
Abstract
This review focuses on the stem cell sources with the potential to be used in vascular tissue engineering and to promote vascular regeneration. The first clinical studies using tissue-engineered vascular grafts are already under way, supporting the potential of this technology in the treatment of cardiovascular and other diseases. Despite progress in engineering biomaterials with the appropriate mechanical properties and biological cues as well as bioreactors for generating the correct tissue microenvironment, the source of cells that make up the vascular tissues remains a major challenge for tissue engineers and physicians. Mature cells from the tissue of origin may be difficult to obtain and suffer from limited proliferative capacity, which may further decline as a function of donor age. On the other hand, multipotent and pluripotent stem cells have great potential to provide large numbers of autologous cells with a great differentiation capacity. Here, we discuss the adult multipotent as well as embryonic and induced pluripotent stem cells, their differentiation potential toward vascular lineages, and their use in engineering functional and implantable vascular tissues. We also discuss the associated challenges that need to be addressed in order to facilitate the transition of this technology from the bench to the bedside.
Vascular Tissue Engineering: Unmet Clinical Need
While synthetic vascular prostheses such as expanded polytetrafluoroethylene (ePTFE) and Dacron are available alternatives for high-flow, low-resistance, large peripheral vessel pathologies, their clinical outcome for small-diameter (<6 mm) vessel replacement has been grim.16–20 Prosthetic graft failure has been attributed to intimal hyperplasia, thrombogenicity, compliance mismatch, and diameter mismatch between the graft and native artery.21–24 Despite decades of effort, the successful fabrication of an ideal vascular graft still remains a challenge. Ideally, a vascular graft should be strong, biocompatible, nontoxic, nonimmunogenic, anti-thrombotic, compliant, vasoactive, and amenable to postimplantation remodeling by the host tissue. To this end, tissue-engineered vessels (TEVs) that can withstand the challenging arterial hemodynamic microenvironment and are amenable to physiological remodeling represent an attractive alternative.
Vascular Tissue Engineering Approaches
Three major approaches have been proposed for the tissue engineering of vascular grafts: (1) decellularized matrices; (2) cell-sheet engineering; and (3) biodegradable scaffolds from natural or synthetic polymers.
Decellularized blood vessels as well as small intestinal submucosa (SIS) have been used to fabricate vascular grafts. The main advantage of using decellularized tissue is that the native three-dimensional (3D) architecture of matrix molecules—mainly type 1 collagen and elastin—is preserved 25 and might be helpful in guiding tissue repair and remodeling postimplantation. Decellularized blood vessels provide an intact tubular acellular scaffold that can be implanted either directly or after the addition of endothelial and smooth muscle cells with the aim of improving patency.26–30 Similar to decellularized vessels, native decellularized tissues, that is, SIS, demonstrated sufficient mechanical strength as a vascular graft in vitro 31 and in vivo.32–35 SIS was recently shown to support the attachment and proliferation of smooth muscle cells (SMCs) as well as endothelial cells (ECs) and has been successfully used for the development of mechanically robust TEVs after only 2 weeks in culture. 36
Cell-sheet engineering was pioneered by L'Heureux et al. 37 and requires no scaffold. Instead, it relies on the ability of the cells to form highly interconnected sheets that can be cultured around a mandrel for several weeks, yielding multi-layered cylindrical tissues which can sustain burst pressure up to 2000 mmHg.37,38 Notably, cell-sheet engineered TEVs showed promising clinical results as arteriovenous shunt grafts in a challenging population of dialysis patients with end-stage renal disease.39,40 This impressive milestone represents a significant step forward in the clinical application of tissue-engineered vascular grafts.
Synthetic and natural polymers have been used as scaffolds that support cell growth and provide mechanical support necessary for implantation. In a pioneering study, Niklason et al. first demonstrated the feasibility of engineering a mechanically robust and implantable tissue-engineered blood vessel. 41 Using a polyglycolic acid (PGA)-based scaffold that was molded into a cylindrical shape and seeded with ovine or porcine SMCs on the outer layer and autologous ECs in the lumen, they obtained vascular constructs with a high burst pressure (2150 mmHg) after 8 weeks in culture. The robust mechanical properties of these tissues enabled implantation into the right saphenous artery of Yucatan miniature pigs, where they remained patent for 4 weeks. Since then, many groups employed synthetic polymeric materials, including co-polymers of PGA with poly-L-lactic acid, polycaprolactone, poly-4- hydroxybutyrate, and polyurethane with various degrees of success.42–45 Recently, Dahl et al. also exploited the extracellular matrix (ECM) secretion potential of SMCs in order to fabricate off-the-shelf TEVs. 46 They seeded human cadaveric allogeneic SMCs into rapidly degradable polyglygolic acid scaffolds under the cyclic radial strain to fabricate TEVs that were subsequently decellularized and rendered nonimmunogenic using detergent. Decellularized TEVs could be stored for approximately 1 year at 4°C without compromising their mechanical properties and remained patent either for 1 year after implantation as CABG (with autologous ECs) in dogs or for 6 months as arteriovenous conduits (without ECs) in baboons. 46 These acellular TEVs underwent significant remodeling in vivo, as host alpha smooth muscle actin (α-SMA)-positive SMCs or fibroblasts populated the grafts and secreted elastin, predominantly near the anastomotic sites. However, very few α-SMA-positive SMCs or fibroblasts were seen, and no elastin was present within the mid sections of the grafts, suggesting limited cell infiltration and remodeling capacity, especially in the middle of longer grafts. 46 This approach has the potential to generate nonimmunogenic TEVs from the allogeneic sources, but their clinical utility has not yet been established.
