
Abstract
Bone has the capacity to repair itself after an injury, and this occurs in normal fracture repair. This reparative process can be harnessed to regenerate segments of bone using distraction osteogenesis, in which the healing bone is slowly stretched. The use of animal models is identifying the important sources of cells for this endogenous bone regeneration, signaling molecules that regulate this reparative process, and the environmental cues important for success bone regeneration. A more complete understanding of the cells and pathways involved in this process can be applied to improve the outcome of distraction osteogenesis and to the development of methods to enhance endogenous bone regeneration.
Introduction
Fracture Repair
Fracture healing is a complex regenerative process initiated in response to injury, in which bone can heal by primary or secondary mechanisms. In primary healing, new bone is laid down without any intermediate. This type of healing is rare in a complete bone fracture, except when the fracture is rigidly fixed through certain types of surgery. In the more common secondary healing, immature and disorganized bone forms between the fragments, which is termed the callus.2–5 During the fracture repair process, cells progress through stages of differentiation reminiscent of those that cells progress through during normal fetal bone development. In normal development of long bone, undifferentiated mesenchymal cells initially form a template of the bone, which differentiate to chondrocytes. This cartilagenous template is termed the bone's anlage. After this phase, blood vessels enter the cartilaginous template, and osteoblasts, which differentiate from perivascular and other cells surrounding the bone form bone.
There are, however, several important differences between bone repair, and development. One is that repair does not need to progresses through a cartilaginous template. Another is that the liberation of growth factors in the extracellular environment and inflammatory mediators initiates fracture repair, and the activation of these factors does not occur during development. Indeed, this inflammatory initiation of repair processes may be the fundamental difference between development and regeneration. This is one reason that agents which modulate inflammation can effect bone formation, and although pathways such as the prostaglandin pathway can have both positive and negative effects on bone repair, an inhibition of prostaglandin activity, especially cyclo-oxygenase two, will inhibit bone formation, and indeed this has been used clinically to prevent bone formation.6–8
Osseous repair progresses through closely integrated phases. In the initial phase of fracture repair, bleeding from the damaged tissues causes a hematoma at the fracture site, stopping blood loss and liberating growth factors and cytokines. Endothelial cells respond by increasing their vascular permeability, allowing leukocytes, monocytes, macrophages, and multipotential mesenchymal cells to reach the fracture site. 9 The blood supply is temporarily disrupted for a few millimeters on either side of the fracture site, producing local necrosis and hypoxia. It is likely that necrosis also results in the release of sequestered growth factors (e.g., bone morphogenetic proteins [BMPs]), which promotes differentiation of the surrounding mesenchymal cells into bone-forming cells.10–14 In the proliferative phase, undifferentiated mesenchymal cells aggregate at the site of injury, proliferate, and differentiate, presumably in response to growth factors produced by the injured tissues. 5 This process involves both intramembranous and endochondral ossification. Intramembranous ossification involves the formation of bone directly from committed osteoprogenitor cells and undifferentiated mesenchymal cells that reside in the periosteum, resulting in hard callus formation. 15 During endochondral ossification, mesenchymal cells differentiate into chondrocytes, producing cartilaginous matrix, which then undergoes calcification and eventually is replaced by bone. The formation of primary bone is followed by extensive remodeling until the damaged skeletal element regains its original shape and size.2,4,15
