
Abstract
Dental pulp from deciduous (baby) teeth, which are discarded after exfoliation, represents an advantageous source of young stem cells. Herein, we discuss the methods of deciduous teeth stem cell (DTSC) isolation and cultivation. We show that based on these methods, at least three different stem cell populations can be identified: a population similar to bone marrow–derived mesenchymal stem cells, an epithelial stem–like cells, and/or a mixed population composed of both cell types. We analyzed the embryonic origin and stem cell niche of DTSCs with respect to the advantages they can provide for their future use in cell therapies and regenerative medicine. In vitro and in vivo differentiation of the DTSC populations, their developmental potential, immunological compatibility, tissue engineering, and transplantation use in studies in animal models are also the focus of the current report. We briefly describe the derivation of induced pluripotent stem (iPS) cells from DTSCs, which can be obtained more easily and efficiently in comparison with human fibroblasts. These iPS cells represent an interesting model for the investigation of pediatric diseases and disorders. The importance of DTSC banking is also discussed.
Introduction
Deciduous teeth are discarded after exfoliation (natural process of primary tooth root resorption), thus DP collection avoids any type of ethical considerations. DP is easy in isolation, in stem cell extraction, manipulation, and in vitro expansion of the cells obtained. The finding of different deciduous teeth stem cell (DTSC) populations, such as stem cells from human exfoliated deciduous teeth (SHED) and immature dental pulp stem cells (IDPSCs), opened a new window for harvesting young stem cells.2,4 These stem cells can be obtained in sufficient quantities once diverse teeth can be used for DP collection.5,6 The current review is focused on DTSCs. Herein we discuss (1) the embryonic origin and DTSC niche, (2) methods of isolation and cultivation of DTSC different populations, (3) DTSC differentiation in vitro and in vivo, (4) developmental potential of DTSCs through chimeras formation, (5) DTSC use in tissue engineering and in animal models of human diseases, (6) immunological properties of DTSCs, (7) induced pluripotent stem (iPS) cells derivated from DTSCs, and (8) stem cell banking.
To distinguish stem cells isolated from adult DP and deciduous teeth, we suggest to refer to the stem cells from deciduous teeth as DTSCs and those from adult teeth as dental pulp stem cells (DPSCs).
Embryonic Origin of DTSCs
Although stem cells from different sources share similar properties, their fate in vivo, developmental, and therapeutic potential are dependent of their embryonic origin and their eventual anatomic location. Overall, teeth develop from oral ectoderm and neural crest-derived mesenchyme, more precisely from neural crest (a transient embryonic tissue), which gives rise to postmigratory neural crest stem cells (NCSCs). These cells constitute pluripotent, migratory stem cell population that displays developmental potential similar to embryonic stem (ES) cells that are able to differentiate into a spectrum of cell types during mammalian development, such as glia, neurons, bones, tendons, melanocytes, chondrocytes, endocrine cells, adipose cells, and others.25,26 Although the pluripotency of NCSCs becomes restricted during development, their postnatal homologs maintain the capacity to self-renewal and “stemness.” 27 Since only limited number of NCSCs can be isolated from embryo, alternative long-term stem cell cultures, which resemble endogenous neural crest cells, are of great interest. The embryonic origin, clonogenicity, self-renewal and proliferation capacities, expression pattern of specific markers, and multipotency observed in DTSCs suggest their comparative equivalence with NCSCs.
Niche of DTSCs
The stem cell's niche (SCN) is a unique microenvironment within each tissue that regulates stem/progenitor cell proliferation, survival, migration, fate, and aging, through specific interaction between the cells and through multiple bioactive molecules.28,29 Niche formation starts in early development and continues after the birth during childhood, providing cells for growth and self-renewal of the organism (renewing of skin, blood, hair, etc). Additionally, SCN can be sporadically activated in response to diseases and different types of injuries to provide cells for tissue regeneration. SCN is constantly under the influence of heritable genetic and/or epigenetic and/or environmental factors, which play a role in modifying and enhancing their effects during human life. Genetic and epigenetic factors are unique for each individual, while environmental factors are variable. For example, nutrition or excessive UV radiation can randomly lead to metabolic disorders (diabetes) and/or can produce mutations in somatic cells. These events can occasionally induce different abnormalities in cell proliferation, differentiation, and homeostasis as well as in tissue function, for example, melanomas formation. The aging of organism should also be taken into consideration since significant number of stem cells can decrease with age.29–33 Each of these factors can influence stem cell quality within SCN.
