 2) expression from day 20 to 40, which was not apparent in rectangular tissues (Supplementary Fig. S4h).
Rectangular tissues show less spatial heterogeneity compared to microisland and square tissues
By day 40 postdifferentiation, an outer tissue ring formed on both microisland and square tissue edges, leaving the middle of tissues less dense, while rectangles showed minimal areas of low cell density (Fig. 4a). To visualize the features of CMs inside the ECTs, whole tissue immunostaining was performed using alpha sarcomeric actinin (αSA), connexin 43 (Cx43), and Hoechst (Fig. 4b). Tissues from each geometry displayed similar bulk tissue characteristics on the tissue edge.
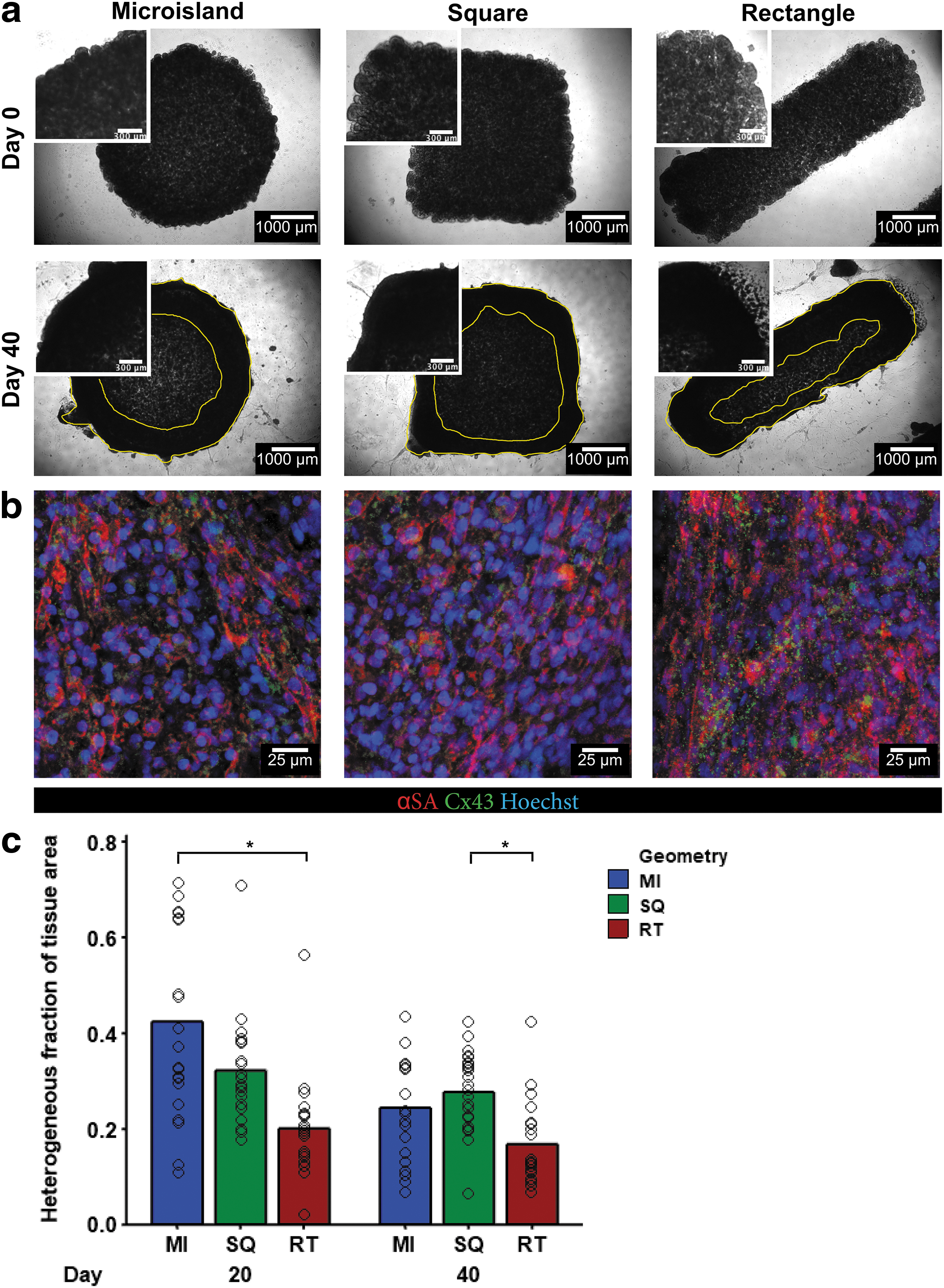
HiPSCs in 3D-ECTs underwent successful cardiac differentiation and remodeled their microenvironment over time.
