
Abstract
Satellite cells are considered to be adult skeletal muscle stem cells. Their ability to regenerate large muscle defects is highly dependent on their specific niche. When these cells are cultured in vitro, the loss of this niche leads to a loss of proliferative capacity and defective regeneration when implanted back into a muscle defect. The most important aspects of the niche will be discussed—in particular, the basement membrane, the niche's mechanical properties, its supporting cells, and the influence these features have on satellite cell activation, proliferation, and differentiation. Understanding more about the control of these satellite cell activities by the niche will facilitate their recruitment and effective deployment for regenerative medicine.
Introduction
Adult stem cells have the capability to self-renew, with a seemingly indefinite number of cell doublings. While self-renewal is functional for regenerative purposes, it is clear that proliferation and differentiation of stem cells need to be tightly regulated to prevent uncontrolled growth. An important part of this regulation is probably provided by the stem cell niche, which exerts this control through guidance of signals by the re-organizing ECM.3,7–9
Skeletal muscle is a type of tissue that typically experiences bouts of high levels of regeneration and repair and for this reason houses stem cells. There has been much debate about which cell type qualifies as the muscle adult stem cell, but the most prevalent notion is that the satellite cell or a subset of satellite cells most likely assumes this role.10–16 The anatomical location of the satellite cell (in between the sarcolemma and the BM; Fig. 1), its capability of self-renewal,15,17–19 its regenerative capacity in vivo, 15 and plasticity in vitro12,20–22 show that the satellite cell possesses all the requisite characteristics of a skeletal muscle stem cell. 23 In case of skeletal muscle injury or another type of stimulus, satellite cells are activated and become proliferating myoblasts. If necessary, they migrate to the designated site where they differentiate and fuse with existing or damaged fibers or form new fibers by fusing with other myoblasts.

(
However, an important and poorly understood limitation of the satellite cell is its in vitro proliferative capacity: After isolation, satellite cells can only divide a small number of times. 24 On the other hand, in vivo, a small amount of tissue-resident satellite cells is sufficient to regenerate large parts of muscle tissue.15,25 The inability to recapitulate the proliferative capacity in vitro is probably due to loss of the highly specific niche that normally surrounds these cells. 26
Satellite cells are commonly defined according to their anatomical location. 27 However, they have been shown to be a heterogeneous population according to their expression of molecular markers, suggesting varying roles for the different subpopulations in the regenerative process.28–36 A number of markers have been proposed that should distinguish the entire population such as M-cadherin, 29 CD34, 33 c-met, 29 and Pax7. 37 Evidence exists for the presence of a small stem cell–like population within the satellite cell compartment,15,19,38 indicated by the observation that some myoblasts do survive after injection into injured host muscle and are capable of robust regeneration, albeit only at the site of injection. 39
Stem cells are the ideal candidates to boost regeneration after extensive injury or to substitute a defective repair mechanism. The oldest regenerative therapy proposed is myoblast transfer therapy (MTT) 40 in which isolated myoblasts are cultured in vitro and then injected into muscles of compromised living recipients. 41 The first MTT studies focused on the restoration of dystrophic muscle in a mouse (mdx) model of Duchenne muscular dystrophy (DMD)39,42,43 and later on in DMD patients.44–48 Unfortunately, most injected myoblasts do not even survive the first hour after injection and do not migrate from the site of injection, resulting in failure to restore function.39,43,49,50 When myoblasts are injected in a fibrin clot, 51 survival does not improve. However, when satellite cells are isolated without enzymatic digestion of the muscle fiber 15 and injected without further culturing in vitro, regeneration of damaged muscles is successful.15,25,52,53
More recently, tissue engineering using primary cells54–60 or cell lines55,61–63 and standard matrices such as collagen and poly (glycolic acid) (PGA)/poly-L-lactic acid (PLLA) has been studied for regenerative purposes, but with limited success.64–68 However, implantation of myoblasts seeded in an decellularized muscle matrix gives long-term repair. 65 Current studies on the optimal biochemical and physical conditions of the scaffolds to support myoblast survival and differentiation are aimed at improving this therapeutic platform. The studies are based on the premise that the environment should resemble the natural recipient environment as much as possible to fully support differentiation and maturation of myoblasts into adult skeletal muscle.
Finally, understanding these niche principles could also facilitate stimulating satellite cells in situ for skeletal muscle regeneration.