Natural biomaterials such as collagen and fibrin have also been employed, because they can polymerize in the presence of cells and contain inherent biological signals that influence cellular activity. Collagen gels have been utilized, as collagen is both a major component of the ECM of blood vessels and a natural cell substrate. 47 Although the application of cyclic strains improved cell alignment, matrix remodeling, and mechanical strength of the collagen-based constructs,48–50 the mechanical strength remained low for implantation. In contrast to collagen, fibrin stimulated the synthesis of collagen and elastin and yielded TEVs with improved mechanical properties,51–53 suggesting that fibrin may be a more appropriate scaffold for cardiovascular tissue engineering. Indeed, long-term culture of fibrin-based TEVs for 7–9 weeks yielded grafts with a burst pressure between 1400 and 1600 mmHg and a compliance comparable to that of native arteries, suggesting that fibrin-based vascular constructs might be sufficiently strong for arterial implantation. 54 Using fibrin hydrogels, our group demonstrated that fibrin-based small-diameter TEVs could be implanted as interpositional grafts in the jugular veins of lambs, where they remained patent for 15 weeks and displayed significant matrix remodeling, especially when prepared using bone marrow (BM)-derived stem cells.55,56
Regardless of the scaffold that is being employed, engineering the correct mechanical microenvironment that mimics the cyclic stresses experiencing a vessel in vivo is necessary to enhance the function of vascular grafts and improve implantability.41,54 Ultimately, any strategy for engineering vascular substitutes aims at producing TEVs with adequate mechanical properties as well as blood compatibility and immunity characteristics, which are essential for long-term patency. Table 1 summarizes the advances in engineering functional and implantable TEVs.
TEVs, tissue engineered vessels; SMCs, smooth muscle cells; EC, endothelial cells; PTFE, polytetrafluoroethylene; PGA, polyglycolic acid; BMCs, bone marrow cells; hTERT, human telomerase reverse transcriptase subunit; BM-MSCs, bone marrow derived mesenchymal stem cells; BM-MNCs, bone marrow mononuclear cells; PEUU, poly(ester urethane) urea; MD-SCs, muscle derived stem cells.
Despite significant progress made in the development of biomaterials and the correct microenvironment for engineering 3D vascular constructs, cell sourcing remains a major problem. In particular, by increasing donor age, the cell's proliferative capacity and ECM synthesis decrease, leading to decreased mechanical properties such as elasticity and burst pressure.67,73 This is particularly important, as the majority of patients in need of a vascular implant are elderly. To overcome the replicative barrier of adult cells, the human telomerase reverse transcriptase subunit (hTERT) gene was expressed in ECs and SMCs from aged individuals, yielding an enhanced proliferation potential of these cells in culture but failed to reverse aging-associated cellular senescence and decreased life span. 67 Given the limited proliferative capacity and cellular senescence-associated changes in the cell's phenotype, it is necessary to seek alternative autologous cell sources that may be better candidates for TEVs. In this direction, multipotent adult stem cells as well as pluripotent stem cells are being currently explored as alternative cell sources for engineering cardiovascular tissues. In the next sections, we discuss various types of stem cells, their differentiation potential toward vascular lineages, and the challenges remaining before clinical applications can be realized.
Stem Cell Sources for Vascular Tissue Engineering
Adult multipotent stem cells
In this section, we discuss adult stem cells that have been used as sources of vascular cells, SMCs and ECs, for promoting vascularization and for engineering vascular grafts. In particular, we discuss the vascular cells derived from adult stem cells from BM, adipose tissue, hair follicle (HF), umbilical cord (UC), and muscle. In a separate section, we also discuss the endothelial progenitor cells (EPCs) that may be useful for endothelializing the lumen of vascular grafts. These efforts are summarized in Table 2.
BM-SCs, bone marrow-derived stem cells; MNCs, mononuclear cells; MSCs, mesenchymal stem cells; AD-SCs, adipose-derived stem cells; HF-SCs, hair follicle-derived stem cells; UC-SCs, umbilical cord-derived stem cells; MHC, myosin heavy chain; FBS, fetal bovine serum; FLK1, fetal liver kinase 1; VEGF, vascular endothelial growth factor; TGF-βl, transforming growth factor beta l; bFGF, fibroblast growth factor basic; ECGS, endothelial cell growth supplement; PDGF-BB, platelet derived growth factor BB; EGM2, endothelial growth medium 2; α-SMA, smooth muscle alpha actin; ITS, Insulin-Transferrin-Selenium; eNOS, endothelial nitric oxide synthase; Dil-Ac-LDL, Acetylated Low Density Lipoprotein, labeled with l,l′-dioctadecyl -3,3,3′,3′-tetramethyl-indocarbocyanine perchlorate; EPCs, endothelial progenitor cells; UCB, umbilical cord blood.
BM stem cells
Bone marrow mononuclear cells (BM-MNCs): BM is the home of hematopoietic stem cells (HSCs) that sustain hematopoiesis and multipotent mesenchymal stromal cells (also known as mesenchymal stem cells [MSCs]), which are thought to support and maintain the HSC niche. 92 MSCs are of mesodermal origin and constitute 0.1% to 0.5% 93 of the BM-MNCs, which are obtained by discontinuous density gradient centrifugation of BM aspirates.92,94 It was estimated that each gram of BM aspirate can give rise to ∼3×105 plastic adherent cells 95 and that 4–5 mL/kg body weight marrow can be easily aspirated from patients, 62 thereby providing a rich source of stem cells for cell therapies.