The formation of SCN in DP of deciduous teeth starts before birth and SCNs are maintained before eruption of the permanent teeth. This short period of an active growing of the organism is characterized by the maintenance of active SCNs that are rich in stem cells, which are not yet heavily affected by accumulative effect of genetic and/or environmental factors. Indeed, DP from deciduous teeth provides a source of healthy stem cells for the future of cell therapies, when compared with DP isolated from permanent teeth.
Methods of DTSC Isolation
Stem cell development in tissues and also in cell cultures follows a hierarchical pyramidal order of stem/progenitor cell lineage organization and cell fate. Our model of mesoderm lineage commitment and differentiation is an example of BM-MSC hierarchical organization and is a prototype for MSCs derived from other sources.17,34 In this case, DTSCs cultured in vitro are not an exception since they follow similar hierarchical patterns. In cell populations isolated and cultured in vitro, the balance in quantity of stem cell and progenitors at the different stages of commitment directly depends on the method of isolation and cell culture conditions. Accordingly, quality and quantity of fetal bovine serum (FBS) added in basal culture medium, use of enzymatic digestion, addition of growth factors, and other changes introduced into the protocol for stem cell isolation can contribute to selection or propagation of different populations of stem cells and/or progenitors.17,18,35
Following two distinct protocols, different populations of DTSCs were isolated: SHED and IDPSCs2,4 (Fig. 1A, B). Figure 1A demonstrates the original process of SHED isolation. Using enzymatic digestion of DP, single-cell suspensions were obtained. Further, these cells were cultured in basal culture medium, which is similar to that used for BM-MSC culture. This culture medium is composed of Dulbecco's modified Eagle's medium (DMEM) low glucose supplemented with 10% of FBS. Single-cell suspension obtained from one tooth is able to generate approximately 12–20 cell colonies. After a short period of proliferation (∼1 week), these multiple cell colonies were harvested and expanded in vitro as SHED. Flow cytometry (FACS) analysis revealed heterogeneous population of SHED, which contains 9% of STRO-1–positive cells.

Protocols of SHED
Figure 1B demonstrates the process of IDPSC derivation. For IDPSC derivation from DP, tissue explant method was used without enzymatic digestion. The tissue was placed on culture dish and was cultured in basal culture medium similar from that for ES cells (DMEM/F12: Nutrient Mixture F-12) supplemented with 15%–20% FBS (Hyclone), 1% nonessential amino acid, and antibiotics. The first outgrowing cells appear after 7–10 days of in vitro cultivation. Next, the DP explant was mechanically transferred into another culture dish, where it continuously produces the cells. Cells remaining in culture dish were harvested using enzymatic digestion and were expanded or cryopreserved in early passages. Ex vivo expansion of IDPSCs leads to isolation of multicellular population standardized with respect to several ectomesenchymal markers' expression, such as CD105 (SH2), CD73 (SH3), nestin, and others.
Immunophenotyping of DTSCs
As reported in early original publications, SHED is a heterogeneous multicellular population of stem cells. A minor proportion of SHED expressed a STRO-1 protein (9%), which potentially defines a multipotent progenitor subset that is also considered to be a marker of osteoblast precursors36,37 (Table 1). Analysis of single-cell-derived colonies, performed by authors, confirmed the heterogeneity of SHED population. Cell colonies derived from SHED presented morphologically and functionally (cell proliferation and differentiation) variable characteristics. 2 In later studies, a variety of stromal, vascular, bone, and neuronal markers were found to be expressed by SHED, such as alkaline phosphatase, matrix extracellular phosphaglycoprotein, basic fibroblast growth factor (FGF) and endostatin, corebinding factor, runt domain, α subunit 1, osterix, osteocalcin, nestin, βIII-tubulin, glutamic acid decarboxylase, neuronal nuclei, glial fibrillary acidic protein (GFAP), neurofilament M (NFM), and 2′,3′-cyclic nucleotide-3′-phosphodiesterase (Table 1, Fig. 2A). Additionally, the expression of CD146 (MUC18), a 118 kD integral transmembrane glycoprotein, which has been identified as a marker of pericytes, was also observed in SHED.23,38 The expression of CD146 in SHED varies between ∼48% and ∼66% (Table 1).
