
Abstract
Bone regeneration has long been a major focus for tissue engineers and the importance of vascularization to the bone regeneration process has been well documented. Over the past decade, technological advances in the areas of stem cell biology, scaffold fabrication, and protein engineering have significantly enhanced our understanding of the interplay between vascularization and bone growth. This review, therefore, describes the commonly used models for investigating the complex interactions between osteoblastic cells and endothelial cells, evaluates the different tools utilized to investigate the relationship between vascularization and bone growth in vivo, and finally, summarizes possible areas of research related to therapeutic development.
Introduction
Numerous studies have shown that angiogenesis is essential to successful bone repair and regeneration. As resorption of the necrotic bone that forms during the initial step of the fracture repair process requires vascularization at the injury site, inadequate blood supply is also implicated as a major contributor to fracture nonunions.2,3 Nonvitalized bone grafts have relatively low angiogenic potential and more long-term difficulties such as disintegration and limited resorbable graft size.4,5 Open fractures in long bones tend to be more susceptible to nonunions, because injury to the surrounding soft tissues deprives the callus of normal blood supply and also results in bone necrosis. 6 In a similar way, postoperative irradiation and associated blood vessel damage is also associated with poor bone graft incorporation and fracture healing. 7
Controlled release of a few known diffusible signals such as vascular endothelial growth factor-A (VEGF-A) to supplement blood around the fracture site after soft tissue damage has been shown to increase healing in a variety of experimental models. 8 This suggests that increasing the blood vessel supply has a direct impact on bone repair and growth, and that future treatment options need to emphasize vascularization of the bone defect regions. However, researchers are increasingly aware of the vast array of both divergent and cooperative molecular signals governing microvascular growth and remodeling, the numerous soluble effectors and their receptors, matrix degrading enzymes, and other mediators of cell–cell and cell–matrix interactions. Impaired or abnormal function of the vasculature is associated with a wide range of disease pathologies, and thus, proper temporal and spatial and regulation of these angiogenic signals is critical to the formation of functional vascular networks, spanning the various arterial, venous, capillary, and collateral vessel systems. Control of microvascular network growth will be critical to successful transfer of new investigational drugs and devices for bone regeneration into clinical application.
The tools and methodologies to evaluate the relationship of vascularization and bone regeneration include in vitro, in vivo, and computational models. This review is focused on the commonly employed in vitro and in vivo model systems used to investigate the complex interplay between microvascular network assembly and new bone formation. We focus specifically on the interactions between osteoblastic cells and endothelial cells (ECs) in well-controlled two-dimensional (2D) and three-dimensional (3D) coculture models. We also review the computational models that help in understanding these interactions and discuss the in vivo preclinical models that provide the basis to move basic science understanding into new clinical reality. Additionally, because the choice of scaffolds and biomolecules used in different therapies must address the need for vascularized bone growth, we describe the effects that common bone implant materials and signals may have on in vivo vascularization.
In Vitro Assessment of Endothelial Osteogenic Cell Interactions
In vitro assays
In vitro assays provide high-throughput capability for studying complex biological processes. They ensure that the experiments can be conducted in a controlled manner, as the multitude of growth factors, cytokines, and other biomolecules that are encountered in vivo are not present. The relationship between osteoblasts and ECs has been studied in various assays that have been successful in elucidating insights into the mechanisms of their interaction by simplifying the complexity of that process.
ECs are osteoinductive as they drive mesenchymal stem cells toward the osteoblastic phenotype. Cocultures of ECs with osteoblast progenitor cells, 9 bone marrow stromal cells (BMSCs), 10 or osteoblast-like cells 11 in different scaffolds such as alginate, 9 poly(lactic-co-glycolic acid) [PLGA], 11 extracellular matrix, 10 or conditioned media10,11 have shown pronounced mineralized matrix production,9,11 enhanced microvascular network formation,10,11 and increased bone regeneration. 10 The interaction between these cells tends to depend on the differentiation state of both cell types and on the type of ECs used in the study. Coculture experiments demonstrate that ECs stimulate the differentiation of osteoprogenitor cells 12 without affecting the extracellular matrix-producing capacity of the osteoblasts. 13 However, Fuchs et al. 14 have shown that ECs prevent the differentiation of BMSCs into mature osteoblasts by interfering with the expression of osterix, a transcription factor that induces the bone phenotype. These discrepancies can be explained by the fact that ECs have varying effects on different stages of osteogenesis, as coculturing them with osteoprogenitor cells results in the upregulation of ALP gene expression and the downregulation of osteocalcin, a late marker of osteoblastic differentiation. 13 The maturation state of ECs used in the assays is an equally important factor. Rouwkema et al. 15 showed that human umbilical venous endothelial cells (HUVECs) and human dermal microvascular endothelial cells induced the prevascularization of bone-engineered tissue, whereas endothelial progenitor cells (EPCs) did not initiate as many prevascularized structures. The specific cell types and maturation stages ideal for osteogenesis are to be yet determined, and future in vitro assays should be designed to answer these questions.