When fracture healing is impaired, osteoblastic differentiation is inhibited, and undifferentiated mesenchymal tissue remains at the fracture site. In patients, this outcome ultimately results in nonunion, requiring additional surgery for successful healing. A variety of mechanical or biological factors, including the amount of motion at the fracture, systemic diseases such as renal failure, and degree of vascularity, can contribute to poor bone healing.16,17 Since up to 5% of fractures, depending on the patient's age and the location of the injury, go on to nonunion, the development of novel therapies to reduce the rate of nonunion and to better treat nonunions is an area of intense investigation. Studies have examined signaling pathways normally activated during fracture repair. In the case of one of these signaling pathways, BMP signaling, such studies have already led to improved clinical management. Certain BMPs are liberated early in the repair process, and they stimulate undifferented mesenchymal cells to achieve an osteoblastic phenotype. Tibial fractures are a high risk injury for developing a nonunion and clinical studies show that treatment with select BMPs will improve the rate of healing in this situation. 18
Another pathway that plays an important role in bone repair is β-catenin. There is upregulation of β-catenin during the healing process in human and murine fractures. β-catenin binds to Tcf transcription factors to activate gene expression. Tcf-dependent transcription is activated in both bone and cartilage formation during fracture repair. Several Wnt ligands, which activate β-catenin signaling, are expressed during fracture repair. Treatment with Dickkopf-1, which inhibits Wnt ligand activation of canonical β-catenin signaling, 19 completely inhibits β-catenin signaling and the normal healing process, showing that Wnt ligands regulate β-catenin in fracture repair.20,21 Healing was repressed in mice conditionally expressing either the null or stabilized β-catenin alleles. Fracture repair was also inhibited in mice expressing osteoblast-specific β-catenin null alleles (driven by a fragment of the type one collagen promoter). In stark contrast, there was dramatically enhanced bone healing in mice expressing an activated form of β-catenin, whose expression was restricted to osteoblasts. These results demonstrate that β-catenin functions differently at different stages of fracture repair. In early stages, precise regulation of β-catenin is required for pluripotent mesenchymal cells to differentiate to either osteoblasts or chondrocytes. Once these cells have become committed to the osteoblast lineage, β-catenin positively regulates osteogenesis. 20 Thus, simply activating a pathway important in bone formation will not necessarily enhance fracture repair. The timing of the use of such agents needs to be precisely regulated in terms of the cells that are exposed and the timing of the exposure to achieve the desired result. Intriguingly, this pathway can be activated using lithium, a drug that is already approved for use in patients.20,22,23
The various signaling pathways that play a role in bone repair also interact with each other during the repair process. For instance, the inflammatory process activates prostaglandin synthesis, which regulates BMP expression in mesenhcymal progenitors. 24 In a similar manner, prostaglandin activity also regulates β-catenin activity. 25 Further, BMP stimulation requires β-catenin to produce bone. 21 Thus, the various signaling pathways involved in bone repair and regeneration do not act alone but in a coordinated manner to allow for bone regeneration.
Distraction Osteogenesis
Distraction osteogenesis stretches the healing callus to gradually regenerate new bone. In this technique a low-energy corticotomy (or cut) in a bone is gradually distracted. This controlled distraction generates new bone within a distracted gap. Typically, an external fixator is used to distract the bone. This process consists of three overlapping phases. It starts with a latency phase, which lasts 5 to 7 days. During this phase, the process of callus formation is initiated. This is followed by a lengthening or distraction phase that is continued until the desired amount of distraction is obtained. Both the rate and rhythm of distraction are vital to the quality of regenerate bone formation. The optimal rate of distraction is 1.0 mm/day, and for practical purposes, this is divided into four daily increments. The rate of distraction is limited not just by the response of the bone, but also due to the ability of soft tissues, such as the nerves, muscles, and vessels to resist damage. While the soft tissues grow to accommodate the new length of the bone, they do not regenerate in a process similar to bone. For instance, although the nerve axon grows in length, the body does not actually produce new neurons. The last part of the lengthening process is a consolidation phase. During this period, the newly created bone becomes strong enough to withstand the stresses of mechanical load. The consolidation phase can last a long time, up to a month for every centimeter lengthened1,26–28 (Fig. 1).