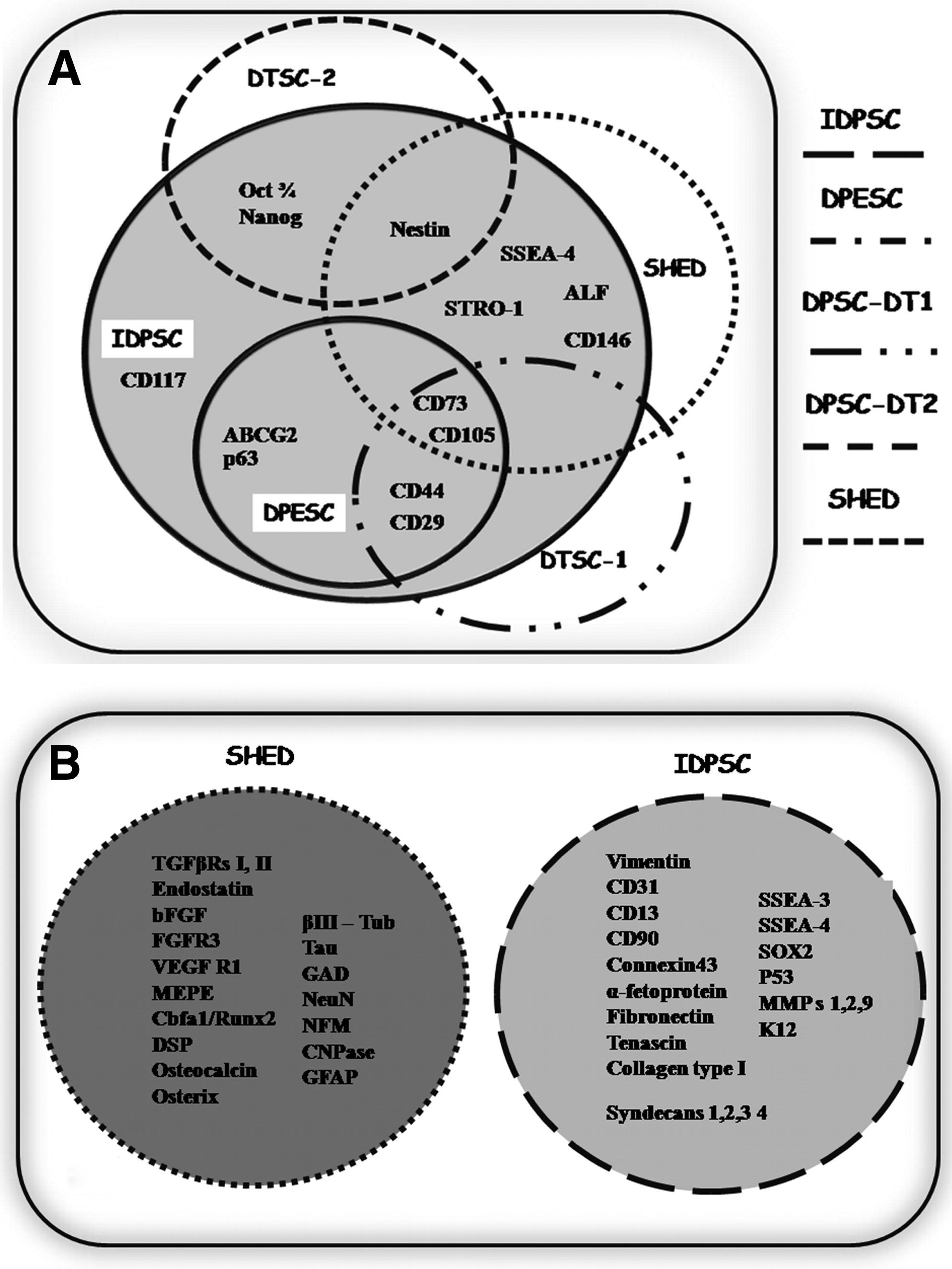
Similarities and differences between protein and gene expression pattern of various DTSC populations.
For the purposes of current review, we denominated stem cell populations described in several recent articles as DTSCs with respective number (exp.: DTSC-1).
n/a–not analyzed, (+) markers found and (−) markers not found yet in DTSCs.