Additionally, the development of microfluidics has led to a better understanding of the microenvironment that the cells experience and their response to it. Many microfabricated devices have been developed to induce and monitor cell responses to either biochemical gradients16,17 or biomechanical forces such as adhesion, 18 flow, 19 or shear. 20 Zhang et al. 21 used inducible gene expression with laminar flow technologies in a microfluidic system to spatially regulate the expression and secretion of bone morphogenetic protein (BMP)-2 and to evaluate engineered tissue interfaces. Kou et al. studied the effect of different shear flows on the cystolic calcium concentration dynamics of osteoblasts. 20 As shear is known to affect bone growth, such controllable assays can be used to relate the two processes. Microfluidic devices could be used to examine the interaction between ECs and osteoprogenitor cells in a more controlled manner as they can be stimulated at time-varying schedules, and the effect of parameters such as cell seeding density and culture medium composition can be categorically evaluated. These devices are a good platform for evaluating the loading efficiencies and release kinetics of biomolecules in scaffolds used for in vivo studies. They have been used to determine the effect of varying concentrations and combinations of different proangiogenic drugs on the sprouting profiles of capillaries and on EC morphogenesis. 22 Such devices can prove to be useful in investigating the relationship between ECs and osteoblasts and understanding the implication of those interactions on bone growth.
Computational tools
In vitro assays are especially useful when obtaining parameters for developing computational models. Computational modeling provides insights into the mechanisms underlying the outcomes measured in experiments and is a useful tool for understanding the temporal action of the growth factors and biomolecules used for vascularized bone growth and measuring the relative importance of the different components in the process. They have been extensively explored in the assessment of angiogenesis and vascularization of 3D tissue constructs. Various models spanning different scales and focusing on different aspects of angiogenesis, an integral part of tissue regeneration, have been developed. These can be classified as continuum models,23–27 discrete models,23,24,28–30 stochastic models,31,32 and cellular Potts models.33,34 In a different approach, the Popel group has developed a multiscale integrative model with specific modules for various growth factor receptor pairs and ECM proteolysis. 35 The development of such multiscale computational models has helped the field of angiogenesis resolve complex interactions and equipped researchers with drug discovery tools. The use of computational models in the bone microenvironment may prove to be similarly effective.
The use of computational modeling in bone healing, though not as extensive, has provided some remarkable observations. Geris et al. 36 used an integrative approach to model nonunions as they occur in a clinical setting and showed that successful fracture healing occurs because of the simultaneous presence of vasculature, growth factors, and precursor cells in the callus. Their mathematical model can be used to test treatment strategies and optimize experimental design. Several models have evaluated bone growth inside a 2D or 3D scaffold.37–39 Byrne et al. examined the impact of scaffold properties such as porosity, Young's modulus, and dissolution rate on tissue differentiation by combining random walk algorithm, mechanoregulation of stem cell fate, finite element models, and rule of mixtures to determine material properties. 40 A recent study used finite element analysis to evaluate the effect of cell seeding density of MSCs and mechanical loading on vascularization and new tissue formation inside a scaffold. 41 The authors partitioned the scaffold into a grid to identify spaces for cellular activities such as migration, proliferation, differentiation, apoptosis, and synthesis of new materials. They concluded that the initial seeding patterns affected the dynamics of angiogenesis and tissue differentiation, and it would be beneficial to identify the optimal loading patterns for site-specific loading conditions. Thus, computational models can be useful in gaining insights into mechanisms that are otherwise difficult to measure experimentally as they allow for low-cost parametric analysis of different interactions. Such computational evaluations could be extended to critical-sized defects, fractures, and other models useful for translating biologics into clinical applications.
In Vivo Assessment of Bone Regeneration
In development, vascularization precedes osteogenesis 42 during both intramembranous and endochondral ossification, and it is suggested that microvessels accelerate bone formation even before flow has been established. 43 Blood vessels have been implicated in the transport of both osteoclast precursors 44 and osteoprogenitor cells 45 to the defect site. It has also been shown that an existing vascular network might serve as a scaffold for subsequent osteoblast differentiation.46,47 The design and implementation of appropriate in vivo models can demonstrate the effectiveness of the different therapies on vascularized bone growth over a long period of time. Adult animal models can be used to study how the induction of angiogenesis impacts the process of host bone growth in two ways. Several enabling imaging technologies used with bone defect models permit evaluation of blood vessel formation and new bone growth simultaneously within the same experiment. These are useful to discern vessel-bone integration and the interdependence of vessel and bone growth. Alternatively, experiments may be designed in parallel to rigorously quantify the structural remodeling response to proangiogenic stimulation in nonosseous tissues that are accessible by optical microscopy and allow for temporal imaging of vasculature. These results can be paired with companion experiments involving more traditional models of bone tissue growth. Such sequential experiments are useful for studying progressive vessel development and maturation in response to a drug, measuring individual cell homing to vessels, and exploiting transgenic animals for identifying mechanisms of drug action.
