
Abstract
Surface-induced thrombosis is a significant issue for artificial blood-contacting materials used in the treatment of cardiovascular diseases. The development of biomaterials and tissue-engineered constructs that mimic the vasculature represents a way to overcome this problem. Elastin is an extracellular matrix macromolecule that imparts arterial elasticity where it comprises up to 50% of the nonhydrated mass of the vessel. In addition to its critical role in maintaining vessel integrity and elastic properties under pulsatile flow, elastin plays an important role in signaling and regulating luminal endothelial cells and smooth muscle cells in the arterial wall. Despite its well-established significance in the vasculature and its growing use as a biomaterial in tissue engineering, the hemocompatibility of elastin is often overlooked. Past studies pointing to the potential of arterial elastin and decellularized elastin as nonthrombogenic materials have begun to be realized, with elastin scaffolds and coatings displaying increased hemocomptibility. This review explores the mechanisms of elastin's nonthrombogenicity and highlights the current problems limiting its wider application as a biomaterial. We discuss the benefits of constructing biomaterials encompassing the relevant mechanical and biological features of elastin to provide enhanced hemocompatibility to biomaterials.
Thrombosis of Vascular Devices
Biomaterials and Thrombogenicity
The development of a truly nonthrombogenic biomaterial has been elusive due to the complex nature and interplay of coagulation, thrombosis, and inflammation that occurs after implantation of foreign materials. Understanding these processes will aid the development of compatible materials.
Classical coagulation pathways
Thrombus formation in vivo results from two enzyme cascades, the extrinsic and intrinsic pathway (Fig. 1). The extrinsic pathway is initiated by the activation of tissue factor following the physiological response to diseased or damaged endothelium and exposure of blood to the subendothelium (Fig. 1, green section, bold text). The intrinsic pathway is activated by contact with implanted materials and subsequent conformational changes in plasma proteins (Fig. 1, blue section, regular text). Ultimately, these pathways converge on a common pathway, which is defined by the proteolytic conversion of prothrombin to active thrombin (Fig. 1, yellow section, italic text). Thrombin catalyzes the conversion of fibrinogen to fibrin, which in turn spontaneously forms crosslinked fibrils resulting in a fibrin clot. 9

Modified coagulation cascade. An overview of the intrinsic, extrinsic (bold), and common (italics) coagulation pathways. The factors that platelets activate are denoted by circles (), and the factors that various complement proteins activate are denoted by open squares ( ) (modified from Refs.13,14,22). Color images available online at www.liebertonline.com/teb.
) (modified from Refs.13,14,22). Color images available online at www.liebertonline.com/teb.
Platelets
Platelets are the cellular component of coagulation. They are 3–5 μm anuclear cells that can adhere to foreign surfaces or damaged vasculature and form aggregates through a cohort of platelet-specific adhesion receptors and integrins. For example, platelets can be activated by adhesion to a variety of proteins in the vascular wall such as collagen, fibronectin, laminin, and vitronectin.10,11 Additionally, coagulation enzymes from both pathways such as thrombin and factor X (FX) directly activate platelets, as does turbulent blood flow caused by implanted material or atherosclerotic plaques. Once activated, platelets contribute to the fibrin clot leading to thrombus formation. Activated platelets in turn feedback and activate various coagulation enzymes (circles in Fig. 1). 9
Thrombosis in relation to biomaterials
Traditionally, it was thought that the two pathways were independent, and that blood contact with biomaterials specifically activated the intrinsic but not the extrinsic pathway. 9 According to this model, albumin and fibrinogen adsorption onto the biomaterial surface after implantation triggered the intrinsic pathway response. This trigger is proposed to be due to localized denaturation of these proteins, which exposes domains that in turn lead to activation of kallikrein and FXII. This effect can be exacerbated by surface charge: negatively charged surfaces directly activate FXII, whereas net positively charged surfaces can bind and activate platelets 12 (boxed regions in Fig. 1).
However, as research into material-driven coagulation continues to accumulate, these traditional views are being challenged. There is now evidence that the intrinsic and extrinsic pathways are not exclusive and that activation of the complement pathway (which leads to inflammation) plays an important role. For example, it has been shown that implanted materials cause tissue factor activation and therefore activate the extrinsic pathway (dashed lines in Fig. 1). 13 Additionally, tissue factor can activate FIX, originally thought to be involved only in the intrinsic pathway.13,14 Complement factors are activated by surface-bound albumin and fibrinogen through FXIIa, kallikrein, and thrombin. Activation of complement leads to activation of platelets, coagulation enzymes (open squares in Fig. 1), and inflammation through leukocyte activation.13,14 The extent of activation of each of these pathways determines the overall thrombogenicity of a blood-contacting material.