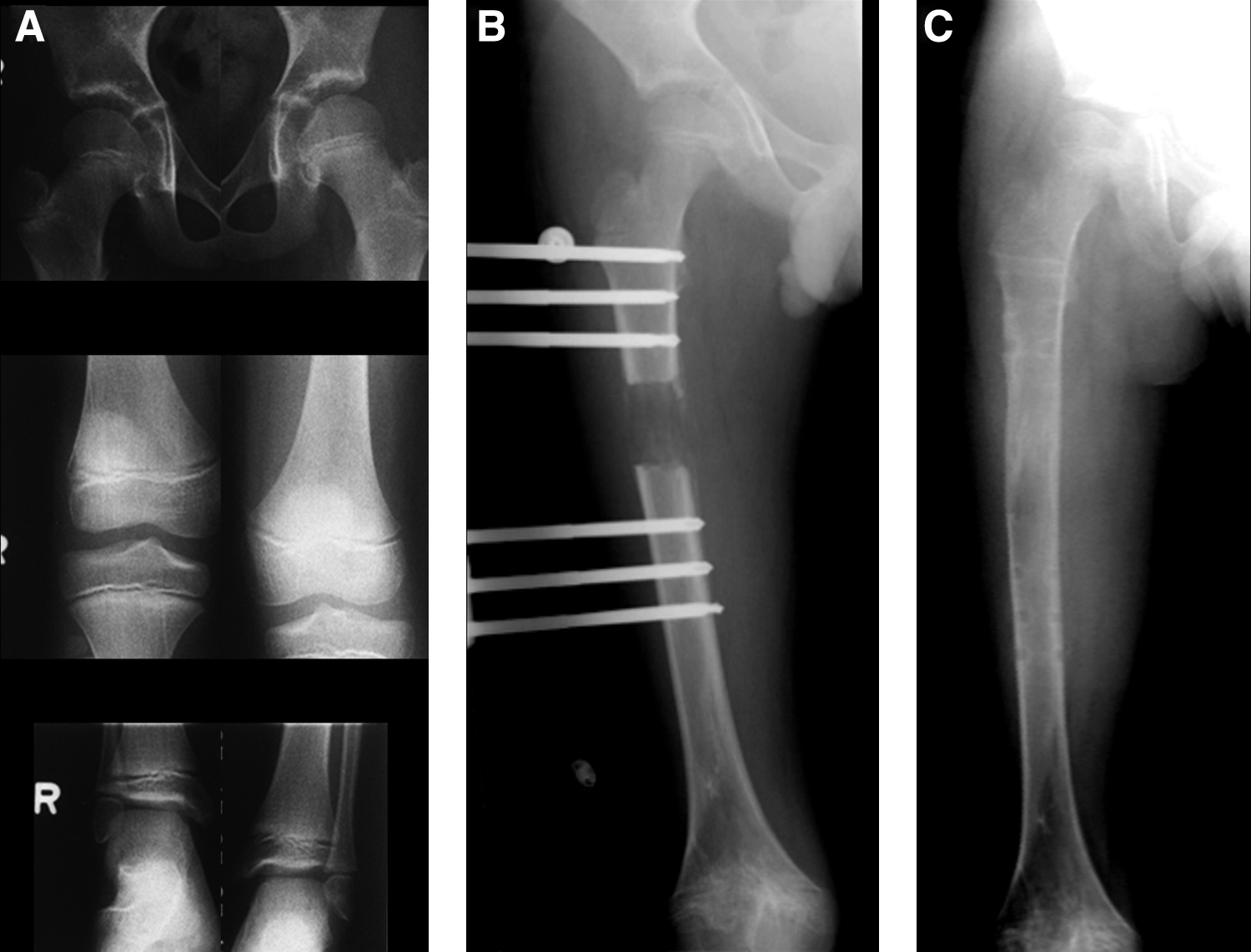
Distraction osteogenesis in a child with a short limb to regenerate bone.
Over the past decade, several studies have investigated the cellular and molecular mechanisms of distraction osteogenesis. During the latency phase, the appearances resemble those of a fracture in its initial stage, with hematoma formation and the accumulation of an inflammatory cell infiltrate. During the distraction phase there is a central fibrous interpose in the middle of the osteotomy site. Vascular sinusoids develop and, along with collagen fibers, become orientated parallel to the force of distraction. It is during the distraction phase that differences from fracture repair can be identified. Cartilage islands rarely form during this phase of distraction osteogenesis, which are more characteristic in fracture healing. After a week of distraction, alkaline phosphatase-positive osteoblast precursor cells appear on the periosteal and endosteal surfaces. As distraction continues, osteoid is laid down, which gradually begins to mineralize. Interestingly, this phase is also distinguished by vascular remodeling, hinting at the importance of the vasculature in this process. During consolidation there is further widespread mineralization, as indicated by alkaline phosphatase activity, and cell differentiation, as demonstrated by the expression of markers of osteoblasts. Upon removal of the external fixation device, a further period of rapid remodeling occurs in response to resuming physiologic load bearing on the limb, where the regenerated bone develops a cortex and medullary canal.29–31
Origin of Osteoblasts
An understanding of the type and source of the cells, as well as the signaling cues important in regulating these cells' behavior, is critical to understanding the process of endogenous bone repair, and the development of methods to improve this regenerative process. Osteoblasts are obviously a critical cell required to make bone. Unfortunately, a lack of unique cell surface markers defining mesenchymal progenitor cells has made the identification of precursor cells problematic. Confounding this issue is the finding that a number of mesenchymal cells can become osteoblasts. Indeed, it is possible that many fibroblastic like cells can behave like osteoblasts when in the appropriate environmental niche. Data showing that bone can form in muscle when the environmental cues are right, as in heterotopic ossification, underscore the ability that different cell types have to become osteoblasts. Cell lineage-tracing studies also suggest that a variety of mesenchymal cells can ultimately contribute to osteoblasts. Muscle cells, skin-derived precursor cells, circulating mesenchymal cells, periosteal cells, and vascular cells all can differentiate to osteoblasts.32–36
The Environment
The environment in which these multipotent mesenchymal cells exist regulates how they differentiate. This environment includes the surrounding tissue type, the extracellular matrix, circulating growth factors, oxygen tension, and the mechanical forces to which cells are exposed. One tissue type that plays a particularly important role in bone regeneration is the vasculature. These vessels potentially provide a source of cells to become osteoblasts. Pericytes are one cell type related to blood vessels, which can become osteoblasts. They surround the external surface of the vasculature, and provide mechanical support to the vessels. This is perhaps the reason that vascular channels are so prominent in distraction osteogenesis. 37 Vascular endothelial growth factor (VEGF) activation, which stimulates vasculogenesis, is required for successful distraction osteogenesis, 38 and adding VEGF to distraction osteogenesis can enhance the repair process. 39
Locally active growth factors can alter how multipotent cells differentiate. Some factors can alter the balance in differentiation from one cell type to another. Some of these factors are active both during repair and during fetal bone development. For instance, during the development of long bones, cartilage is replaced by osteoblasts, and this process can also occur during fracture repair. During these processes, tilting the balance from cartilage to bone formation can enhance bone repair, and several growth factors can achieve this goal. Indeed, the use of BMP2 can enhance the bone formation at the expense of cartilage formation,40–42 a process that is especially important in the latency phase in distraction osteogenesis, when this agent seems to help bone mature faster.