BM-MNCs lack a major histocompatibility complex II as well as other immune co-stimulatory molecules, including CD40, CD80, and CD86.96,97 BM-MNCs were also shown to inhibit humoral and cellular immunity, as they suppressed the proliferation of B and T lymphocytes and the maturation of antigen presenting (dendritic) cells,98–100 suggesting that it might be possible to employ allogeneic BM-MNCs for the preparation of vascular grafts. When injected into mice blastocysts, BM-MNCs contributed to the development of all three germ layers, suggesting that these cells may be pluripotent. 101 Indeed, BM-MNCs have been shown to differentiate into neurons, 102 hepatocytes, 103 and cardiomyocytes, 104 indicating a broad differentiation potential.
The first evidence that BM-MNCs contribute to in vivo angiogenesis by differentiating into ECs and SMCs is derived from the studies conducted by Matsumura et al. 63 After removing blood cells, the cells from BM aspirates were labeled with a green fluorescent tracing dye and seeded onto polymeric scaffolds that were implanted in the inferior vena cava of dogs. 63 An immunohistochemical analysis of the explanted TEVs confirmed that labeled BM-MNCs had differentiated into ECs and SMCs. 63 SMCs and ECs were also derived from canine BM-MNCs and used to populate decellularized canine carotid arteries. 68 These TEVs had a suture strength that was similar to native carotid arteries and when implanted into a canine model, remained patent for 8 weeks. In another approach, Ross et al. derived multipotent adult progenitor cells (MAPCs) from BM-MNCs after removal of the CD45+/Glycophorin A+ hematopoietic fraction. MAPCs were shown to be multipotent, and they were successfully coaxed to differentiate into contractile SMCs using a combination of transforming growth factor beta l (TGF-β1) and platelet-derived growth factor BB (PDGF-BB). 74
In a pioneering study, Shin'oka et al. started a clinical trial using TEVs that were prepared by seeding BM-MNCs in the operating room to avoid long culture times.60,62 To this date, 25 children with single-ventricle physiology have been implanted with such TEVs with no evidence of graft-related mortality and limited stenosis, as observed after a mean follow-up time of 5.8 years. This is a very significant step forward, demonstrating the clinical application of TEVs in the low-pressure circulation of young patients. It remains to be seen whether the same approach can be used to repair the arterial vasculature of adult patients.
Bone marrow-derived mesenchymal stem cells (BM-MSCs): When plated on plastic culture flasks, the mesenchymal stromal fraction of MNCs adhere to the plastic and give rise to fibroblastic colonies, while hematopoetic precursors do not adhere to the surface and are subsequently removed.92,94 As defined by the International Society for Cellular Therapy (ISCT), MSCs are adherent fibroblastic cells that are positive for CD105, CD90, and CD73 and negative for CD34, CD45, and CD14 or HLA-DR, CD19, CD11b, and CD79a. 105 In terms of differentiation potential, MSCs differentiate along the osteogenic, chondrogenic, and adipogenic lineages. 105 Besides BM, MSCs have been isolated from different tissues, including blood, adipose tissue, HFs, muscle, lung, spleen, and liver.94,101,106–111 Interestingly, a recent hypothesis postulated that the origin of MSCs may be in the perivascular space of all tissues throughout the body, where they promote tissue repair and homeostasis in a swift and highly localized manner. 112
BM-MSCs were used with nanofibrous poly (L-lactic acid) (PLLA) to fabricate TEVs that were implanted as interpositional grafts into rat common carotid arteries. BM-MSCs-based grafts remained patent for 2 months and demonstrated extensive remodeling, with α-SMA+ cells in the medial layer and CD31+ cells in the lumen, suggesting that ECs from the host might have homed to the grafts. 70 Another study made use of GFP to label BM-MSCs before implantation and clearly demonstrated that the ECs covering the lumen of the TEV grafts originated from the host and not from the implanted BM-MSCs. 113 In agreement, when co-implanted in immunodeficient mice, human BM-MSCs and blood-derived EPCs formed a patent microvasculature, while the implantation of BM-MSCs or EPCs alone failed to form vessels. 75 Taken together, these studies suggest that in contrast to BM-MNCs, which may contain endothelial progenitors, BM-MSCs have very limited—if at all—EC differentiation potential, but they serve in stabilizing the microvasculature generated by ECs. Finally, it remains to be been seen whether the findings in rodent animal models can be translated into more physiologically relevant large animal models—a very challenging task.
Human BM-MSCs have been used as a source of SMCs as well as for TEVs fabrication. The Niklason group studied the role of ECM, growth factors, and mechanical forces in human BM-MSCs differentiation toward SMCs and highlighted the importance of combining mechanical and chemical factors to fabricate the vascular wall. 77 In our laboratory, we employed a tissue-specific promoter (α-SMA) and flow cytometry to purify SMCs from BM-MSCs and used them to engineer vascular grafts, which exhibited robust vascular contractility in response to receptor and non-receptor-mediated vasoconstrictors. 55 When implanted into the jugular vein of an ovine animal model, the grafts remained patent for 5 weeks and demonstrated extensive remodeling capacity. In contrast to vascular SMCs from the jugular veins of neonatal lambs, BM-MSCs-derived SMCs synthesized high amounts of collagen and, most notably, fibrillar elastin similar to native veins, indicating the great potential of these cells for regenerating the vascular wall.