Models for simultaneous evaluation of vascularization and bone growth
Cranial and segmental defects are the common animal models used for studying simultaneous vessel and bone growth for nonload-bearing and load-bearing bones, respectively. These have been implemented in mice, rats, rabbits, sheep, canines, and other large animals. Other animal models such as rat vertebral defects 48 and rat mandibular defects have also been used to demonstrate the effects of growth factors and scaffolds on vascularized bone growth. Many studies have used critical-sized defect models to assess the effect of an angiogenic factor on bone healing.49–51 Wang et al. showed higher bone growth when a vascular bundle was implanted along with autologous bone marrow mesenchymal stem cells plus beta-tricalcium phosphate in a 15 mm segmental defect in a rabbit femur. 52 This suggests that cells from the vasculature either aid in the transport of factors that aid in bone growth or participate in the process directly. Fracture repair models have also been extensively used to measure the efficacy of different therapies. In addition to studying the effectiveness of different growth factors and scaffolds, several reproducible fracture models that explicitly examine the effect of vasculature have been developed. An ischemic mouse model in which femoral ligation affects limb vascular perfusion results in a smaller, less vascularized callus, leading to the formation of more fibrous and adipose tissue. 53 A nonunion mouse femur model, where the devascularization of the lateral femoral periosteum results in no bony callus formation even 35 days after injury, provides pivotal evidence for the importance of vascularization during bone healing. 54 The use of such animal models to evaluate the importance of vascularization is only as good as the evaluation tools available to assess them. Hence, the development and validation of techniques that enable measurements of vessel and bone growth in such animal models is imperative for validating translational therapies.
Over the last three decades, since its introduction in the 1970s, contrast-enhanced micro-CT has been extensively used to measure and characterize bone growth in defects created in animal models. The development of high-resolution scanning and the availability of dedicated scanners for noninvasive small animal imaging has made micro-CT a very useful tool in bone growth research. Schambach et al. 55 discuss the applications, advances, and limitations of micro-CT in their review, the details of which are beyond the scope of the present article. One of the main limitations is that micro-CT does not allow for the measurement of vascularization and blood flow in live animals. Microfil-enhanced micro-CT has been used to measure the density of blood vessels in bone defects filled with implants at the end of the study. However, this is not possible in the case of live animals, where other methods have to be adopted. Kaigler et al. 10 used laser Doppler perfusion to assess blood flow and enhancement of local angio/osteogenesis in nonirradiated and irradiated rat calvarial defects after implantation of a PLGA scaffold and/or a VEGF scaffold.
Imaging tools have been employed to access bone healing in many different assays. As cell recruitment to the implant site is crucial to healing, imaging that allows cellular visualization is useful to discern the mechanism of both new bone growth and remodeling of existing bone. Recently, Ishii et al. 56 used two-photon microscopy to show that sphingosine 1-phosphate (S1P) mobilizes osteoclast precursors and regulates bone homeostasis. Zilberman et al. 57 did real-time imaging to measure nonunion fracture repair by genetically engineered mesenchymal stem cells following systemic administration of the fluorescent bisphosphonate imaging agent OsteoSense, an hydroxyapatite-directed bone-imaging probe, and imaging by fluorescence molecular tomography (FMT). The use of fluorochromes that emit in the near-infrared spectral window makes the deep tissue penetration of photons possible, because hemoglobin and water absorb minimally at that range. Unlike reflectance imaging, FMT can penetrate a few centimeters into the tissue. Although there are not many dyes or markers that have excitation spectra in that region, mice models with infrared reporter genes are currently being developed. FMT can be combined with high-quality reflectance imaging, overlaying the tomographic images with anatomical features and thus obtaining highly sensitive molecular information in 3D. Bioluminescence imaging (BLI), another in vivo imaging tool, is based on the detection of light emitted by living cells that express the luciferase gene, thus allowing for noninvasive imaging of whole animals. As BLI is based on luciferase expression, which can be put under the control of tissue-specific elements, it is very useful in monitoring specific processes such as vasculogenesis. Although BLI has been used to study and monitor the effectiveness, survival, and localization of luciferase-labeled stem cells in healing calvarial defects,58,59 it could be potentially used in VEGF receptor luciferase mice to monitor vessel ingrowth in bone grafts and fracture sites. One of the limitations of BLI is the inability to resolve depth because of tissue scattering and adsorption, which limits spatial resolution. Combining intravital imaging of single-cell homing through newly formed vascular conduits to defect sites with bone growth measurements could prove to be a useful tool for assessing impact of therapies on different cell phenotypes.