In this review, we provide an integrated view on the role of the most important aspects of the satellite cell niche (the BM, mechanical properties, and supporting cells) and their putative pathways to control stem (satellite) cell activation, proliferation, and migration in regenerating skeletal muscle (Fig. 2).
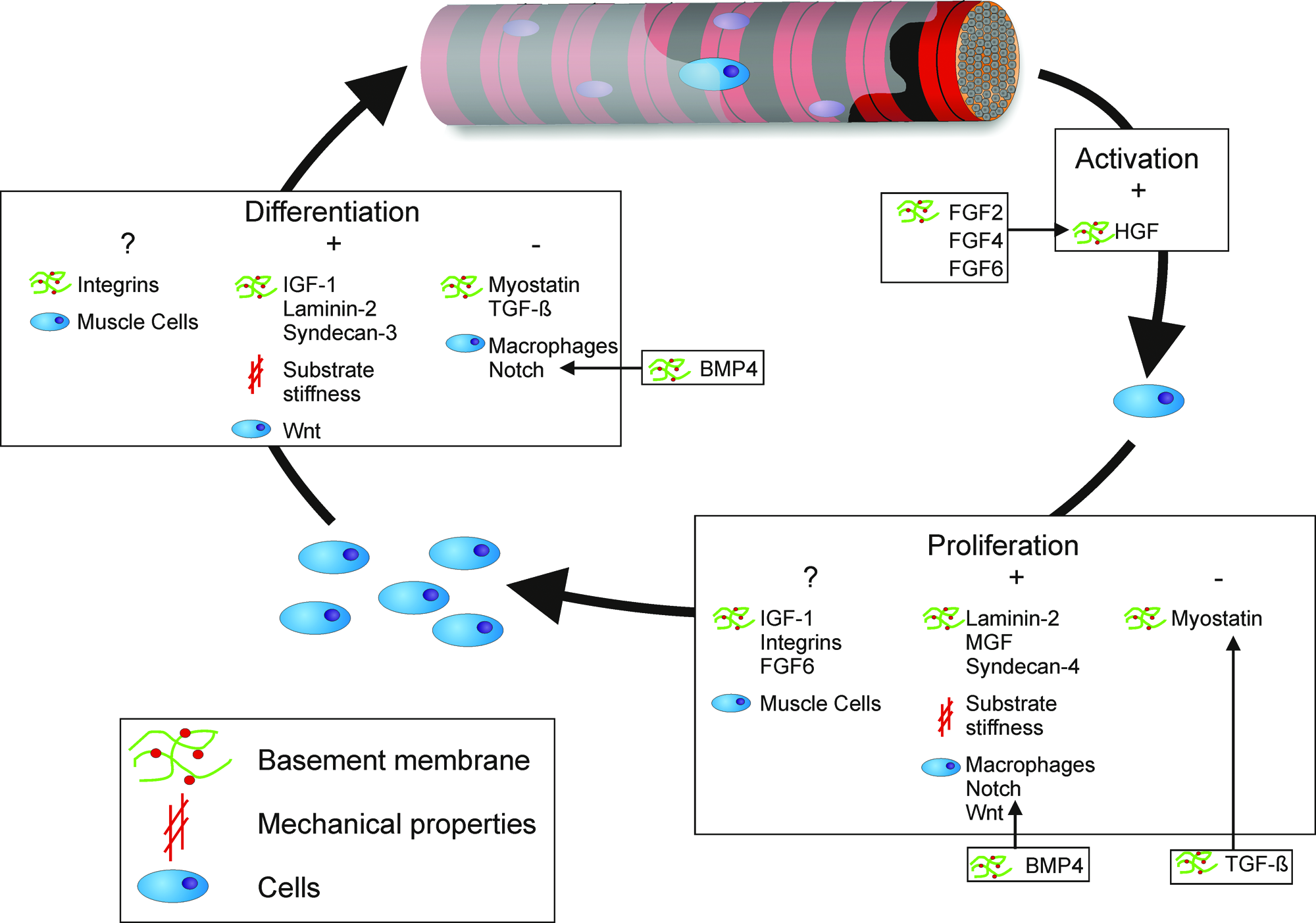
Overview of the most important components of the satellite cell niche and their influence on activation, proliferation, and differentiation. Color images available online at www.liebertonline.com/ten.