Adipose-derived stem cells
Adipose-derived stem cells (AD-SCs) were first isolated from the stromo-vascular fraction (SVF) of adipose tissue harvested from liposuction. 114 A few hundred milliliters to a few liters of adipose tissue is routinely liposuctioned from patients, and the lipoaspirate is subsequently digested with collagenase, washed, and filtered. AD-SCs are derived from the filtrate, also known as processed lipoaspirate, after culture for several days in the presence of fetal bovine serum. 114 One group obtained 2×105 plastic adherent cells per gram of lipoaspirate 95 and ∼1.5×1010 AD-SCs from 1 L of lipoaspirate after two passages, 95 and others showed that AD-SCs could undergo ∼34 population doublings in culture, suggesting a high proliferation capacity. 115 AD-SCs have a fibroblastic morphology, MSCs-like immunophenotype (CD34+/CD105+/CD45−/CD31−, 116 or CD29+/CD44+/CD71+CD90+/CD105+/STRO-1+/CD49d+/CD31−/CD34−/CD45− 109 ) and exhibit a clonal differentiation potential along all mesenchymal lineages. 114 In addition, several studies demonstrated that the differentiation potential of AD-SCs extends beyond mesodermal boundaries and includes neurons,117–119 hepatocytes,120,121 and pancreatic islets.122,123 Finally, AD-SCs were found to suppress the mixed lymphocyte reaction and lymphocyte proliferation, supporting their potential for allogenic transplantations. 124
AD-SCs are a rich source of vascular cells. Several studies demonstrated the successful differentiation of AD-SCs into contractile SMCs,78–80 which compacted hydrogels and proliferated on decellularized saphenous vein scaffolds, indicating their potential for reconsituting the vessel wall. 78 More recently, SMC were derived from AD-SCs by treatment with TGF-β1 and bone morphogenetic protein 4 (BMP4) and were used on a PGA scaffold to generate the vascular wall. 125 Interestingly, the application of pulsatile force significantly improved both collagen synthesis and vessel wall mechanics 125 ; however, the implantability of these vascular grafts was not demonstrated.
Several groups reported that AD-SCs are a rich source of ECs.80,126 In addition to the total AD-SCs population, the CD34+/CD31− cell fraction from the gluteal, abdominal, and visceral adipose tissue of human donors expressed endothelial markers and enhanced vascularization after implantation in a hind limb ischemia mouse model. 127 Another cell fraction, the FLK1+/CD31−/CD34−/CD106− cells from SVF could also be coaxed to differentiate into ECs by culture on matrigel-coated dishes in the presence of vascular endothelial growth factor (VEGF) and fibroblast growth factor basic (bFGF). 85 Interestingly, the EC differentiation capacity of AD-SCs was not affected by either donor aging or vascular disease.83,128 Some studies suggested the presence of a common progenitor among AD-SCs that are capable of differentiating along both mesenchymal and endothelial lineages.126,129 However, others showed that in AD-SCs cultures, the CD31 and CD144 endothelial-specific promoters were methylated even after stimulation with EC-promoting growth factors, suggesting that the differentiation potential of AD-SCs toward endothelial fate might be epigenetically limited. 130 Given the heterogeneity of the SVF, it is not clear whether AD-SCs-derived ECs either differentiate directly from a common multipotent progenitor or represent a population of EC progenitors in SVF. Nevertheless, taken together, these studies support the notion that AD-SCs constitute an important stem cell source for vascular regeneration.
HF-derived stem cells
The HF is a dynamic mini organ containing cells of ectodermal and mesodermal origin. Anatomically, HF can be divided into four regions of ectodermal origin: (1) infundibulum; (2) isthmus; (3) suprabulbar region; and (4) bulb. Throughout life, HFs undergo many cycles of growth (anagen), regression (catagen) and quiescence (telogen), suggesting the presence of stem cells that support the process of continuous regeneration. However, the anatomic location where stem cells reside remained elusive until the early 1990s, when the first experiments were designed to follow the fate of label-retaining cells, showing that these cells resided in the bulge region of the isthmus. 131 This finding was later verified with sophisticated cell-fate mapping experiments132,133 that established the bulge as the stem cell niche of the HF. The multipotency of hair follicle-derived stem cells (HF-SCs) has also been demonstrated by coaxing them to differentiate into corneal epithelial cells, 134 melanocytes,88,135 neural stem cells,88,135 Schwann cells, 136 neurons,137,138 and myelinating glial cells. 137
The HF is surrounded by the dermal sheath (DS), which contains progenitor cells that maintain and regenerate the dermal papilla (DP), a mesodermal structure underneath the bulb region, which is very important for promoting hair regeneration.139,140 The DP and DS of rat HFs were shown to contain MSCs (termed HF-MSCs),141,142 which showed similar proliferation and differentiation potential as rat BM-MSCs. 143 Notably, HF-MSCs showed hematopoietic differentiation potential, as displayed by transplantation experiments in which cultured DP cells repopulated the BM of lethally irradiated mice. 144 In our group, we showed that the immunophenotypes of human HF-MSCs are similar to those of BM-MSCs (CD73+/CD105+/CD44+/CD49b+/CD90+/CD309−/CD144−/CD34−/CD45−) and that they are clonally multipotent, as individual cells can be coaxed to differentiate into fat, bone, cartilage, and muscle cells. 145 We also noted that HF-MSCs were less susceptible to culture senescence than BM-MSCs, as they could undergo ∼45 population doublings and maintained their myogenic differentiation potential as exhibited by the development of contractile properties even at late passages. Using a tissue-specific promoter (α-SMA) and flow cytometry sorting, we purified a proliferative population of ovine HF-MSCs that could be coaxed to differentiate into SMCs, as demonstrated by the expression of α-SMA, calponin, and myosin heavy chains as well as significant contractility in response to vascular agonists, suggesting that these cells were functional SMCs.86,87 When combined with the natural scaffold, SIS, HF-MSCs-derived SMCs generated vascular media with significant vascular contractility and mechanical properties approaching those of native arteries, 146 demonstrating the potential of HF-MSCs for use in vascular tissue engineering. Indeed, cylindrical TEVs generated with this approach in our laboratory are currently implanted as interpositional grafts in the arterial circulation of an ovine animal model (unpublished data).