Conventionally, biomaterials were selected on the basis of their perceived relative inertness and empirical assessment of a minimally elicited tissue response. However, due to the growing appreciation of the complex, multiple pathways that contribute to thrombosis, the failure of this approach to achieve successful clinical outcomes has led to a new paradigm. This encompasses the need for specific biomaterial-initiated physiological responses that are increasingly based on the location and environment of the implant. 15 The integrated use of human proteins with medical implants is being explored as a way to develop this approach. Extracellular matrix proteins in the vascular wall such as collagen, fibronectin, laminin, and vitronectin are known to activate thrombosis through the activation of platelets.10,11 Although these molecules are available in animal-derived and recombinant forms, their thrombogenicity has limited their use. A nonthrombogenic vascular compatible protein would present the opportunity to develop a range of hemocompatible biomaterials and tissue-engineered constructs.
Elastin in the Vasculature
Elastin is a dominant component of the vasculature, having both important mechanical and cell signaling roles. Tropoelastin, the precursor of elastin, is produced by endothelial cells and vascular smooth muscle cells (VSMCs) in blood vessels. In the extracellular space, tropoelastin is exposed to lysyl oxidase-mediated oxidation at exposed lysines and then self-associates by coacervation for cross-linking. 16 The growing elastic polymer is incorporated with microfibrillar proteins to form mature elastic fibers. 17 Elastin accounts for ∼50% of the dry weight of the arterial wall where it is confined to specific locations based on its function. 18 Elastic arteries are comprised of three layers, the tunica intima, media, and outermost adventitia (Fig. 2). The adventitia is characterized by elastic fibers interspersed with collagen, providing mechanical strength, recoil, and resilience. The media is predominantly comprised of alternating layers of elastin-rich lamellae interspersed with smooth muscle cells, which are responsible for maintaining blood pressure during pulsatile flow.19–21 Of particular relevance to the potential for blood contact, a distinct continuous sheet of elastin forms the internal elastic lamina, providing a surface on which endothelial cells form a monolayer and separating the intima from the tunica media (Fig. 2). 22
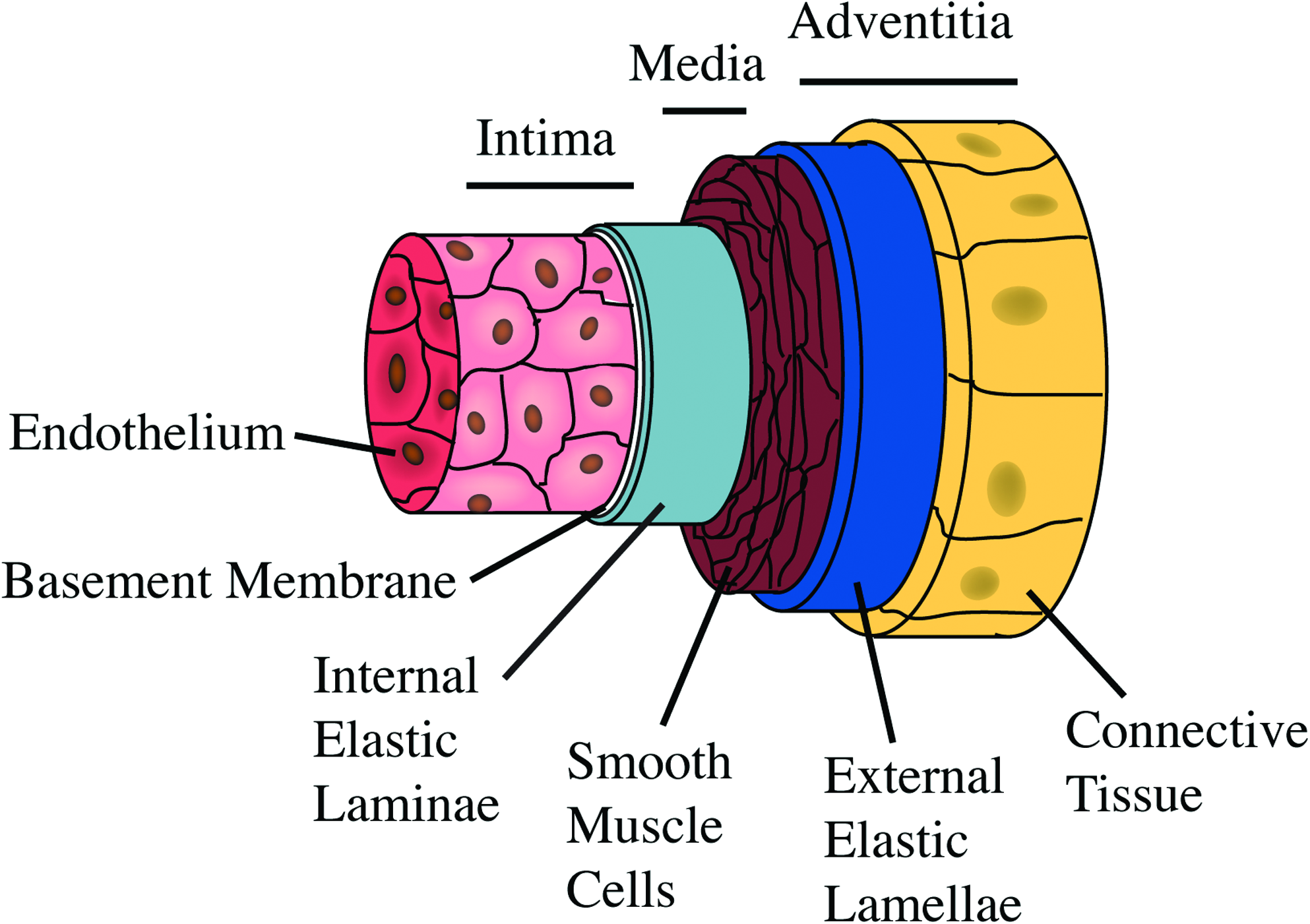
Elastic artery schematic. Schematic representation of an elastic artery showing the three layers and major components of these layers (modified from Ref. 22 ). Color images available online at www.liebertonline.com/teb.