Other noninvasive imaging modalities may lend themselves to experimental and computational integration and provide parametric information such as rate of vessel or bone growth that are important for measuring the efficacy of different treatments. Magnetic resonance imaging (MRI) enables the identification of pretreated transplanted cells migrating to the defect site over multiple time points unlike more traditional end-point methods such as flow cytometry. Computational models can also be used to gain additional insights into in vivo models. Swider et al. 60 did parametric sensitivity analysis to estimate the effect of cell adhesion and growth factor gradients on heterogeneous bone growth in noncritical defects in a canine model. The development of new enabling technologies such as multiscale computer models, MRI for tracking recruitment of circulating vascular tissue progenitor cells, near-infrared reporter mouse models for in vivo imaging, and intravital bone growth imaging will allow for the integrated quantitative assessment of bone formation and vascularization. This integration will be important for identifying and evaluating clinically relevant therapies.
Models for evaluating vascularization prior to bone growth
As the rate and extent of vascularization directly relates to bone growth, in vivo assays that assess angiogenic capabilities of contending factors prior to their impact on bone growth are equally important. The models that have been used are transparent chambers (rabbit ear, hamster cheek, dorsal skin), corneal micropockets (rabbit, rodent), vertebrate embryos (knockouts), and ischemic hind limbs (rabbit, rodent). Microvascular remodeling is an important indicator for the development of new therapies that harness the host's own responses to injury. The transparent chamber and corneal pocket assays enable the evaluation of changes in vascularization and angiogenesis as a consequence of local or systemic delivery of small molecules or growth factors. Additionally, they provide an easily accessible platform for imaging, as they allow us to monitor and quantify the temporal and spatial remodeling in the local microvascular network. Unlike critical-sized defect models discussed earlier where vascular ingrowth is a terminal measure, the dorsal window chamber or backpack model holds the constructs with drugs in place without sutures, allowing for temporal assessment of the drug effects by intravital microscopy.61,62 Chen et al. have created a subcutaneous implant of prevascularized fibrinogen gel surrounded by a PDMS well. 63 Histological analyses of explanted tissue allows quantification of functionality of the engineered vessels by identifying those lumens through which red blood cell flow occurs and by determining the area fraction of the tissue consisting of functional vessel lumens. In this way, tissue constructs can be optimized for vascular integration in vivo prior to implantation in a defect model. For example, one study showed that the increased capability of engineered vessels comprised of EPC-derived ECs (EPC-ECs) versus HUVECs to form anastomoses upon implantation, 64 allowing future research to focus on the more promising EPC-ECs.
Repeated measures of vascularization are important to determine the rate of vessel growth as a function of different treatments. Additionally, the temporal evolution of vasculature in response to different drugs is an important indicator of the long-term impact of the treatment. The backpack and corneal pocket models described above allow for such measurements. These models are also useful for studying vascularization in transgenic animals, thus providing information about the receptors and ligands involved in the process. For example, Nickerson et al. used CCR2−/− mice to conclude that CCR2 receptors are needed for arteriole remodeling. 65 Future studies can involve evaluating the effects of different drugs in knockout animal models to elucidate their mechanisms. Although such animal models are extremely valuable and provide insights into the effects of different therapies on vascularization, they are not frequently used in concurrence with bone defect models.
Scaffolds, Biomolecules, and Cells for Vascularized Bone Growth
Scaffold technology
Although autografts have been established to have fewest complications and highest success rates as bone grafts, they are scarce and cannot be used for large surgeries, and they are associated with donor site morbidity. 66 Alternatively, allografts, although easily available, have the risk of disease transmission and immune reactions. 67 This has been the motivation for research in engineered scaffolds and, over the years, their design parameters have been optimized to have the properties of osteoinductivity, osteoconductivity, biocompatibility, variable degradation rate, and high mechanical integrity at sites of bony repair. 68 The scaffold architecture, which includes porosity, pore size, surface area to volume ratio, and pore interconnectivity, has been shown to affect its functionality. This is because vessel infiltration into scaffolds is dependent on these parameters. However, fabrication of appropriate biomaterials for tissue regeneration requires knowledge of both the biomaterial and the normal repair mechanisms of the defect site. The latter is especially crucial, as there are fundamental differences in bone formation, 42 mechanical requirements,69,70 and local progenitor populations 71 related to the exact site of a bony defect.
Several natural polymers such as collagen, matrigel, and chitosan have been used as delivery vehicles for biomolecules and cells into the defect region. With the advent of synthetic polymers, we have been able to control properties of the scaffold architecture such as porosity and biodegradability to a greater extent, thus allowing for the design of more compatible and need-oriented delivery vehicles. Although a detailed discussion of natural and synthetic polymers is beyond the scope of this review, this section will focus on the impact of scaffold design and on vascularization and bone growth.