We estimated that ∼1016 HF-MSCs can be obtained from single HF 145 and given the ease of accessibility and the high hair density on adult scalp (175–300 hairs/cm2; 147 ), HF-MSCs may be considered a robust autologous stem cell source for vascular progenitors. In an intriguing transgender experiment conducted on humans, where DS cells from two male donors were transplanted to a female recipient, it was observed that grafted DS/DP cells gave rise to intact HFs without immune rejection. 148 If proved to be true, this finding is very exciting, as it suggests that akin to BM-MSCs, HF-MSCs may also be immunoprivileged, thereby enhancing their potential as a readily available, allogeneic stem cell source for engineering cardiovascular tissues.
UC-derived stem cells
The UC connects the developing fetus with the placenta and is composed of three vessels—two umbilical arteries and one umbilical vein—surrounded by gelatinous mucopolysaccharides called Wharton's jelly. At term, newborn's UC is ∼30–50 cm long and weighs ∼50 g. 149 Both umbilical cord blood (UCB) and umbilical cord tissue are repositories of multipotent stem cells. Approximately 75 mL of UCB can be obtained from each UC at birth that reportedly contains a heterogeneous cell population, including HSCs,150,151 MSCs, 152 unrestricted somatic stem cells, 153 multilineage progenitors, 154 endothelial progenitors, 155 and very small embryonic-like stem cells. 156 Interestingly, UCB-derived HSCs have been in clinical use for the past 20 years for the treatment of malignant hematopoietic disorders. 150 MNCs, derived from UCB, have been shown to be MSCs that are based on plastic adherence, immunophenotypic profile, and multilineage differentiation potential toward adipogenic, chondrogenic, and osteogenic lineages.152,157 It was reported that 108 cord blood MNCs can give rise to 0.7±0.2 MSC colonies, 158 and these cells can undergo approximately 20 population doublings in culture. 159 In addition, endothelial progenitors have been isolated from UCB160,162 using CD34+, 160 CD133+,163,164 CD34+/CD133+ 162 and AC133−/CD14+ 161 surface markers, and their angiogenic potential has been demonstrated in vivo.155,163,164 Pesce and colleagues reported that 83% CD34+ cells were present among the MNCs of UCB, 162 suggesting that UCB is a rich source of EPCs. UCB is amenable to cryopreservation and can serve as an autologous source of UCB-derived MSCs and EPCs. 165
The UC tissue and Wharton's jelly have been shown to possess MSCs.149,157 Specifically, MSCs have been isolated from umbilical artery, 166 umbilical vein, perivascular site, 167 Wharton's jelly, 168 and whole UC 169 and are collectively denoted here as UC-MSCs. Approximately 1.75×105 MSCs can be obtained per gram of UC tissue (total 5–6 g of UC tissue is obtained from each UC) that can reportedly undergo ∼55 population doublings. 149 Approximately 1010 UC-MSCs can be obtained from 2–5×106 cells after 30 days of culture, suggesting that UC-MSCs represent a highly proliferative stem-cell source. 149
Besides mesenchymal lineages,149,157 UC-MSCs were shown to differentiate into cardiomyocytes, 170 hepatocytes, 171 and neurons. 172 In addition, UC-MSCs were shown to differentiate toward endothelial and myogenic cells in vitro. When cultured on matrigel-coated surfaces in the presence of bFGF and VEGF, UC-MSCs differentiated into ECs, which formed neo-vessels on implantation into the ischemic limbs of nude mice. 89 Farias et al. demonstrated that CD10+/CD29+/CD44+/CD73+/CD90+/CD105+/CD34−/CD45− UC-MSCs were ultra-structurally and functionally very similar to myofibroblasts, as they contracted collagen gels on cytokine treatment. 90 The myogenic differentiation potential of Wharton's jelly-derived MSCs (CD105+/CD31−/KDR−) was demonstrated by their ability to promote healing of the rat tibialis anterior muscle that was damaged by bupivacaine treatment. 173 Similar to other MSCs, UC-MSCs have also been shown to possess immunomodulatory properties, suggesting that they may be suitable for allogeneic transplantations.174,175 However, their potential to generate contractile and implantable TEVs is yet to be demonstrated.
Muscle-derived stem cells
Muscle-derived stem cells (MD-SCs) are another accessible cell source for regenerative medicine. These are the quiescent cells residing beneath the basal lamina of muscle fibers, 176 which are activated on injury and participate in the repair process. 177 MD-SCs were isolated from muscle biopsies and although the majority demonstrated robust myogenic differentiation potential, an Sca-1 expressing fraction of late passage cells also exhibited tri-lineage differentiation potential in fat, bone, and cartilage.91,178,179
Vascular grafts that were engineered using rat MD-SCs and polymeric poly(ester-urethane) urea scaffolds showed impressive mechanical properties with a burst pressure reaching ∼2000 mmHg. 180 These TEVs remained patent and sustained arterial blood flow up to 8 weeks postimplantation as interpositional grafts in rats, 71 suggesting that MD-SCs may be potentially useful as a source of autologous SMCs for vascular tissue engineering.