Elastin interacts with many cell types and causes a variety of cellular responses.23–25 The major cell types in the vasculature are endothelial cells present in the tunica intima, and VSMCs present in the tunica media. Studies of the elastin knockout mouse revealed that elastin is essential for arterial morphogenesis and for controlling smooth muscle cell proliferation. The null mutation is lethal with the mice dying from obstructive arterial disease, caused by the uncontrolled proliferation of VSMCs, by day 4 postpartum. 26 Elastin inhibits VSMC proliferation by inducing a quiescent contractile phenotype through its VGVAPG motif.26–29 In contrast, various forms of elastin interact favorably with endothelial cells and support their growth.30–33 Elastin peptides are chemotactic for endothelial cells 34 and are consequently pro-angiogenic. 35 These properties are not readily utilized because elastin peptides are also chemotactic for additional cell types, including fibroblasts and monocytes and can lead to proinflammatory responses. 36 However, intact elastic fibers have an opposite effect, demonstrating anti-inflammatory properties. 37
In addition to modulation of cell signaling, tissue-engineered elastin constructs and elastin-based materials have the potential to mimic the essential mechanical properties of major vessels. Synthetic elastin scaffolds have been shown to be structurally similar to native elastic fibers (Fig. 2 in Ref. 25 ) and to have comparable elasticity, demonstrating a youngs modulus of 220–280 kPa compared with 300–600 kPa for native elastin. 25 Further incorporation of synthetic elastin into a vascular conduit facilitated the close matching of the conduits compliance to that of a native artery (320 ± 25 kPa vs. 267 ± 46 kPa for the native). 38 In addition to this work elastin and elastin-like materials have been coated onto surfaces, made into hydrogel materials and spun into fibers.21,39 However, the broad use of naturally occurring elastin is limited due to the insolubility of the mature cross-linked biopolymer and the necessity to purify it from animal sources. Elastin-rich tissues such as large arteries and bovine ligamentum nuchae can be treated with solvents, cyanogen bromide, denaturing agents and selected proteases to remove the cellular and extracellular matrix components, leaving the insoluble elastic network intact. However, to utilize elastin in biomaterial construction it must be rendered soluble by acid hydrolysis or enzyme degradation. 40 Alternatively, intact full length material in the form of recombinant human tropoelastin can be purified from substantial overexpression 41 and has been used to construct a diverse range of materials. 42 Additionally, synthesized elastin-like polypeptides are increasingly being used for construction of biomaterials 43 as well as for a range of medical applications. 44 This review assesses the existing knowledge of these various forms of elastin and their interactions with blood.