The distribution of cells throughout tissue culture showed spatial heterogeneity between the center and edge of the tissue. To quantify this heterogenous fraction in each tissue geometry, total tissue area was manually measured from phase contrast images on day 20 and 40. Next, the heterogenous region inside the tissue was measured and divided by the total tissue area resulting in a fraction from 0 to 1. Rectangular tissues showed less regional heterogeneity on day 20 compared to their time-matched microisland tissue and on day 40 compared to square tissues (day 20: MI 0.4 ± 0.2, SQ 0.32 ± 0.11, RT 0.20 ± 0.10; day 40 MI 0.24 ± 0.11, SQ 0.27 ± 0.08, RT 0.17 ± 0.09; Fig. 4c).
All tissue geometries contract at similar frequencies and velocities at late time points
Spontaneous contraction was recorded on days 20 and 40 and was analyzed using an open-source MATLAB code. 20 Whole tissue averages were calculated by taking the average of several regions per tissue in each geometry. Frequency of contraction increased for all geometries from day 20 to 40 (Fig. 5a). The time interval between contraction and relaxation was significantly longer in square and rectangular tissues on both day 20 and 40 (Fig. 5b). On day 20, contraction and relaxation velocity were significantly slower in rectangles compared to microislands (Fig. 5c, d), but these differences were not apparent by day 40. On day 40, the frequency, contraction velocity, and relaxation velocity increased for all geometries, as expected with prolonged culture time.
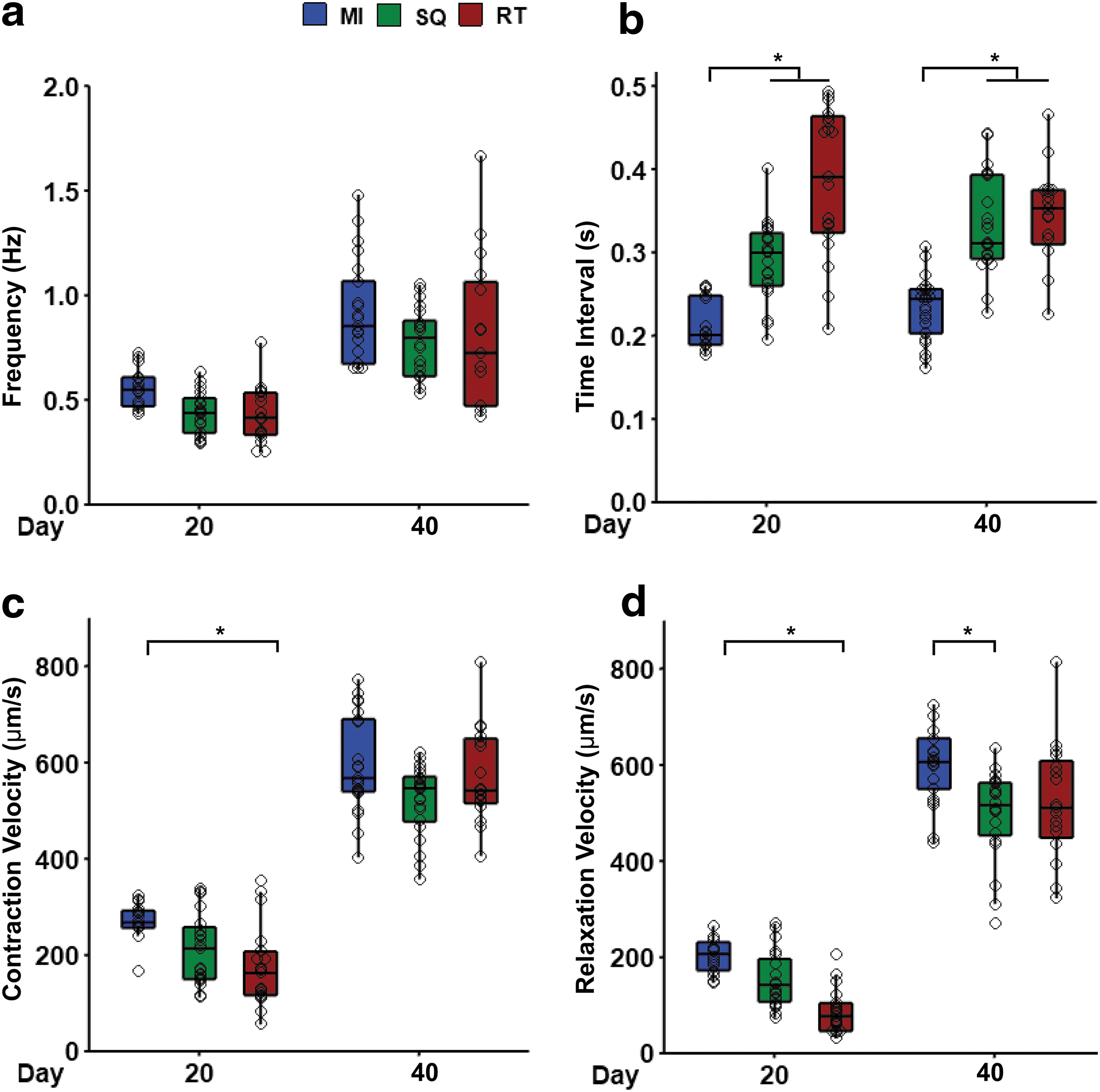
ECTs exhibited an increase in contraction velocity with tissue development in all geometries.