Satellite Cell Niche
A stem cell niche is commonly defined as “a specific location in a tissue where stem cells can reside for an indefinite period of time and produce progeny cells while self-renewing.” 69 Most niches contain a BM, to which the stem cells attach. 7 At either site of the BM, permanent so-called supporting cells can be localized that are of importance to stem cell functioning. 7 The satellite cell niche indeed contains a BM and muscle fibers, 10 and/or endothelial cells 70 are close by and could function as supporting cells. However, although they have been shown to have an effect on proliferation of myoblasts in vitro, endothelial cells have not been shown to be in direct contact with satellite cells. In most stem cell niches (e.g., in the intestine), the supporting cells are small cells that are located at the opposite site of the BM, but cases exist (e.g., in the testis, reviewed by Wong et al. 71 ) in which stem cells are supported by big cells located at the same site of the BM. Therefore, the muscle fiber is the most promising candidate as the satellite cell niche's supporting cell. Alternatively, in the Drosophila midgut, stem cell niches were described that do not rely on supporting cells for their function, 72 opening the possibility that the satellite cell niche exists without any supporting cells. This hypothesis is reinforced by the fact that satellite cells with or without the parent fiber regenerate defects similarly after transplantation, as long as the isolation methods are optimal. 15
Another prerequisite of a stem cell niche is that when it is depleted of stem cells, it should persist and be able to house new stem cells. 73 Bone marrow stem cells, for example, can enter and leave the circulation to occupy empty niches in a process called homing (reviewed by Whetton and Graham 74 ). Not only satellite cells25,30 but also bone marrow cells30,70,73 have been shown to be able to occupy empty satellite cell niches, satisfying also this requirement.
When the satellite cell niche is compared to niches in other common stem cell systems, such as skin and intestine, the most obvious difference is that satellite cells in their niche are quiescent, whereas in most systems stem cells are constantly active to replace cells that are lost due to daily wear and tear. Usually, an intermediate cell called transit-amplifying (TA) cell takes care of actual expansion, whereas the stem cell only continuously replenishes this TA population. In the intestine, for example, stem cells at the bottom of crypts constantly divide to give rise to TA cells that move up and differentiate to ensure a steady flow of cells that are shed at the surface (reviewed by Yen and Wright 75 ). However, in other tissues, niches do exist that contain quiescent stem cells. In the heart, for example, cardiac stem cells (CSCs) are thought to take care of regeneration (reviewed by Leri et al. 5 ). It is hypothesized that CSCs are quiescent cells, surrounded by a BM, that give rise to a TA population after activation, which is then responsible for actual repair. 5
Possibly, such a TA population also exists in the muscle stem cell system. 36 It has been shown that satellite cells are a heterogeneous population containing a small stem cell compartment,10,28,29,36,76 which could mean that the rest of the satellite cells should be perceived as TA cells. Evidence for this theory comes from elegant studies showing a hierarchical lineage relationship of satellite cells.77,78 Alternatively, all satellite cells should be considered TA cells that descend from a common precursor stem cell located elsewhere, for example, in the interstitial spaces of skeletal muscle.36,78 However, observations that satellite cells self-renew15,18,79 and regenerate muscle 15 clearly show that they are more than just TA cells. 23
Basement Membrane
The BM is a specialized sheet of connective tissue. Its most important constituents are type IV collagen, laminin, and heparan sulfate carrying proteoglycans (HSPGs). A laminin network faces the muscle fiber, complemented by a network of collagen IV. These proteins are linked by entactin-1 (also called nidogen-1) to form a complex structure containing a high number of binding sites, for example, for proteoglycans and integrins (Fig. 1). In skeletal muscle, the interstitial fibroblasts are responsible for the production and excretion of ECM molecules. During development and regeneration, mononuclear muscle cells also synthesize and remodel ECM, whereas the contribution of the adult muscle cell is relatively small. 80 Homing, residence, and activation of stem cells can be regulated through differential expression of integrins and tissue-specific laminin isoforms in the BM.81–84 Integrity of the BM is crucial to keep the stem cells physically in place; defects or regulated gaps in the BM will allow cells to move through the tissue. 85