The selection of a scaffold is important in experimental design, because a good material can stimulate and assist bone growth by itself. Rentsch et al. 72 showed that tissue-engineered polycaprolactone (PCL) scaffold coated with collagen I/chondroitin sulfate shows a relative increase in bone volume compared with the PCL scaffold coated with collagen I/chondroitin sulfate and seeded with hMSCs in long bone critical-sized defects in rats, although the latter group showed an increase in matrix deposition inside the scaffold. Discerning the relative effects of scaffolds and other vasculogenic factors used is also important when designing and evaluating in vivo experiments. A recent study evaluated the efficacy of a polyethylene glycol-based bioartificial hydrogel matrix in inducing vascularization in vivo. They showed that these matrices could sustain VEGF growth for over 2 weeks, giving rise to an increased rate of reperfusion in a mouse model of hind-limb ischemia. 73 These studies reveal the importance of knowing and harnessing the synergies between proangiogenic molecules and the host tissue responses to the scaffolds.
In the absence of a vascularized graft, the transport of nutrients occurs primarily through, and is thus effective for, only 100–200 μm in high metabolic tissues. The transport of perivascular mesenchymal stem cells, which differentiate into osteoblasts, also depends on the establishment of blood vessels. It is important that the neovessels that form in the defect region anastomose with the systemic circulation. Therefore, a graft that induces vascularization by supporting EC migration, adhesion, and proliferation is more conducive to bone defect healing. In some cases, RGD peptides that are attached to the scaffolds can increase EC and osteoblast adhesion 74 and promote regeneration. Techniques such as increasing the pore size of the scaffold, 75 using VEGF-releasing scaffolds in bone defect regions, 76 and in vitro prevascularization of tissue constructs in cocultures of endothelial and bone cells 12 have been adopted to increase the vascularization capabilities of the implanted graft. Increasing the pore size could enhance the connectivity and rate of formation of blood vessels in the center of the implants, and prevascularization of the bone grafts would negate the time required for the recruitment of ECs.
Prepatterned scaffolds have been used to increase vasculature by “vascular guidance.” Muller et al. 77 used computer-aided design to model the vascular channels within a fronto-parietal defect in a patient. They used vasculature originating from the anterior branch of the middle meningeal artery, incorporated it into PCL scaffolds fabricated using fused deposition modeling, seeded it with bone marrow-derived stem cells, implanted it into a rat model, and observed vessels growing along the prepatterned lines. However, the concern over the host vasculature fusing with the graft vessels remains to be thoroughly investigated. Nevertheless, this method could be potentially adopted for creating vitalized bone grafts that could be used in massive defects as the time taken to vascularize an implant from outside in takes longer than the ischemic tolerance levels of the cells seeded in the center of the grafts. 78 Alternatively, scaffolds may be coated with ECs or “vasculogenic agents,” such as growth factors, and small molecules that induce blood vessel formation resulting in accelerated bone defect vascularization.
Biomolecules
BMP-7 and BMP-2 are two growth factors that have been approved by the FDA for use in humans for long bone repair with bone-derived collagen as the delivery vehicle. In some instances, these are considered equivalents to the gold standard of autogenous iliac crest bone graft for bone regeneration. 79 Recombinant BMP (rhBMP)-7 is marketed as Osteogenic Protein-1 (Stryker, Kalamazoo, MI) and has been evaluated in open tibial shaft fractures, 80 distal tibial fractures, 81 and tibial nonunions, 82 among others. BMP-2 is marketed as Infuse (Medtronic Sofamor Danek, Memphis, TN) and has been shown to be effective in open tibial fractures 83 and segmental bone defects. 84 However, the use of these molecules clinically has not yielded as good results as seen in the preclinical trials. A recent, controversial, randomized trial of open tibial fractures showed no improvement in the rate of fracture healing with the use of BMP when compared with the controls. 85 BMP-2 has also been shown to affect EC tube formation and neovascularization, although it has little effect on EC proliferation. 86 This begs the question of whether or not the exogenous use of a single growth factor does indeed result in biologically relevant fracture healing, given the mechanistic complications of the process. If so, are there other possible candidates?