Rectangular tissues show unidirectional contraction
While tissue geometries contracted at similar frequencies, the qualitative videos showed differences in the way the different geometries contracted (Supplementary Movies S1–S3). Microisland and square tissues contracted from the dense outer band inward toward the center of the tissue in all directions. The rectangular tissues contracted along their longitudinal axis. To quantify this, velocity vectors of spontaneous contraction were analyzed on day 40. Frames of a maximum contraction and maximum relaxation were superimposed with their vector fields (Fig. 6a, red lines). These images were then used to create histograms for the frequency of each angle.
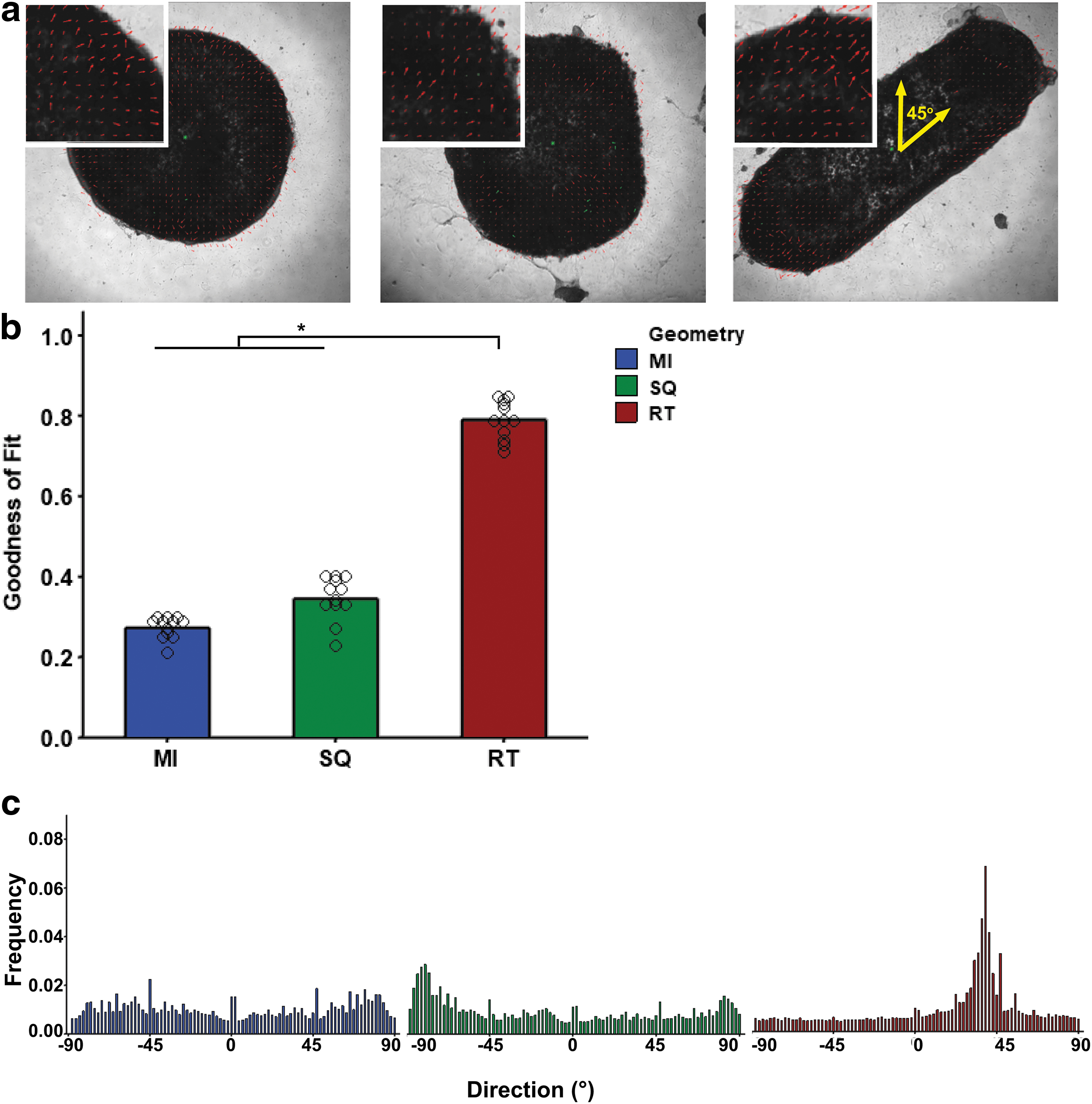
Rectangular ECTs exhibited anisotropic contraction along a longitudinal axis, whereas the other geometries did not.