Thrombogenicity of Elastin
The interactions of elastin with components of the blood have not been fully characterized; however, key research points to elastin being nonthrombogenic. 45 Research before the 1980s revealed that elastin isolated directly from blood vessels had anti-thrombogenic properties.46,47 Soluble bovine elastin and elastin directly purified from rabbit aorta had no reaction with platelets from heparinized platelet-rich plasma (PRP). 46 This was surprising given the profound thrombogenicity of other vascular matrix components such as collagen. 47
Continuing these initial findings, an in situ perfusion system was used to study platelet adhesion and platelet thrombus formation on components of the vessel wall. 48 Rabbit arteries were either intact or decellularized to varying degrees to reveal different vessel components on the surface. Inverted arteries displaying adventitial collagen, or endothelium denuded arteries displaying the basement membrane, caused the most platelet adhesion. Arteries digested with collagenase, exposing the microfibrils of the subendothelium were less thrombogenic. Arteries displaying elastin, remaining intact after collagenase and trypsin digestion, showed the least amount of platelet adhesion. 48 Further studies showed that elastin isolated from rabbit or pig aorta caused minimal platelet adhesion and no spreading, aggregation, or degranulation, which is characteristic of platelet activation.49,50 Later research confirmed that the elastin component of elastic fibers is responsible for this lowered thrombogenicity, whereas the microfibrillar components were shown to cause platelet activation, though to a much lower extent than collagen. 51
These findings were extended in studies of exposed canine femoral arteries in vivo. The arteries were treated proteolytically to remove the endothelium. The now-exposed elastin layer caused a substantial delay in thrombus formation, with thrombus occurring from 7 to 35 days compared to arteries that were injured to destroy the elastic lamina that thrombosed within 48 h. 52 Overall, this research demonstrated that elastic fibers from decellularized vessels are the least thrombogenic protein of the major subendothelium components tested (Table 1).
ePTFE, expanded polytetrafluoroethylene.
Elastin as a Blood-Contacting Biomaterial
Given the compelling nature of the experiments to date, researchers increasingly sought to adapt elastin for use as a biomaterial. There is great potential to utilize its unique elastic mechanical properties to construct synthetic vascular grafts or take advantage of its signaling properties as a coating for vascular devices. Examples of elastin being used as a biomaterial are discussed below with emphasis on the thrombogenicity of the materials.
The first indication that elastin had the potential to be used to construct nonthrombogenic biomaterials came from in vivo implantation of elastin scaffolds. Elastin purified from porcine common carotid arteries was used to construct a stent graft that placed elastin at the site of an injured vessel to reduce VSMC hyperproliferation. Although hemocompatibility was not the main end point of this study, there was no observed thrombotic or inflammatory response. 29 Additionally, scaffolds of elastin purified from rat aortas were implanted back into rat as abdominal grafts. These had been prepared by treatment with alkali to reveal distinct vascular wall surfaces. This approach showed that the elastin matrix caused the least leukocyte and platelet adhesion compared to basal lamina or adventitial collagen. 53
There have been two studies in which elastin scaffolds were constructed into vascular grafts to directly assess their in vivo performance. The first study utilized purified elastin and collagen scaffolds obtained from porcine aortas and common carotid arteries. The elastin scaffolds showed decreased platelet adhesion and aggregation compared to collagen scaffolds in vitro using PRP. When implanted in a rabbit carotid artery, the elastin-based scaffolds displayed substantially reduced platelet adhesion. 54 The second study utilized grafts constructed from purified porcine arterial elastin and fibrin-bonded layers of acellular small intestinal mucosa, used in a porcine carotid artery interposition model to directly test acute thrombogenicity. The elastin-containing grafts remained patent for significantly longer and only had thrombosis associated with the suture line, compared to commercially available expanded polytetrafluoroethylene, which had thrombus along the full length of the graft. 55 This research was particularly significant because it was the first to show a reduction in acute thrombus formation in vivo (Table 1). These animal studies support the logic of using elastin in the construction of vascular grafts, taking advantage of the properties of elastin to provide essential mechanical properties to mimic native arteries structurally and provide a nonthrombogenic luminal surface. Indeed, this approach has been employed to construct a vascular graft incorporating recombinant human tropoelastin and polycaprolactone, with mechanical properties matching a human mammary artery and a 100% elastin luminal layer. The elastin-rich lumen enhanced endothelial cell attachment and proliferation and had low platelet interactions in vitro. 38
While the mechanism for elastin's nonthrombogenicity has not been fully elucidated, research into the thrombogenicity of other matrix proteins is illustrative of potential mechanisms. Fibronectin and collagen, both thrombogenic matrix proteins, specifically bind multiple platelet integrins.10,56 Elastin and derivatives do not contain an integrin binding arganine, glycine, aspartic acid (RGD) sequence 24 and this lack of an RGD sequence could also account for its low platelet binding ability. A second consideration is that platelets can bind through carbohydrate modifications on proteins. 57 Elastin is not post-translationally modified with carbohydrates. 58 Similarly, albumin displays low thrombogenicity and contains very little carbohydrate modification.57,59 It may be that a paucity of glycosylation works to elastin's advantage by reducing its platelet binding capacity.