Endothelial progenitor cells
EPCs were initially isolated from peripheral blood as CD34+ or Flk1+ cells that differentiated into functional ECs in vitro. 181 EPCs are produced in BM182,183 and are mobilized into the circulation by VEGF, 184 vascular trauma, 185 and several other cytokines. 186 EPCs have also been isolated from UCB160–164 and were shown to have enhanced proliferation potential as compared with those originating from peripheral blood. 187 Three strategies have been employed in the past to isolate EPCs: (1) adhesion to fibronectin-coated surfaces in conjunction with the ability to uptake the lectin Ulex Europeaus agglutinin-1 and fluorescence-labeled acetylated low-density lipoprotein (acLDL)181,188; (2) CD34+/FLK1+/CD133+ mmunophenotype181,189; and (3) the capacity to form cobblestone colonies, 190 which were shown to correlate inversely with the risk of CAD.190,191 Accumulating evidence suggests that EPCs derived from BM or peripheral blood may represent hematopoietic lineage cells at various stages of maturation and with varying angiogenic potential, 192 providing a potential explanation for the lack of standardized criteria and cell surface markers to define and isolate them.
Regardless of the method of isolation, EPCs have been shown to promote neo-angiogenesis. 193 The delivery of EPCs—either by a direct injection or after the seeding on scaffolds—enhanced the angiogenesis in animal models of ischemia or infarction.194–197 Shi et al. demonstrated that CD34+ EPCs from BM-colonized vascular prostheses in vivo, 182 suggesting their potential application in vascular regeneration. In another study, Kaushal et al. isolated EPCs from ovine peripheral blood and seeded them onto decellularized porcine iliac vessels that were implanted into an ovine animal model. 27 The implanted vascular grafts remained patent for 130 days, suggesting that the EPCs were successfully substituted for mature ECs in the TEVs lumen. 27
EPCs are rare cells that are present in the blood at a frequency of 1 in 10,000 cells 198 or according to other reports as low as one cobblestone EPCs colony per 108 MNCs. 192 In addition, EPCs can only undergo 20–30 population doublings in vitro, 187 and their proliferation capacity was shown to further decline with age198,199 and cardiac disease, for example, CAD,190,191 further limiting their use in vascular regeneration. However, a recent report showed that the peroxisome proliferator-activated receptor-gamma agonist, rosiglitazone prevented the decline of EPC function against the accumulation of advanced glycation end products. 200 More studies are needed in this direction to better understand the molecular underpinning of aging and to develop strategies that attenuate the effects of senescence and realize the clinical potential of EPCs for vascular regeneration.
Challenges and limitations of multipotent stem cells
Maybe the most challenging obstacle hampering clinical applications of adult stem cells is cellular senescence, which is the decreased proliferation and differentiation potential by increasing donor age. 201 In addition to organismal aging, MSCs also suffer from culture senescence, limiting their culture time to about 8–10 passages and preventing their expandability to the large cell numbers required for cellular therapies. This is a major concern, as the patients who are mostly in need of cellular therapies, in general, and of vascular grafts, in particular, are elderly.
Age-related decline in cell number has been reported for BM-MSCs202,203 and AD-SCs. 204 Our laboratory demonstrated that SMCs differentiated from adult BM-MSCs were not only less proliferative but also significantly less contractile than their neonatal counterparts. 205 In another study, the Niklason group showed that terminally differentiated vascular cells isolated from aged patients exhibited reduced functionality and decreased ECM remodeling, resulting in failed grafts. Although ectopic expression of the hTERT subunit enhanced the proliferation of ECs and SMCs from aged patients, it failed to reverse the aging associated cellular senescence and decreased life span. 67 Interestingly, our recent studies showed that HF-MSCs were more resistant to donor aging compared with BM-MSCs, as evidenced by their ability to proliferate and maintain their vascular differentiation potential for longer times in culture ( 145 and unpublished data). In addition, the neutralization of insulin-like growth factor-1 reversed the aging defects in the HSC niche in the BM 206 and pulsed the administration of rapamycin-rejuvenated HSCs in aged mice, 207 but their effects on MSCs have not been evaluated as yet. More studies are required to understand the molecular underpinning of cellular senescence, which may lead to the design of rational treatments that delay or even reverse the loss of cellular function.
Increasing the use of adult stem cells in vascular tissue regeneration also calls for a better understanding of their interactions with host cells in balancing the inflammatory response and promoting tissue remodeling. Observations from vascular implants in rats suggested that MSCs remained in the grafts for a limited time period, as they were gradually replaced by host cells, 70 suggesting that the presence of cells in the vessel wall may not be necessary for graft patency. However, a recent study showed that the cell presence in the wall decreased macrophage infiltration and reduced the incidence of graft stenosis. 208 The inhibition of macrophage infiltration also blocked the formation of vascular neo-tissue, as evidenced by the absence of endothelial and smooth muscle cells, suggesting that macrophages play a critical role in vascular graft remodeling. This study also suggested that BM-MSCs might play an important role in promoting graft remodeling by balancing the immune response, an interpretation that is compatible with the notion that MSCs may be immuno privileged. Notably, the clearance of MSCs from the implants within a few weeks postimplantation strongly supports the use of allogeneic stem cells for engineering TEVs, as immunosuppression medication may need to be administered for a limited time postsurgery. The use of allogeneic cells will certainly facilitate the commercialization of TEVs and, therefore, the translation of this technology into the clinic.