Histograms for microisland and square geometries showed no major peaks corresponding to a single directionality (Fig. 6c). Rectangular geometries demonstrated a single peak at 45°, which corresponded with the longitudinal length of the tissue. Goodness-of-fit values for each histogram confirmed that the rectangular geometry had significantly more uniform contraction directionality compared to both microisland and square geometries (MI 0.28 ± 0.03, SQ 0.35 ± 0.05, RT 0.79 ± 0.04), where 0 represents no fit and 1 represents a perfect fit (Fig. 6b). Similar to the native myocardium, the rectangular tissue showed anisotropic contraction along the longitudinal axis.
Square and rectangular 3D-ECTs show more mature structural features
3D-ECTs were dissociated and plated on fibronectin-coated coverslips for single-cell analysis. To visualize sarcomeres and gap junction proteins, dissociated cells were stained with αSA, Cx43, and Hoechst (Fig. 7a). Sarcomeres were ∼1.8 μm in length, agreeing with literature values for hiPSC-CMs, 8 and showed no significant difference between tissue geometries. To further visualize ultrastructural features, transmission electron microscopy (TEM) imaging was performed on late time point (day 140) samples (Fig. 7b). These images revealed potential signs of higher maturation in square and rectangular tissues, including sarcomeres with regularly spaced Z bands and adjacent mitochondria compared to microisland tissue, which presented with an immature arrangement of myofibrils and mitochondria with rudimentary sarcomere structures containing Z bodies.
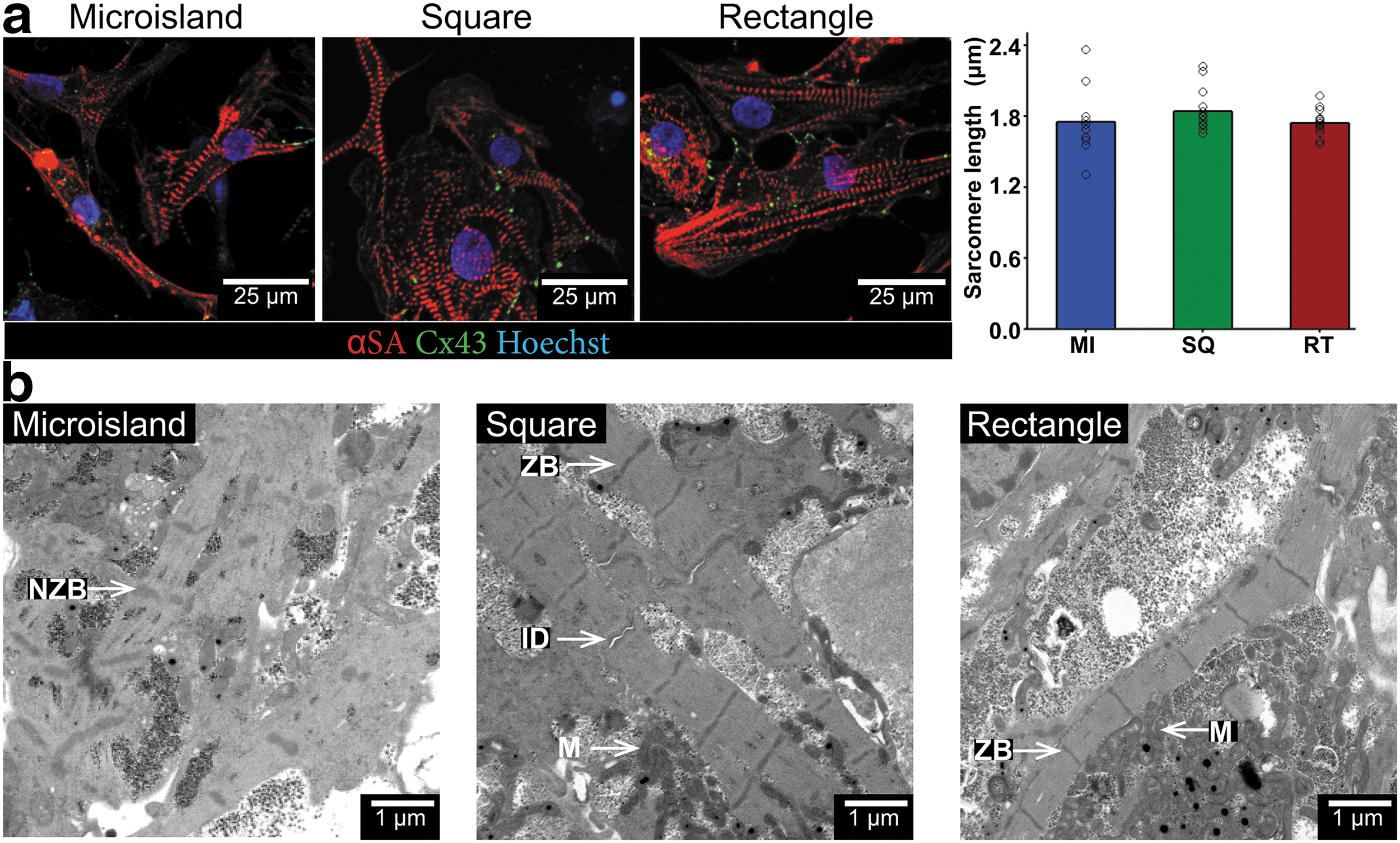
Rectangle and square 3D-ECTs show features of advancing maturity compared to microislands. 3D-ECTs from each geometry were dissociated, plated on coverslips, and immunostained for sarcomere quantification (αSA-red, Cx43-green, and Hoechst-blue).