VEGF signaling, one of the most vital angiogenic pathways, has been shown to promote bone growth.44,87,88 Traditionally thought of as an endothelial target, this protein family has also demonstrated mitogenic effects on both osteoblasts and osteoclasts. 89 Further, osteoblasts express an array of VEGF-specific receptors and are capable of responding to and secreting VEGF proteins. In response to low oxygen availability, osteoblastic populations will increase production of VEGF, mediated via the hypoxia inducible factor(s) [HIF]α pathway. 90 Further, the VEGF pathway acts directly on osteoblast populations to increase production of many osteoinductive factors, including transforming growth factor-β, insulin-like growth factor (IGF)-1, and fibroblast growth factor (FGF)-2. Alternatively, VEGF stimulation of ECs initiates secretion of bone-forming factors.91,92 The dual delivery of angiogenic and osteogenic growth factors increases healing in critical-sized cranial defects, implying a synergistic effect between these factors. 49 Although many preclinical studies used VEGF either alone or in combination with other growth factors, one of the major drawbacks of clinically using VEGF is that its recombinant form is very fragile. The need for optimizing the delivery of this molecule has led to a successful cell-based nonviral gene therapy using VEGF in the segmental defect model. 93
Behr et al. 94 recently showed that fgf-9+/− mice have reduced bone regeneration compared with wild-type mice because of a decrease in vascularization at the defect site. Qu et al. 95 showed that β-FGF-mediated ex vivo gene transfer based on BMSCs seeded on nanohydroxyapatite/polyamide66 composite scaffold can accelerate vascularization and bone regeneration in rat calvarial defects. Several studies have shown that platelet-derived growth factor (PDGF) stimulates both migration96,97 and proliferation 98 of osteoprogenitor cells and osteoblasts, increases VEGF production, 99 enhances angiogenesis, and stimulates fracture healing and bone regeneration in rat tibial fractures 100 and rabbit tibial osteotomies. 101 Recombinant PDGF in a beta-tricalcium phosphate matrix is currently available for periodontal bone defect treatments as GEM 21S (Biomimetic Therapeutics, Franklin, TN), and clinical trials for ankle fusion and fracture healing started in 2007.
S1P is a bioactive phospholipid that has been shown to enhance migration and proliferation in both endothelial and smooth muscle cells in vitro.102–105 S1P has also been implicated as a potent osteoblast mitogen in vitro for stimulating proliferation 106 and exerting antiapoptotic effects in osteoblastic cells. 107 Indicative of its role in promoting arteriogenesis, S1P possesses significant ability to promote vascular stabilization by recruitment of pericytes and mesenchymal stem cells to newly formed vessels 108 and to stimulate SMC proliferation, 103 migration, 108 and differentiation into a more contractile phenotype. 104 Ishii et al. 56 showed that the reciprocal regulation of S1P-dependent chemotaxis via S1P receptor-2 controls bone remodeling by finely regulating osteoclast precursor localization. This could be a therapeutic strategy for osteoporosis and other osteoclast-dependent bone remodeling events. Sustained release of S1P and FTY720, a S1P receptor-1 and receptor-3 agonist from biodegradable polymers, promotes microvascular network growth and arteriolar expansion. 62 Petrie et al. 50 showed that the use of FTY720 promotes local microvasculature formation and cranial defect regeneration in a rat model. Superior osseous integration across the host–graft interface, significant enhancement in smooth muscle cell investment, and reduction in leukocyte recruitment were evident in FTY720-treated groups in a rat tibial defect model as well. 51 FTY720, a small molecule drug, is more stable and has a longer half-life than many growth factors that are candidates for clinical applications.
Some of the major drawbacks of biomolecules are the cost and potential safety issues that arise from using recombinant proteins. To that end, attempts have been made to study the efficacy of inorganic angiogenic factors such as copper ions. Barralet et al. evaluated the efficiency of copper sulfate in a calcium phosphate scaffold on inducing microvessel growth and compared it with VEGF-loaded scaffolds. 109 They showed they could accelerate and direct vessel growth with such inorganic ions. The high cost of biomolecules as therapeutic agents also merits further investigation of small molecules such as S1P receptor drugs, prostaglandin E2 agonists, and HIF1-α stabilizers for use in bone tissue engineering. These alternative options may also make getting FDA approval for such therapies relatively easier.
Cell-based therapy
Over the last few years, scaffolds seeded with stem cells like bone marrow derived mesenchymal stem cells (BMMSCs) and adipose derived stem cells (ADSCs) have been used as bone grafts, and in many cases the cells have made the scaffolds better regenerative agents. Stem cells from different sources have been shown to release a multitude of cytokines such as urokinase plasminogen activator, hepatocyte growth factor (HGF), tumor necrosis factor alpha, and VEGF, which are important for vascularization. 110 The ECs use these angiogenic cues to migrate to the defect site and initiate protease mechanisms that result in extracellular matrix degradation. BMMSCs have also been shown to differentiate into osteoblasts in vitro, and they have been seeded into scaffolds both before and after differentiation, with and without additional growth factors. BMMSCs along with BMP-2 have shown accelerated bone healing in the rat cranial defect model when used in collagen composites and chitosan scaffolds.111–113 Although BMMSCs increase bone formation, their effect is augmented when they are seeded along with EPCs, as shown in a critical-sized femoral defect model. 114 EPCs can affect vascularization by differentiating into ECs and by releasing chemotactic molecules. The combination therapy involving BMMSCs and EPCs results in both a larger callus and increased bone formation in the first 4 weeks, suggesting that the earlier presence of vasculature is important for bone formation. Additionally, although BMMSCs have shown significant promise for regenerative medicine, they have a short lifespan and are not available in large quantities.