Integrins are involved in linking the BM to the intracellular cytoskeleton 86 and consist of a large family of cell surface receptors composed of an α and a β subunit. They act as signal transducers after binding to the appropriate ligand, influencing cell migration, cell shape, and cell–cell interactions.80,82,87 Integrin α7 is the main isoform in mature skeletal muscle.88,89 It binds muscle fibers to laminin and dystroglycan and is upregulated during regeneration.90,91 Myoblasts and newly formed myotubes in vitro express integrin α5, 88 and together with integrin β1 it forms the main receptor complex for fibronectin, which is also present in the BM.92,93 Integrin α3 is found on quiescent satellite cells and myoblasts and plays a role in migration and differentiation by forming complexes with the integrin β1 subunit and A disintegrin and metalloproteinase 12 (ADAM12) (see end of this section).31,94 Integrin β1 plays a role in laminin binding of myoblasts in vitro.89,93,95 While these integrins are of obvious importance for mechanical coupling and deficient mice develop different types of muscular dystrophy, a role in regeneration remains speculative.31,96,97
One of the BM components that is specific for muscle and therefore is likely to play an important role in the niche functioning is laminin-2. Laminins are heterotrimers composed of α, β, and γ subunits that form a network-like structure in BMs (Fig. 1). Laminin-2 is composed of α2, β1, and γ1 proteins and is also referred to as merosin. 98 The laminin α2 subunit is the muscle-specific part of laminin-2 and is connected to muscle fibers through integrin α7β1 and dystroglycan 99 in a large dystrophin-associated protein complex.85,100,101 The significance of laminin α2 for satellite cell function is underscored by the phenotype of the laminin α2 knockout mouse. Not only is the skeletal muscle BM almost completely absent in these animals 102 leading to a decrease in the relative amount of satellite cells, 103 during regeneration the amount of myoblasts is additionally decreased due to reduced proliferation rates 103 and increased apoptosis. 102
Multiple proteoglycans can be found in the ECM in skeletal muscle, but only a few seem to be important during regeneration. Proteoglycans consist of glycosylated core proteins with one or more covalently attached sulfated glycosaminoglycan chains. Quiescent satellite cells express syndecan-3 and -4, which are transmembrane HSPGs that become upregulated upon activation. They only differ in their extracellular domains 104 and are required to transduce signals 105 directly through binding to signaling molecules or by presenting them to their specific receptors. 106 Syndecan-4 has been implicated in fibroblast growth factor (FGF) signaling107,108 and seems to be required for early satellite cell activation and proliferation. Satellite cells in knockout mice express reduced levels of syndecan-3 and c-met, and the BM appears disorganized in vivo. When muscle damage is induced in these animals, regeneration fails and results in nonfunctional myofibers and scar tissue. 109 In cell culture, proliferation of myoblasts is delayed and differentiation does not even take place. 109 In syndecan-3−/− mice, the number of satellite cells and myonuclei is increased in muscles in spite of normal size, location, and gene expression profile of satellite cells and intact BM. When cells isolated from these mice are cultured, differentiation is abnormal, which is evident by aberrant fusion into syncytia instead of fibers. 109 Therefore, it seems that integrity of the BM and the ability of cells to bind to the BM (for instance through HSPGs) is essential for regeneration.
In addition to laminins and HSPGs, growth factors play an important role in the regeneration process. They can be produced and secreted by muscle cells or immune cells and can be liberated from the ECM where they are bound to proteoglycans. 110 Growth factors that are involved in the regeneration program include hepatocyte growth factor (HGF), members of the transforming growth factor-β (TGF-β) superfamily, FGFs, and insulin-like growth factor-1 (IGF-1) isoforms. HGF is a heparan-binding protein that activates quiescent satellite cells through its receptor c-met111–113 and is normally present in an active form in the ECM of uninjured muscle. 114 HGF is released by matrix metalloproteinases (MMPs)114,115 after stretch or other types of injury 116 and can be produced by satellite cells in vitro. 117 Next to HGF, FGFs are involved in the regulation of regeneration. FGFs 2, 4, and 6 stimulate proliferation of myoblasts in vitro, 118 but only in the presence of HGF. FGF2 can be secreted by infiltrating macrophages 119 and is found to be upregulated in regenerating muscle 120 together with FGF6, which is thought to be secreted by injured muscle fibers.121,122 FGF6 knockout mice show a severe regeneration defect that is most likely caused by disturbed activation or proliferation of satellite cells. 123 Migration of FGF6−/− myoblasts is also impaired. 124 IGF-1 isoforms resemble FGFs in their actions. At least four different isoforms exist, two of which play a role in muscle growth and repair. 125 Mechano growth factor (MGF), which is induced during the initial satellite cell activation, 126 especially after mechanical stimulation 127 and damage, 128 stimulates proliferation of myoblasts. 