Rectangular 3D-ECTs show increasing trend of conduction velocity with increased external pacing
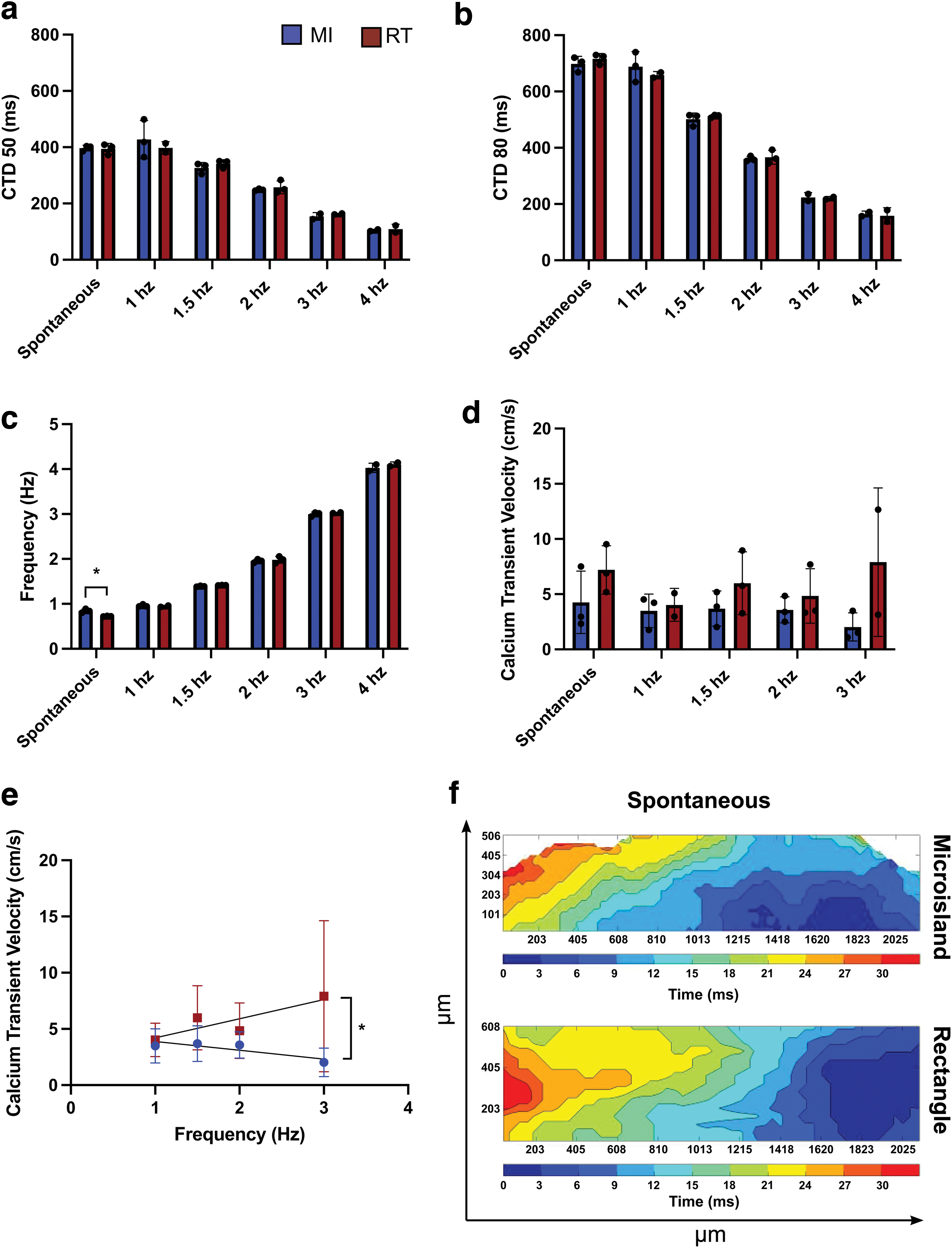
To assess electrophysiological responses, microisland and rectangle 3D-ECTs were optically mapped. 3D-ECT spontaneous depolarization rates were 0.84 ± 0.04 Hz for MI and 0.72 ± 0.01 Hz for RT. 3D-ECTs for both geometries exhibited 1:1 capture during external pacing up to 4 Hz. During spontaneous depolarization, calcium transient durations at 50% (CTD50) and 80% (CTD80) repolarization for MI were 397 ± 10 and 698 ± 27 ms, and for RT, they were 394 ± 19 and 716 ± 19 ms, respectively (Fig. 8a–c, n = 3 tissues). Calcium transient velocities were 4.2 ± 2.8 cm/s for MI and 7.2 ± 2.2 cm/s for RT (Fig. 8d, n = 3 tissues).