DTSCs, deciduous teeth stem cells; SHED, stem cells from human exfoliated deciduous teeth; IDPSCs, immature dental pulp stem cells; ES, embryonic stem; DPESCs, dental pulp epithelial stem–like cells.
Studies have revealed that IDPSCs is a multicellular stem cell population expressing neural crest/mesenchymal stem/progenitor cell markers, such as CD105 (Endoglin), CD73, CD29 (integrin β-1), CD44, and nestin (Table 1).4,5 Approximately 36% of IDPSCs expressed CD146, a marker of pericytes. Additionally, IDPSCs express markers of ES cells, such as Oct-4, Sox2, and Nanog. The expression of ES cell markers persists during several passages in multicellular IDPSC population and in single-cell-derived colonies. However, Oct-4 and Nanog expression is downregulated after induction of differentiation. 4 It seems that IDPSCs did not express markers of committed cells (Fig. 2B). IDPSCs showed high cloning capacity, which varies from 70% to 98% between cell populations isolated from different patients. IDPSC single-cell-derived colonies were almost of identical morphology, presented similar gene expression pattern, and differentiation properties similar to multicellular IDPSC population. The cloning capacity of IDPSCs was shown to be maintained after cryopreservation. 5
More recent studies described DTSC populations that shared similar properties with both SHED and/or IDPSCs (Table 1). Thus, Huang et al.11,12 used enzymatic digestion for DTSC isolation and used ES cell basal culture medium supplemented with 20% FBS (Hyclone) for cultivation. The resulting cell population (DTSC-1, here) shared similar phenotypic characteristics with IDPSCs (Table 1). These cells showed high cloning efficiency (∼83%) forming robust colonies. Nam and Lee 39 isolated two distinct populations—dental pulp epithelial stem–like cells and DTSC-2 (here)—using enzymatic digestion and two different cell culture medium—basal culture medium for keratinocytes and DMEM-low glucose supplemented with 10% FBS (Hyclone) (Table 1). Figure 2 demonstrates that all DTSC-isolated populations showed complete or partial overlap in the expression pattern of analyzed markers with SHED and IDPSCs. Such overlapping in the gene expression pattern was also observed between different cell colonies derived from the same ES cell line.40,41 These data show that all DTSC populations are composed of stem cells of ectomesenchymal origin in different proportions.
Telomere Length
Cell division and DNA replication result on telomere shortening in most adult cells. This shortening is a proposed mechanism of cell aging and may have important consequences for cell therapy. In actively proliferating stem cells, long telomeres (10–20 kb) are observed, whereas in somatic cells telomere length is about 5–15 kb. The study of telomere length variation during in vitro cultivation of rhesus monkey DTSCs demonstrated that telomeres were not significantly shortened in early (P5) or late (P17) passages. 11
Developmental Potential of DTSCs
A chimera is an organism composed of cells from two or more individuals from the same or different species. Production of preterm chimaeras is a method currently used to analyze the developmental potential of human stem cells.42,43 The developmental potential of human IDPSCs was tested in mouse/IDPSC preterm chimeras. It has been shown that five to eight of IDPSCs were able to survive, proliferate, and demonstrate significant contribution into inner cell mass of mouse blastocysts, which achieved the hatching stage (blastocysts hatched out of the zona pellucida) following 48 h of in vitro culture (Fig. 3A, B). After transferring these blastocysts into foster mice, they developed into preterm chimera embryos. The analysis of IDPSC distribution in different tissues in these chimeras demonstrated that the human cells engrafted into tissues from the three embryonic germ layers (Fig. 3C, D). The evaluation of their functional characteristics showed that IDPSC-derived epithelial cells express human nuclear protein and cytokeratin within mouse epithelial tissues. In mouse muscles, they formed hybrid human/mouse myotubes, which expressed human myosin (Fig. 3E). These data demonstrate that IDPSCs are biologically compatible with mouse fetal environment and display functional characteristics in the context of the tissues in which they reside. 44

Developmental potential of IDPSCs: homing and differentiation.