129 After this initial phase, MGF is replaced by systemic IGF-1, through alternative splicing.128,130 Systemic IGF-1 has been shown to induce faster differentiation in C2C12 myoblasts 131 and increased regeneration in mice. 132 In IGF-1 transgenic mice, myoblast proliferation and differentiation is augmented, 133 whereas IGF-1 knockout mice die at birth because of severe muscular dystrophy. 134 In contrast to IGFs, several members of the TGF-β superfamily, including TGF-β, bone morphogenetic proteins (BMPs), and myostatin, are involved in negative regulation of regeneration by inhibiting proliferation or differentiation. TGF-β135,136 and myostatin137,138 reduce myoblast recruitment and differentiation, and TGF-β also remodels and repairs ECM and BM. 139 In a number of known stem cell niches (e.g., in neural crest 140 ), BMPs prevent stem cell proliferation, which can be counteracted by upregulation of Noggin.141,142 In C2C12 myoblasts and satellite cells, BMP4 in combination with Notch signaling can block myogenic differentiation. 143
To get to the site of injury for repair, myoblasts have to migrate through the ECM. MMPs are involved in degrading ECM components, which not only enables myoblast migration but also leads to release of and exposure to cytokines and growth factors that target myoblast proliferation and differentiation.94,144–147 MMP2 and 9 can be secreted by satellite cells148,149 and are upregulated in muscle during injury and regeneration.150–152 They degrade not only collagen IV but also other BM components such as dystroglycan, 85 and are secreted into the ECM in an inactive form that is activated after cleavage. Inhibition of these MMPs prevents migration of myoblasts in vitro,148,153 and when injected in vivo, migration of myoblasts is triggered. 147 ADAMs are cell surface receptors involved in regeneration by mediating adhesion and transmembrane signaling. ADAM12, also called meltrin-α, is expressed during muscle development and regeneration.31,154,155 It is also expressed in activated satellite cells 156 and binds to integrin α7β1, 93 integrin α9β1,157–159 integrin α3β1, 31 and cell surface syndecans (probably syndecan-4), leading to integrin-dependent spreading of cells. 160
Obviously, the BM plays a crucial role in maintaining the stem cell function of satellite cells. However, it is hard to consider the effects of integrins, different matrix proteins, growth factors, and MMPs individually because they all play interconnected roles in the complex signaling pathways between the satellite cell and its environment.
Mechanical Properties
It is clear, although under-appreciated, that the mechanical properties of the matrix greatly affect cellular phenotype. Examples are surface tension–induced cellular organization during embryonic development 161 and locomotion. 162 In muscle cells (cyclic) stretch has been shown to induce hypertrophy 163 and protein expression of myogenic regulatory factors 164 and IGF-1 splice variants, 127 activate satellite cells,116,165,166 and improve tissue-engineered muscle constructs.60,167–169
Cells can feel their surroundings by anchoring and pulling with their cytoskeletal proteins, integrins, and other molecules that mediate adhesion to the ECM. 170 Recently, the stiffness of the substratum that cells are cultured on has gained interest. Mesenchymal stem cells, for example, differentiate into the neuronal, muscular, or osteogenic lineage when cultured on substrates with a stiffness of 0.1–1 kPa, 8–17 kPa, and 25–40 kPa, respectively. After 1 week on such a matrix, differentiation pathways seem to be fixed and can no longer be reprogrammed with specialized media. 171 For skeletal muscle cell differentiation and especially maturation into mature, striated skeletal muscle, the stiffness of the environment is equally important172,173 (personal observations). Muscle has been shown to possess a Young's modulus of about 12 kPa in rest when measured in the transverse direction (the direction of adhesion of cells), which is the same as for differentiating skeletal muscle cells in vitro. 174 Differentiation and maturation of myoblasts have been shown to be optimal on this stiffness. When C2C12 murine myoblasts were cultured on gels with different Young's moduli, cross-striation, which is an indicator of muscle maturation, only occurred on gels of intermediate stiffness (8 and 11 kPa) 173 (Fig. 3) or on top of a layer of myotubes (Young's modulus, 12–15 kPa). 173 Probably, the proliferative capacity of cells 175 is also influenced by substrate stiffness (personal observations), which could partly explain the discrepancy between the in vivo and in vitro proliferative behavior of satellite cells. Primary myoblasts were shown to proliferate at a higher rate on stiffer gels (12 and 45 kPa) compared to softer gels (1 kPa). 175

Cross-striations in myotubes depend on substrate stiffness. Two or 4 weeks after plating cells on collagen-coated polyacrylamide gels of different stiffnesses, only myotubes on gels of intermediate stiffness showed cross-striations of myosin. Bars: 20 μm. Reproduced from The Journal of Cell Biology, 2004, 166:877–887. © 2004 The Rockefeller University Press. Color images available online at www.liebertonline.com/ten.