The knowledge that distracting a callus at the appropriate rate causes bone formation shows how the mechanical environment alters the differentiation of mesenchymal cells. 43 Thus, the mechanical stresses on cells at the osteotomy site will alter how they differentiate and can enhance osteogenesis. Compression or very gradual distraction will tilt differentiation toward osteoblasts, whereas higher levels of tension, or shear forces will inhibit bone formation. Part of the control of mechanical environment on bone formation may be through mechanosensors in osteoblasts. 44 The mechanical environment helps regulate the differentiation to fibroblasts, osteoblasts, and chondrocytes.
Despite the current use of distraction osteogenesis to regenerate bone, there are several areas in which research is needed to improve patient outcome. Distraction osteogenesis can take a great deal of time, and a better understanding of the cell pathways that can be used to speed the process is needed. While Wnt signaling, VEGF signaling, and BMP play a role, the study of these pathways is only scratching the surface. Manipulation of the remodeling process can be achieved not only by enhancing the anabolic process of laying down new bone, but also by attempting to slow the process of bone resorption. The use of agents to inhibit osteoclasts has been used in this regard in animals and patients with some success, and much needs to be learned about the particular situations in which osteoclast inhibition will help speed the distraction osteogenesis process. There are various cells that interact in bone repair and remodeling. For instance, osteoblasts and osteoclast interact to regulate each other's activity, and blood vessels are required for osteoblast activity. As such, agents that modulate one cell type often have effects on other as well.32–36
Opportunities for Research in Endogenous Regeneration
There are ongoing challenges in research in this field to identify the cells that will most effectively become new osteoblasts. A lack of unique markers for mesenchymal progenitors is hampering this effort. The better identification of such markers will aid in this research area by better allowing investigators to discover which cells may readily become osteoblasts. The use of genetically modified mice is providing new tools to definitely study the role of various signaling pathways and cell types in this process. Only a more definitive analysis of how the various pathways regulate these processes will we be able to develop pharmacologic or cell therapy approaches to improve the outcome in endogenous regeneration. Data from differences in clinical outcomes in various genetic disorders shed light into pathways important in this process, and a more definitive study of these conditions will identify key factors in regulating bone repair. For instance, rapid consolidation is a common occurrence in distraction osteogenesis in achondroplasia, and understanding the role of fibroblast growth factor signaling may identify new methods to improve bone regeneration. Understanding the similarities and differences between repair and development also likely provides key insights into understanding bone healing, such as the role of inflammatory processes. Finally, a comparative developmental biology study of organisms with enhanced regenerative capacity will likely yield important insights into the regulation of more efficient and effective repair and regeneration of bone.
Clinical Use of Endogenous Regeneration
Despite many unknowns, endogenous repair, or distraction osteogenesis, is currently being used clinically to treat a large variety of bone problems. This includes treating nonunions, by compressing normal bone around the pseudarthrosis; congenital or traumatic limb length inequality, where short bone can be lengthened; problems in which there is a congenital bone deficiency, such as congenital pseudarthrosis of the tibia; or traumatic or pathologic loss of bone, during which new bone is regenerated using a technique known as bone transport, whereby a segment of bone is gradually transported through the limb forming regenerate bone in its wake, until it docks distally, thereby reconstructing the bony defect.
Bone clearly has the ability to regenerate itself. Through the use of distraction osteogenesis, this capacity can be harnessed to regenerate large segments of bone. Understanding the regulatory cellular processes and signaling pathways is crucial to enhancing the efficiency of distraction osteogenesis, which will ultimately improve the outcome for patients with bone defects.
Footnotes
Disclosure Statement
No competing financial interests exist.