ADSCs have been shown to differentiate into osteoblasts and, unlike BMMSCs, are easily available. Cowan et al. 115 showed bone bridging in ADSCs seeded in PLGA scaffold in rat calvarial defects. Similar to BMMSCs, ADSCs are more effectual when implanted after osteoinduction.116,117 ADSCs secrete angiogenic factors such as VEGF and HGF at bioactive levels and could serve to increase vascularization when used at the defect site. 118 Freshly isolated ADSCs also contain a substantial subpopulation of cells that stain positive for EPC and HSC markers, suggesting that many of them can be induced to differentiate into vascular cells. 119
There are several studies that support the use of cells for enhancing the amount or rate of bone healing, but many others show that the addition of cells does not add to the effect of growth factors such as BMP-2. As mentioned earlier, Rentsch et al. 72 obtained better bone volume with just their scaffolds in tibial bone defects compared with animals treated with cell-seeded scaffolds. On the contrary, Stephan et al. observed minimal cranial defect healing with only BMP-2 in chitosan in the absence of MSCs. 113 Recently, Kolambkar et al. 120 showed that the perforations in the nanofiber mesh used as tubular implants in rat tibial defects served to expedite bone remodeling and new bone growth, suggesting that osteoprogenitor cells migrate into the defect region from the surrounding muscle. It is possible that an external cell source is required for defect healing in cases where there is a lack of surrounding soft tissue, as in cranial defects. This hypothesis requires further investigation, and the possibility that growth factor versus growth factor plus cell treatments are a function of the defect site must be examined.
The need for an external source of cells for ectopic bone formation is well documented, 121 but bone formation in a nonbony environment has been observed with BMP-2 without the application of cells. 122 Additionally, it is possible that the occurrence of injury could stimulate stem cell circulation from soft tissues via hypoxia and thereby diminish the need for external cells. Thus, the need for a cell-based therapy can also be translated into the need for recruitment of stem cells to the defect area. It is hypothesized that this recruitment of cells to an injury site is directly related to the number of stem cells circulating in the peripheral blood. The possible sources of stem cells being recruited to the defect region are shown in Figure 1. External stem cells have been shown to migrate to the fracture 123 and critical-sized defect sites. 59 Granulocyte colony stimulating factor (G-CSF), commonly used for mobilizing stem cells from the bone marrow, is known to modulate the cytokines in the inflammatory response as well. Bozlar et al. 124 used a subcutaneous injection of G-CSF in rats with tibial fractures and showed that the radiographic score of the treatment groups was better than that of the control group. However, they could not establish any substantial difference in bone union between the two groups. In addition to mobilization, stem cell homing at the defect area is crucial to healing. Stromal cell-derived factor-1 (SDF-1) is known to mediate the homing of stem cells to injured tissues via the CXCR4 receptor. 125 A recent study showed that the addition of SDF-1 increased the effectiveness of BMP-2 in ectopic bone formation, suggesting that it effectively directs circulating stem cells to the implant site. 126 The homing of circulating stem cells is dependent on the surrounding vasculature, as the cells would be arrested there, causing them to migrate across the endothelium and differentiate into functional tissue. Thus, therapies that increase the vasculature of the injury site and mobilize stem cells into the peripheral blood can be very effective in bone defect healing. This type of therapy combining systemic stem cell mobilization with local tissue regeneration has been extensively exploited for cardiac tissues and remains to be further explored for bone healing.