This aspect deserves further exploration because it is likely to be of importance not only for standardization of cell culture experiments but also for the design of matrices that support cell therapies in regenerative medicine. In terms of mechanical properties, for example, the recipient environment in patients that qualify for MTT is very different from the required environment for myoblast proliferation and differentiation. In mdx mice and Duchenne patients, extensive scar formation replaces the affected muscles, and this fibrotic tissue might prevent myoblasts from surviving and regenerating the diseased muscle, not only by interfering positionally, but also by increasing stiffness beyond the optimal parameters for myoblast proliferation and differentiation. 40
The BM and, most importantly, its collagen/laminin network are probably responsible for determining the stiffness of muscle fibers.80,151,176 However, up till now, no mechanical tests have been performed on muscle BM. Nonetheless, not only the mechanical properties themselves but also the combination of stimulatory effects with the different BM components can have synergistic, complementary, or opposing effects on myogenesis. For example, mechanical stimulation through the laminin receptor results in differentiation of C2C12 murine myoblasts and has been shown to be mediated through β1 integrins, whereas mechanical stimulation through the fibronectin receptor encourages proliferation. 164
In summary, the mechanical properties of the satellite cell niche are almost certainly determined by the BM and are pivotal for proper stem cell function of satellite cells. However, it is not likely that the stiffness of the environment is actively regulated to trigger different processes in the regenerative process; it could therefore be considered to be an essential requirement.
Cells
Another important niche component is the cells other than the stem cells. For bone marrow for instance, these supporting cells, most notably osteoblasts, play an important role in stem cell maintenance and regulation (reviewed by Wilson and Trumpp 4 ). In the muscle stem cell niche, mature muscle cells, other mesenchymal cells, and cells of the specific and innate immune system connect with the satellite cells.
In the normal situation, satellite cells are only in direct contact with the adjacent muscle cell. The importance of this connection is underscored by the fact that the result of myoblast transplantation is better when fibers containing satellite cells are transplanted as a whole compared to transplantation of cells liberated from the fiber by enzymatic digestion. 15 However, when satellite cells are dissociated from the muscle fiber by physical trituration, their regenerative potential in vivo is comparable to transplantation of single fibers. Therefore, although the parent fiber itself does not seem to be required for satellite cell function, the ability of the satellite cell to bind and communicate to the fibers or the niche present in the recipient muscle seems to be essential and might be impaired by enzymatic digestion. 15
In the event of injury and muscle damage, cells of the immune system are also able to come into contact with the satellite cell and convey signals directing proliferation, migration, and differentiation, for example, through the production and excretion of growth factors. Macrophages are important for removal of dead cells and the dead parts of muscle fibers. They have also been shown to be able to directly stimulate satellite cell proliferation and delay their differentiation. This effect is probably mediated by FGF2, platelet derived growth factor (PDGF), or leukemia inhibitory factor (LIF).