Optical mapping of microisland and rectangle 3D-ECTs.
When 3D-ECTs were electrically paced at 1.0, 1.5, 2.0, 3.0, and 4.0 Hz, the CTD50 were 427 ± 67, 326 ± 19, 248 ± 3, 154 ± 13, and 104 ± 4 ms, respectively, for MI and 397 ± 22, 341 ± 14, 257 ± 23, 162 ± 2, and 109 ± 17 ms for RT. The CTD80 were 698 ± 27, 688 ± 53, 501 ± 22, 360 ± 9, 223 ± 17, and 164 ± 10 ms for MI and 658 ± 13, 513 ± 5, 366 ± 24, 221 ± 5, and 158 ± 29 ms for RT (Fig. 8a, b, n = 3 tissues). Calcium transient velocities for MI 3D-ECTs were 3.5 ± 1.5, 3.7 ± 1.6, 3.6 ± 1.2, and 2.0 ± 1.3 cm/s for 1.0, 1.5, 2.0, and 3.0 Hz pacing, respectively; RT calcium transient velocities were 4.0 ± 1.5, 6.0 ± 2.8, 4.8 ± 2.5, and 7.9 ± 6.7 cm/s (Fig. 8d, n = 3 tissues). In both cases, calcium transient velocities for 4 Hz recordings were unable to be accurately analyzed.
Discussion
In this work, we produced 3D-ECTs in three tissue geometries and investigated the impact of initial tissue geometry on cardiac differentiation, maturation, and functionality. The hybrid biomaterial, PEG-fibrinogen, was selected to provide the 3D microenvironment based on its relevance in clinical trials 21 and previously published work. Previous microisland studies using both PEG-fibrinogen 17 and GelMA 18 produced functional cardiac tissues that presented with adult-like ultrastructural features, including T-tubules adjacent to Z lines and appropriate responses to external stimuli. While these studies provided vital foundational information, the microisland tissue geometry (diameter ∼4 mm) resulted in a dense outer ring of contracting tissue with a less viable tissue center.
To alleviate regional tissue heterogeneity and improve cardiac tissue maturation, in this study, two new encapsulation geometries were investigated. The square geometry provided a straight edge for cell alignment, while maintaining similar edge to center (roughly 4 × 4 mm) distances as the microisland. The rectangular geometry was designed to mimic a straightened version of the dense outer ring formed in the microisland tissues (1.5 × 5 mm). All three tissue geometry designs maintained the same initial tissue volume, cell density, and tissue thickness. HiPSCs were encapsulated in PEG-fibrinogen in each of the three initial tissue geometries on acrylated glass coverslips and differentiated, forming spontaneously contracting 3D-ECTs by day 7.
The similarities in differentiation efficiency, gene expression, and contraction velocity between geometries were well supported by the observation that each of these characterization methods is primarily assessing the properties of the dense outer band of cardiac tissue, which contains many more cells than the middle of the microisland and square 3D-ECTs. Early time point characterization, including day 10 cardiac differentiation efficiency and gene expression, showed that initial encapsulation geometry does not impact whether successful cardiac differentiation occurs. Differentiation efficiency in all three tissue geometries was similar to values reported in the literature for direct cardiac differentiation of hiPSCs in 3D.17,18,22 While there was no difference in cardiac differentiation efficiency between geometries, current literature supports in situ formation of engineered cardiac tissues through 3D hiPSC cardiac differentiation over the use of predifferentiated CMs for engineered tissue formation.