Pluripotent stem cells
Embryonic stem cells and induced pluripotent stem cells
Pluripotency refers to the cell's ability to give rise to all three germ layers, that is, ectoderm, mesoderm, and endoderm. 209 In 1981, researchers developed a technique to grow a pluripotent inner cell mass of the blastocyct mouse embryo in vitro, and these cells were termed mouse embryonic stem cells (mESCs). 210 Almost two decades later, the Thomson group established similar techniques for culturing human ESCs. 211
In a breakthrough study conducted in 2006, Yamanaka and colleagues demonstrated that mouse embryonic fibroblasts and adult fibroblasts acquire properties similar to ESCs when transduced with retroviruses containing four transcription factors, OCT4, SOX2, KLF4, and cMyc212–215
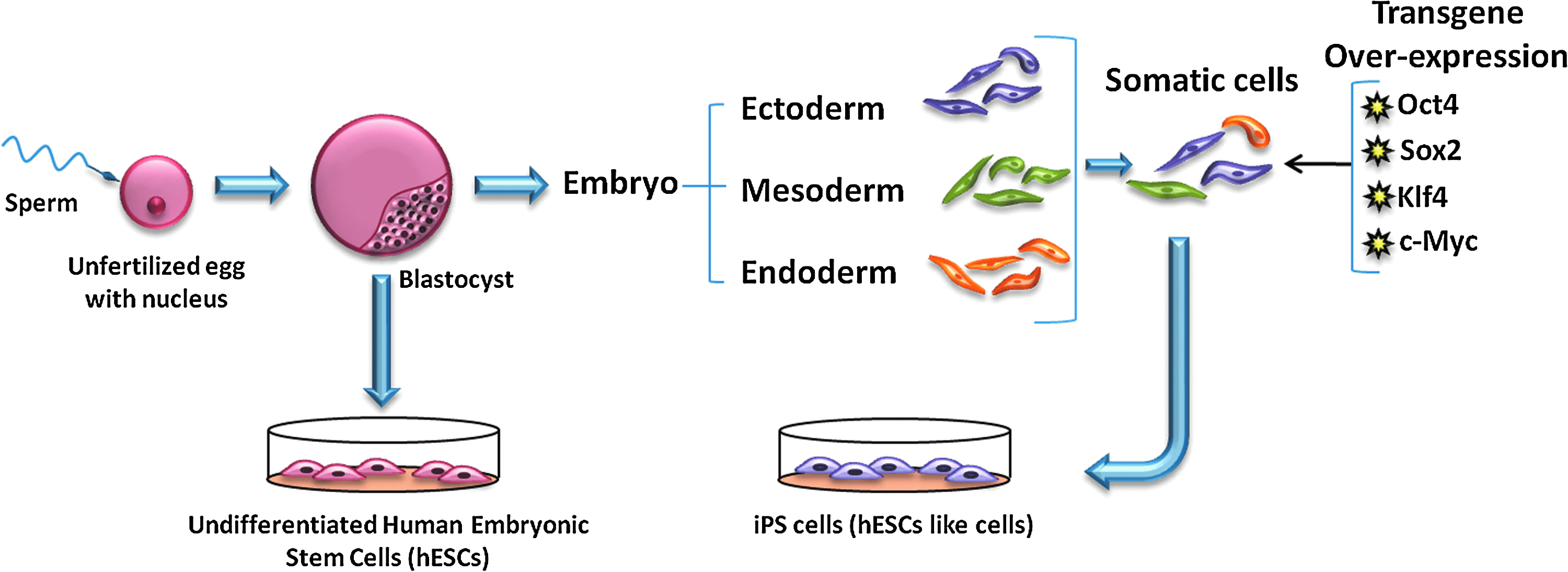
Schematic diagram of pluripotent stem-cell derivation. iPS cells, induced pluripotent stem cells. iPS cells, induced pluripotent cells. Color images available online at www.liebertpub.com/teb
Pluripotent stem-cell differentiation strategies
Three strategies have been employed for directed differentiation of pluripotent stem cells

Schematic diagram showing the strategies for in vitro differentiation of pluripotent stem cells. FACS, fluorescence-activated cell sorting. Color images available online at www.liebertpub.com/teb
Pluripotent stem-cell differentiation toward vascular cells
Several studies involving vertebrate model systems, primarily mice and zebra fish, identified the key signaling pathways involved in vascular development.
244
Others studied pluripotent stem cell differentiation toward vascular fate in vitro and determined the growth factors and timing schedule required for vascular specification in vitro

Schematic diagram depicting the pathways of human pluripotent stem cell differentiation toward vascular fate. Positive and negative markers and functional criteria for particular lineages are represented in italics. The time for each differentiation step is depicted in capital letters. Studies using the embryoid body, stromal cell co-culture, or monolayer culture methods are represented using thin lines, broken lines, and thick lines, respectively. FGF, fibroblast growth factor; BMP4, bone morphogenetic protein 4; TGF-βl, transforming growth factor beta l; PDGF-BB, platelet-derived growth factor BB; SCF, stem cell factor; VEGF, vascular endothelial growth factor.
Human pluripotent stem cells have been successfully differentiated in vitro toward ECs258–263 and SMCs.258,260,264 Using the EB method, both vascular cell types (ECs and SMCs) could be derived from hESCs and could generate vasculature on implantation into mice. 265 Using the same method, it was shown that the sequential treatment of EBs with BMP4, Activin A, FGF2, VEGFA, and SB431542 (a TGFβIIR inhibitor), resulted in a robust EC fate. 263 The co-culture of hESCs with OP9 cells for 8 days also yielded vascular progenitors (FLK1+/TRA-1-60−), which on sorting and culture on collagen IV could differentiate to either SMCs (CD31−/α-SMA+) or ECs (CD31+), on treatment with serum or VEGF, respectively. 260 In another approach, simply treating monolayer cultures of hESC with serum resulted in CD34+ cells, which could be coaxed to differentiate further into EC in the presence of VEGF-A and FGF2. The differentiated ECs were functional, as demonstrated by their capacity to form blood vessels when co-implanted with 10T1/2 cells in nude mice. 266 A recent study took this approach a step further by demonstrating that a chemically defined medium could induce monolayer cultures of pluripotent stem cells that give rise to the mesoderm and neuroectoderm, which on further treatment with PDGF-BB and TGF-β1 yielded functional SMCs. 267 Interestingly, hiPSCs-derived ECs were shown to have a limited proliferation potential as compared with hESCs-derived ECs, suggesting that reprogramming or the source of hiPSCs might affect the regeneration potential of differentiated vascular cells. 268 More studies are needed to examine the potential of hiPSCs-derived vascular cells to generate functional and implantable small-diameter vascular grafts. Table 3 summarizes the studies in the direction of the vascular differentiation of pluripotent stem cells, including selection criteria and the necessary soluble factors.