More than 60% of the satellite cells are located close to capillaries and are thought to receive signals from endothelial cells although they are not in direct contact with them. The number of capillaries per muscle fiber has been shown to correlate to the number of satellite cells, and loss of capillaries leads to loss of satellite cells, pointing to some sort of interaction. 70 In addition, transwell experiments showed that endothelial cells have a positive effect on proliferation of myoblasts, which is mediated by growth factors. 70
Cell–cell interactions have been shown to be pivotal for muscle regeneration and can be mediated by components of the cell-bound receptor–ligand complexes of the Notch and Wnt family. Wnt signaling plays a role in stem cell determination 177 in different stem cell niches (e.g., the intestinal stem cell niche 142 ) and is also thought to activate stem cells during muscle regeneration.8,178 The canonical Wnt pathway is mediated by β-catenin, which translocates to the nucleus where it induces transcription of specific genes involved in cell proliferation and survival together with transcription factors of the Tcf/Lef family. Overexpression of β-catenin increases proliferation and induces hypertrophy in vitro in C2C12 myoblasts and increases regeneration and the number of satellite cells in vivo in mice. 179
Notch is an evolutionary conserved heterodimeric transmembrane receptor that is involved in cell fate control by local cell interactions. Most of the ligands involved in Notch signaling are also membrane bound and interact with Notch receptors on adjacent cells, leading to a signaling cascade that regulates the transcription of specific genes involved in self-renewal, proliferation, and differentiation of stem cells together with functional Wnt signaling.180–183 During muscle regeneration, satellite cells are activated and start proliferating due to Notch signaling, through the ligand Delta-1.32,76,184–186
Different types of coculture experiments have been performed to investigate the effect of contact with a muscle environment on different types of cells. In general, it seems to be the case that direct contact with either muscle fibers or myoblasts is essential for myogenic conversion.187–191
Overall, the ability to bind to and be in direct contact with muscle fibers seems to be essential for the regenerative potential of satellite cells. Different cell–cell interaction and signaling molecules seem to play an important role in this process, although we need more direct results to explain some of the contradictory roles attributed to these molecules.
Discussion
Satellite cells from skeletal muscle are identified as correlate of a tissue-resident stem cell; that is, they are capable of in vivo tissue regeneration.10,12,15,18,22 Since satellite cells are easily identifiable and can be harvested from intact muscle, therapeutic applications for MTT or for tissue engineering have been envisaged. However, it appears that the myoblast population that is harvested from adult skeletal muscle quickly loses its self-renewal capacity during in vitro expansion.27,192,193
Pivotal evidence suggests that the isolation procedure and/or in vitro expansion causes this fate change; enzymatically isolated or cultured myoblasts have been shown to lose their regenerative capacity, whereas direct implantation of nonenzymatically isolated myoblasts or single fibers with resident satellite cells is extremely effective to regenerate damaged muscle tissue. 15 In addition, when sliced muscle grafts are used in muscle transplantation, cells survive for up to a year.194–196
We suggest that preservation of critical stem cell niche components is important for maintaining the regenerative capacity of satellite cells. We have reviewed the BM contribution to this niche. Appealing candidates are muscle-specific laminin isoforms 81 and syndecans 3 and 4. 109 However, systematic studies addressing the essential components for the satellite stem cell niche are needed. Once identified, reconstitution of these components during cell culture might improve the applicability of satellite cells in regenerative medicine. An under-appreciated aspect of the stem cell niche is the mechanical framework it provides. There is now increasing evidence that cells sense the mechanical properties of their matrix and respond by phenotypic change,171,173 possibly by differentiating away from their precursor state (e.g., in the case of rigid culture plastic). Support for the importance of the matrix in the stem cell niche also comes from observations that BM integrity during injury accelerates natural healing.84,197 Not only loss of the matrix context but also loss of the cellular context may cause fate change of the satellite cell. Experience with cocultures of cells of different origins with adult muscle fibers or myoblasts strongly suggests that physical contact of stem cells with differentiated muscle favors muscular differentiation.187–191
In addition, the isolation procedure might select nonregenerative subpopulations of satellite cells. 79 Different subpopulations of stem cells in skeletal muscle appear to exist, and it is very likely that some have more regenerative capacity than others.15,19,38 Clearly, detailed marker studies complemented by clonal analyses are required to dissect the importance of these subpopulations and the extent to which the isolation procedure affects their presence and performance.
Elucidating the microenvironmental needs of satellite cells will have considerable implications for the use of these cells for regenerative medicine. In addition, the two approaches, MTT and tissue engineering, will likely differ in their requirements concerning proliferative and migratory capacity. The indications and therefore the recipient characteristics, for instance large defects in tissue engineering versus generalized dystrophy for MTT, will be different as well.
For both therapies to advance it is therefore essential that we understand the biochemical, cellular, and mechanical cues that promote satellite cell proliferation and differentiation in vitro and in vivo. The current evidence provides a sound basis for systematic studies of these cues in the setting of regeneration of skeletal muscle.
Footnotes
Disclosure Statement
No competing financial interests exist.