Direct cardiac differentiation of hiPSCs to cardiac tissues results in high cell densities as cells continue to proliferate postencapsulation and enables elimination of the CM dissociation step postdifferentiation, which disrupts important cellular connections.17,22
This study demonstrated differences in CM functionality in response to changes in initial hiPSC encapsulation geometry. Comparable percentages of CMs postdifferentiation provided a baseline for evaluation of the impact of initial encapsulation geometry on cardiac tissue function and maturation with extended culture time. Further study is warranted to provide additional insights, including in-depth analysis to assess potential differences in the non-CM, cTnT-negative cell populations. Since differences in the efficiency of differentiation were not identified, evaluation here did not focus on the impact of tissue geometry on shorter-term key milestones of differentiation. Rather, the important differences in cardiac tissue maturation and functionality were observed at later time points between tissues geometries when tissues were cultured long term (30–150 days), as evidenced by reduced tissue heterogeneity and enhanced myofibrillar organization.
Enhanced maturation is critical for applications in pharmaceutical development and regenerative medicine. Unfortunately, most engineered cardiac tissue never progresses beyond the fetal developmental milestones, limiting its applicability in preclinical studies and cell therapy. Several methods of varying complexity have been utilized to enhance maturation, including, but not limited to, altering substrate stiffness, contact guidance through micropatterning, and electrical and mechanical stimulation. 23
Plating CMs on stiffer substrates (up to 10 kPa) has been shown to increase CM force generation and improve calcium handling. 24 Similarly, seeding CMs on nanopatterned 15 micropatterned 16 surfaces has been shown to improve CM function and myofibrillar alignment by mimicking nanoscale cues seen in native ECM and the microscale cues seen in myocardial architecture. While these methods are applicable to predifferentiated CM monolayers, they are not relevant for 3D engineered cardiac tissue maturation, which is composed of several layers of CMs. Introduction of intermittent electrical 25 and/or mechanical26,27 stimulation postdifferentiation has been shown to improve cardiac electrophysiology and increase myofibril alignment over time in 3D engineered tissues. However, these methods sacrifice simplicity for enhanced maturation, which may not enable for widespread acceptance in cardiac tissue engineering and pharmaceutical testing applications.
In this study, by modulating the initial geometry of hiPSC-laden, engineered tissues before cardiac differentiation, developing CMs in the rectangular tissues experience both contact guidance as a result of the macroscale tissue axis and then experience aligned mechanical stimulation as a result of endogenous tissue contraction starting on approximately differentiation day 10. The developing engineered cardiac tissues are crosslinked to the acrylated glass coverslips, which provide resistance during this aligned contraction in a manner potentially similar to systems that employ posts or pillars. Results show that functional cardiac maturation can be improved by simply using a rectangular tissue geometry during the initial hiPSC encapsulation step, rather than implementing cell selection or tedious postdifferentiation manipulation.
Overall, increased tissue homogeneity was consistently seen in the rectangular geometry compared to the microisland and square geometries. During initial stages of cardiac differentiation, we saw a change in viable cell density distribution from day −2 to 7. On day −2, all tissue geometries showed uniform viable cell density distribution at both the edge and middle of hiPSC-laden hydrogels. By day 0, hiPSC colonies increased in size and remained evenly distributed throughout the hydrogels. After cardiac differentiation on day 7, there were regional differences in viable cell density for all tissue geometries. The exact causes of regional viable cell distributions are unknown; cells in the center may have died or migrated to the outer edge where they have greater access to nutrients and waste removal. 28
Due to the inherent nature of the rectangular geometry, where the middle is never as far from the edge compared to square and microisland geometries, regions of low viable cell density were smaller, resulting in more homogenous cardiac tissues over time. Tissue homogeneity is important for studying the impact of therapeutics on cardiac electrophysiology, where anomalies in cardiac tissue can result in unwanted arrhythmias and disruptions in electrical signal propagation. To further refine tissue production and alleviate these areas of low viable cell density, additional testing could be performed to produce tissues with smaller dimensions to avoid nutrient diffusion limitations, or vasculature could be incorporated to improve oxygen and nutrient distribution and waste removal. 29 While these methods add varying levels of complexity beyond the scope of this study, the knowledge gained would further narrow the focus for optimal cardiac tissue production for enhancing maturation.
Throughout long-term culture, tissue contraction progressed from initially localized areas of spontaneous beating to strong synchronous beating as the cardiac tissue became more interconnected in late-stage differentiation and maturation. Significant increases in contraction frequency and velocity were quantified from day 20 and 40. While ECTs from each geometry contracted at similar speeds, the directionality of propagation was significantly different between geometries. Microisland and square ECTs contracted from their outer edges inward, toward the center of the tissue, potentially indicating CM alignment from the periphery to the middle of the tissue. Conversely, rectangular ECTs propagated along the longitudinal axis, indicating anisotropic CM alignment and contraction propagation.