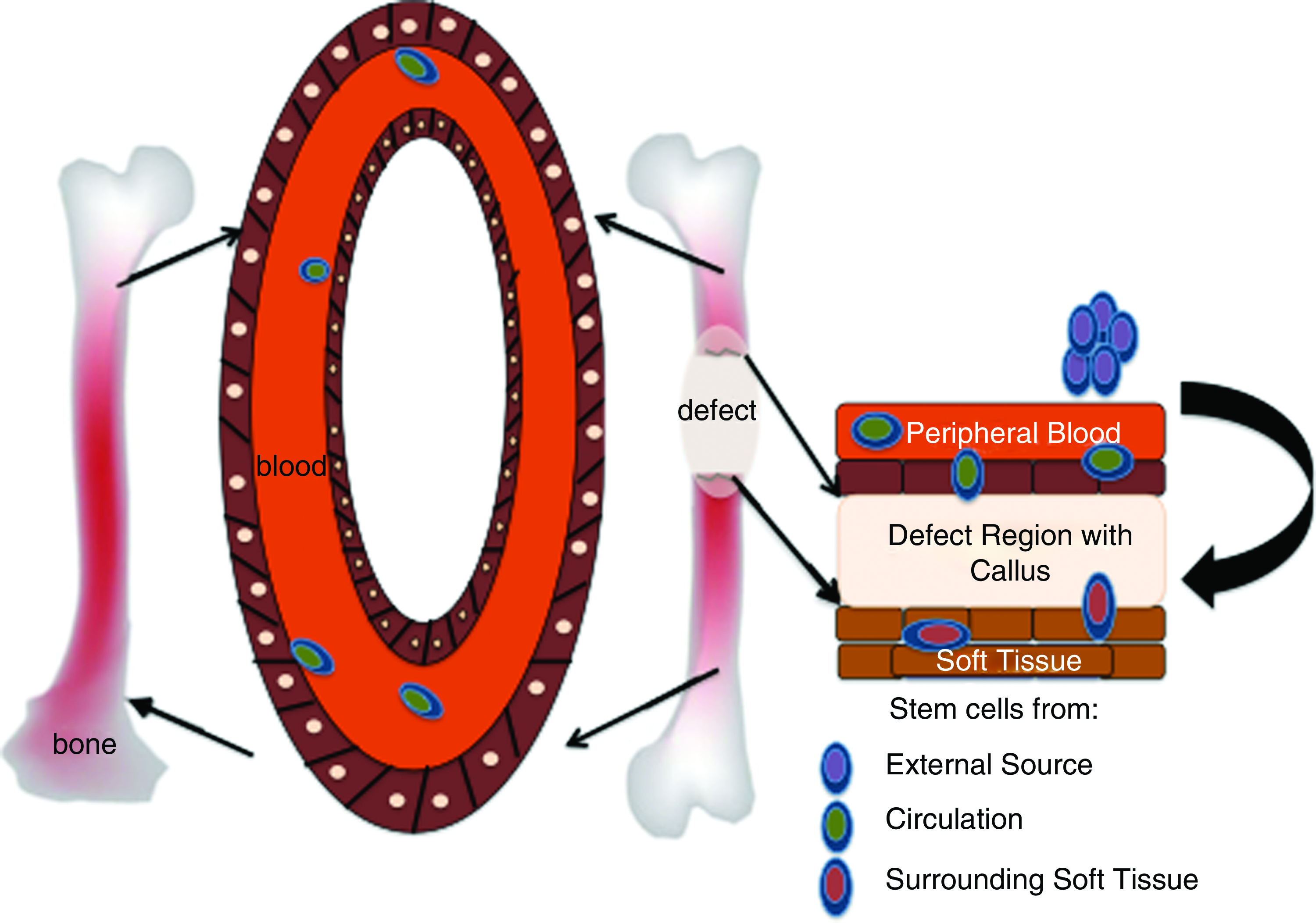
Cell sources for bone regeneration. Color images available online at www.liebertonline.com/teb
Regulatory Approval
The ultimate goal of bone tissue engineering is to develop therapies that will gain regulatory approval and improve patient care. To that end, it is important that all assays and models are standardized. The different variables in in vitro culture systems, such as the contact mechanism between cell types, the passage number of the cells, the relative seeding density of both cell types, and the culture media, need to be optimized and kept consistent to enable comparison of results across studies. It is also crucial to important to remember that FDA approval of investigations supported by animal models require usage of appropriate animal models, identification of indicators for long-term prediction of outcomes, and standardization of surgical techniques, assessment metrics, and evaluation time points.
The choice of scaffolds and biomolecules should also be guided by the time taken to get FDA approval. Their design, architecture, intended use, and mechanism of action are critical in this process. Although synthetic scaffolds enable easier manipulation of specific characteristics, natural polymers likely go through the regulatory process more easily. This does not imply one is better than the other, although it is important to be aware of the possible consequences of using different materials. Combination therapies that involve biomolecules and cells need to go through more rigorous approval processes. The lack of predicate proangiogenic therapies for cardiovascular regeneration might complicate the prospect of developing successful bone vascularization treatments that can be effectively translated form bench to bedside. Nevertheless, the clear importance of vascularization for bone growth as shown by numerous in vitro and in vivo experiments necessities that research continues to progress in this field.
Conclusion
Vascularization plays a key role in bone growth and healing as suggested by several in vitro and in vivo studies. In vitro approaches strip away the variability of this complex interaction and allow research to penetrate to the fundamental processes in play, whereas in vivo models assess the translational capacity of different treatments by interfacing engineered therapies with the full complement of biological activity. The advent of new analytical tools, imaging techniques, and computational models has accelerated the progress of research and provided additional insights into not only the effectiveness of different treatments, but also the biological means by which these treatments are able to promote vascularization and subsequent bone growth. These techniques have been applied to develop tissue-engineering scaffolds, which support vascularized bone growth, many incorporating osteogenic biomolecules. Although these efforts have resulted in a few FDA-approved clinical therapies such as BMP-2, the needs of the field are far from fulfilled. Alternative therapies should be further investigated, such as combining small molecules with prevascularized scaffolds, or endogenous stem cell homing to defect sites, to exploit the synergistic effects of combined angiogenesis and osteogenesis. Elucidation of the time-dependent relationship between vessel growth and bone repair, standardization and optimization of evaluation tools, and universal adoption of metrics for assessing in vitro assays and other preclinical models are essential for advancing the vascularization techniques that will result in more successful therapies in the future.
Footnotes
Disclosure Statement
No competing financial interests exist.