ESCs, embryonic stem cells; mESCs, mouse embryonic stem cells; miPSCs, mouse-induced pluripotent stem cells; hESCs, human embryonic stem cells; hiPSCs, human-induced pluripotent stem cells; EB, embryoid bodies; T, brachyury; SFM, serum-free media; BMP4, bone morphogenetic protein 4; Pax6, paired box gene 6; GFP, green fluorescent protein, PDGFR-α, platelet-derived growth factor receptor alpha; SCL, stem cell leukemia protein; SCF, stem cell factor; IL3, interleukin3; EPO, erythropoietin; G-CSF, granulocyte colony stimulating factor; SMGM2, smooth muscle growth medium 2; TRA-1-60, tumor rejection antigen 1–60; mSCF, mouse stem cell factor; mIL3, mouse interleukin3; hTPO, human thrombopoietin; hEPO, human erythropoietin; SB431542, TGF-β1 receptor ALK5 inhibitor; SMCM, smooth muscle cell medium; atRA, all trans retinoic acid.
Challenges and limitations of pluripotent stem cells
Although pluripotent stem cells such as human (h)ESCs and hiPSCs are great potential cell sources; they also suffer from major disadvantages. hESCs are plagued by ethical concerns regarding their isolation from human embryos as well as potential immunogenicity. On the other hand, hiPSCs are more attractive, because they can be generated from the patient's own cells obviating immunogenic rejection and can differentiate into vascular cells using similar protocols as those developed for ESCs.274,276,278 However, several issues currently prevent the use of hiPSCs in regenerative medicine, including (1) low reprogramming efficiency; (2) use of viral vectors for reprogramming; (3) accumulation of chromosomal abnormalities281,282; (4) oncogenic potential; and (5) variations in differentiation potential depending on the donor cell type. 283
The safety concerns over iPSCs generation by the use of recombinant retrovirus led to the development of novel methods to avoid transgene integration, including standard transfection, viral vectors that can be excised from the genome after integration, mRNA, and protein delivery strategies.215,284–291 Small molecules are being increasingly used and can be substituted for some of the genetic factors, either by affecting histone deacetylases or through other mechanisms.292–299 These studies increased the safety as well as the efficiency of reprogramming and facilitated iPSCs generation from the somatic cells of patients, thereby increasing the potential of iPSCs for studying the molecular mechanisms of disease and for regenerative medicine.
On the other hand, recent studies documented chromosomal aberrations in pluripotent stem cells, resulting either from culture adaptation or the original parental cells.300,301 In addition, iPSCs were found to acquire genetic aberrations such as copy number variation and mutations as a result of the reprogramming process.281,282,302 Finally, iPSCs were recently found to contain on an average five-point mutations in protein coding regions, independently of the method of reprogramming. 282 Therefore, before clinical use, it is necessary to ensure that iPSCs are rendered free from any genetic or epigenetic abnormalities which can compromise the long-term function of implanted cells or tissues. In addition, the epigenome as well as the differentiation potential of iPSCs may depend on the donor cell type, 283 calling for studies to compare the vascular differentiation potential of iPSCs originating from different donor cells. Finally, the development of rigorous approaches for selecting large numbers of differentiated cells from undifferentiated ones is also required to eliminate the tumorigenic potential of the latter before delivery to the patient.
Conclusions
Despite progress in the development of biomaterials and engineering the correct microenvironment with the mechanical and biological cues supporting blood vessel function, the source of cells remains a major challenge in engineering a successful human vascular graft. Multipotent and pluripotent stem cells have tremendous potential for vascular tissue engineering. Autologous adult multipotent stem cells such as MSCs and EPCs have shown robust differentiation potential toward vascular lineages and beneficial attributes in balancing the inflammatory response and promoting neotissue formation. However, their limited lifespan due to organismal or culture senescence and slow rates of ECM synthesis pose challenges in engineering vascular grafts from elderly patients. On the other hand, hiPSCs can be prepared from the patient's own cells and have a significantly more protracted lifespan, but concerns about chromosomal aberrations, oncogenic potential, incomplete differentiation, and cumbersome and lengthy reprogramming strategies limit clinical applications at the moment. Although currently ongoing clinical trials demonstrate the efficacy of TEVs using differentiated autologous vascular cells, the lengthy culture times and the custom-made preparation of the grafts increase the cost of treatment significantly. In this regard, the enhanced proliferation and differentiation potential along with the immune-privileged attributes of stem cells may increase the quality, decrease the time and cost of tissue development, and even facilitate the preparation of off-the-shelf grafts that can be employed on demand.
Footnotes
Acknowledgments
This work was supported by grants from National Institute of Health (ROI HL086582) and the New York State Stem Cell Science (NYSTEM contract #CO24316) to Stelios T. Andreadis.
Disclosure Statement
No competing financial interests exist.