In addition, TEM imaging revealed higher myofibrillar organization and Z line formation in rectangular tissues compared to microislands. The late time point of day 140 was selected to align with prior TEM analysis of the microisland geometry, which had shown evidence of T-tubule formation. 17 Given limitations at the time of this study, additional evaluation was not possible. Future study to evaluate potential geometry-dependent differences in the timeline and extent of structural maturation is warranted. Both microisland and rectangular 3D-ECTs showed uniform calcium propagation, as shown in the representative isochrone maps. 3D-ECTs appropriately responded to point electrical stimulation up to 4 Hz at 20 V.30,31 Optical mapping data showed a higher frequency of spontaneous calcium transient propagation for microisland as compared to rectangular 3D-ECTs, which aligns with the higher contraction frequency observed for microisland 3D-ECTs.
No other significant difference between geometries for CTD50, CTD80, and calcium transient velocity (CTV) for the spontaneous activity or electrical pacing was observed. Calcium transient duration for both microisland and rectangular 3D-ECTs was comparable with other hiPSC-CM results (CTD50) at 1 Hz stimulation (∼340 ms); however, the observed 3D-ECT CTD50 was higher than that of human adult CMs (∼250 ms). 32 Nevertheless, CTV in rectangular tissues showed an increasing trend with increased frequency of external pacing, which may be a sign of advancing maturity compared to microisland tissues. Taken together, these findings demonstrate that encapsulating and differentiating hiPSCs in a rectangular geometry results in more mature ECTs, with contractile and structural properties that more closely mimic native myocardium.
The approach presented in this study is relatively straightforward to implement and has several advantages for producing 3D-ECTs in vitro. However, making more intricate tissue geometries or utilizing additional cell types requires more complex solutions like bioprinting. First, the ability to 3D print positive casts for producing custom-made PDMS molds allows for virtually unlimited design options for PDMS mold preparation. However, we are currently limited in tissue complexity by pipetting the polymer precursor solution by hand. Rudimentary geometries like microisland, squares, and rectangles are easily pipetted by hand, but achieving even polymer precursor distribution and tissue thickness in complex geometries is challenging.
Bioprinting of 3D-ECTs shows great potential for solving this problem where tissue can be printed with submillimeter resolution. 22 Second, while encapsulating hiPSCs on acrylated glass coverslips enables ease of characterization and is likely important in the observed improved cardiac maturation, it adds a limitation for tissue size and thickness. The acrylated coverslip prevents nutrient and waste diffusion from the bottom surface area of the tissue; this most likely contributes to the low viability in the center of microisland and square tissues.
Conclusions
In this study, we demonstrate the ability to impact cardiac tissue functionality by changing the initial encapsulation geometry of hiPSCs in hydrogel biomaterials. Rectangular 3D-ECTs displayed increased tissue uniformity with unidirectional contraction and evenly spaced and aligned sarcomeres, which are hallmarks of native adult myocardium. This robust yet simple encapsulation platform has the potential to be used in a range of applications, including cardiac drug development and cardiotoxicity analysis.
Footnotes
Acknowledgment
The authors gratefully acknowledge Dr. Ronald Gordon, Department of Pathology at Icahn School of Medicine at Mount Sinai, for performing transmission electron microscopy.
Authors' Contributions
The article was written through contributions of all authors. All authors have given approval to the final version of the article.
Disclosure Statement
Auburn University holds two patents related to the cardiac differentiation approach used here and one patent application has been submitted for the microfluidic cell encapsulation system employed. 1. P Kerscher*, AJ Hodge*, EA Lipke, “Encapsulation and Cardiac Differentiation of hiPSCs in 3D PEG-Fibrinogen Hydrogels.” U.S. Patent No. 9,587,221, Issued 3/7/2017. 2. P Kerscher*, AJ Hodge*, EA Lipke, “Encapsulation and Cardiac Differentiation of hiPSCs in 3D PEG-Fibrinogen Hydrogels.” Continuation, U.S. Patent No. 10301597, Issued 05/28/2019. 3. EA Lipke, Y Tian*, WJ Seeto*, “Microfluidics Device for Fabrication of Large, Uniform, Injectable Hydrogel Microspheres for Cell Encapsulation.” Provisional Patent 62/568,652, filed 10/6/2017. US Patent Application No. 16/153,095, filed 10/5/2018, published, pending.
Funding Information
National Institutes of Health (No. UL1TR003096), the Department of Education GAANN (No. P200A150075 and P200A180087), National Science Foundation (No. NSF-CBET-1743445), and Auburn University Undergraduate Research Fellowship, National Institutes of Health/NHLBI K01-HL133424 and R03-HL154286.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.