Another potential mechanism involves inhibition of thromboxane B2-induced platelet aggregation. Soluble elastin and elastin fragments inhibited thromboxane B2 in nonactivated and thrombin-activated platelets, though high concentrations of elastin were used. 60 A mechanism of thromboxane B2 inhibition is suggested to be by a decrease of free cytosolic Ca2+ in platelets. 61 However, how elastin could decrease intracellular platelet calcium was not explored.
As it is known that insoluble elastin binds calcium, 62 calcification could be a concern in the context of vascular biomaterials. Elastin abnormality in diseased arteries is one of many factors that contribute to calcification in atherosclerosis 63 ; however, this is likely due to microfibrillar components that are known to bind calcium. 64 There is evidence that partially purified elastin isolated from vessels can calcify39,65,66; however, this can also be explained by contaminating microfibrillar components.40,64 In contrast, when purified components such as tropoelastin and intact purified elastic fibers are used, they inhibit calcification in smooth muscle cells.63,67 The importance of purity is further confirmed by the recognition that elastin-based peptides inhibit calcification. 68 Therefore, for use as a biomaterial, impure elastin should be avoided due to potential contaminants. Instead, the use of pure, soluble material would be preferred; this could be in the form of tropoelastin, or elastin-based peptides.
Elastin-Based Peptides and Tropoelastin
Elastin-based peptides bound to surfaces have already been shown to impart reduced thrombogenicity (Table 1). In the first such study to look at the thrombogenicity of elastin peptides, polyGVGP was photochemically immobilized onto silicone rubber; this surface showed decreased fibrinogen adsorption, 69 indicating a reduced capacity for fibrin clot formation. Elastin peptides containing exons 20–24 of the human protein were also coated on to catheters and vascular conduits. When incubated with PRP these showed decreased platelet adhesion and aggregation. Reduced platelet activity was further confirmed by decreased sP-selectin and microparticle release, which are markers of platelet activation. In vivo, these elastin peptide-coated catheters remained patent for a longer time period than uncoated controls and showed decreased fibrin accumulation and thrombosis. 70 Further research revealed that increasing the molecular weight of these elastin-like peptides reduced fibrinogen adsorption and platelet adhesion. 71 Of clinical relevance, expanded polytetrafluoroethylene grafts coated with recombinant elastin-mimetic triblock polymers were assessed in a baboon extracorporeal femoral arteriovenous shunt model, in which a section of an exposed artery was replaced with the graft. These grafts demonstrated increased patency, accompanied by decreased platelet adhesion and lower fibrin deposition. 72
Similarly, we have recently shown that recombinant human tropoelastin 41 decreased the time to thrombus formation upon exposure to static and flowing whole blood when bound to metallic 73 and polymeric substrates (Bax et al. in preparation) (Table 1). For optimal use of elastin and its derivatives in biomaterials, pure tropoelastin and elastin-based peptides should be used to specifically exploit their biological and mechanical features. Further, determining the mechanism responsible for elastin's hemocompatibility would allow these goals to be accomplished, while avoiding the risk of calcification.
Conclusion
There is mounting evidence for the hemocompatibility of elastin and its derivatives. This is demonstrated by examination of its limited interactions with platelets and by a reduction in thrombus formation both in vitro and in vivo. Elastin displays lowered thrombogenicity in various forms, including elastic fibers, purified elastin, tropoelastin, and elastin-based peptides. Additionally, it is a promising candidate for use as a vascular biomaterial or in tissue-engineered constructs due to persistent mechanical properties, favorable association with endothelial cells, and inhibition of smooth muscle cell proliferation. The next generation of elastin-based constructs for use as biomaterials should encompass knowledge of the functional regions of tropoelastin, including delineated nonthrombogenic and cell interactive regions of the molecule, to generate constructs that elicit the desired biological responses.
Footnotes
Disclosure Statement
No competing financial interests exist.