In Vitro Differentiation of DTSCs
Two types of stem cell differentiation in vitro were distinguished: spontaneous and induced. Induced differentiation maybe caused by chemical inducers, for example, retinoic acid, different growth factors, or by other biologically active molecules. Spontaneous differentiation of stem cells is triggered by cell confluence and/or by withdrawal of growth factors or substrates. For reference, ES cells are known to undergo spontaneous differentiation following withdrawal of mouse embryonic fibroblast (used as a feeder layer), or factors (leukemia inhibitory factor), which maintain their undifferentiated state. 35
Spontaneous differentiation of IDPSCs, similar to ES cells, was observed through manipulation of the cell seeding density and/or time of their transfer into chemically defined medium. IDPSCs cultivated at low density in chemically defined medium produced robust skeletal muscle differentiation, while smooth muscle cell differentiation was observed by cells seeding these under the same culture condition but at high density. 4
Induced osteogenic and adipogenic differentiation in SHED, IDPSCs, and DTSC-2 was achieved using specialized induction culture medium. IDPSCs were also able to undergo induced differentiation into skeletal muscle.1,2,45,46 Neural differentiation of SHED and IDPSCs was induced using neurobasal culture medium supplemented with B27. Both SHED and IDPSCs were able to form floating neurosphere-like clusters (NSLCs) in this medium. SHED grown in monolayer differentiated into neuron-like cells that expressed early and late neuronal markers (Table 1). 2 IDPSC-derived NSLCs adhered onto plastic and formed rosette-like structures producing outgrowth of neuron-like cells, demonstrating a process of neuronal differentiation similar to that observed with ES cells and with progenitors of central neural system. Following neuronal differentiation, IDPSCs were observed to form sandwich-like glial/neuronal structures mimicking primary culture of neuronal cells. 4
Recently, a capacity of DTSCs isolated from intact deciduous molars to differentiate into pancreatic cell lineage resembling islet-like cell aggregates (ICAs) has been demonstrated. Pancreatic cell lineage commitment was confirmed within DTSC-derived islets by dithiozone-positive staining and by the expression of proinsulin C-peptide, Pdx-1 (pancreatic duodenal homeobox–1), Pax4 and Pax6 (paired domain transcription factors), Ngn3 (neurogenin 3), and Isl-1 (Insulin gene enhancer protein) proteins. Furthermore, at the day 10, ICAs were able to produce insulin and C-peptide in a glucose-dependent manner confirming their functional state. 47
In Vivo Differentiation of Individual DTSCs
The differentiation potential of individual cells can be traced by clonal culture. Single-cell-derived colonies were obtained from SHED and IDPSCs. One-fourth of SHED clones (3 of 12) were shown to generate ectopic dentin-like tissue, when cultured on the hydroxyapatite/tricalcium phosphate ceramic powder carrier. This dentin was equivalent to that generated by multicolony-derived SHED. To assess in vivo bone formation, single-cell-derived SHED colonies were injected into immunocompromised mice. Variation in amount of bone production was observed between different SHED-derived colonies: approximately 40% of them induced formation of a significant amount of new bone in mice. Some of these colonies were also able to induce recipient murine cells to differentiate into bone-forming cells. 48 Single-cell-derived IDPSC colonies were transplanted intraperitoneally in immunocompromised mice. These colonies showed high migration ability and robust heterotopic grafting in all analyzed tissues, preferentially in liver, spleen, kidney, and bone marrow. They formed cell clusters, which exhibited morphological features of the recipient tissue.4,5
DTSCs in Dental Tissue Regeneration
Due to their origin, DTSCs could be expected to be a good candidate for dental tissue regeneration. To demonstrate this capacity, SHED were seeded in biodegradable scaffolds prepared within human tooth slices and implanted subcutaneously into immunodeficient mice. Differentiation of multicellular SHED population into functional odontoblastos (which expressed dentin sialoprotein and generated tubular dentin) was observed. In the same study, SHED differentiation into endothelial-like cells was shown; using SHED which had been stably transduced with LacZ. Positive B-galactosidase staining in the cells lining the walls of blood vessels within the tooth slice/scaffolds was found close to nonstained (host) blood vessels. This investigation confirmed that SHED has the capacity to generate DP-like tissue in vivo, whose architecture and cellularity resemble that of the natural tooth.48,49 This finding suggests the potential use of DTSCs for DP tissue reconstruction and biologically relevant tooth formation. 13
Osteogenic Potential of DTSCs
Bone formation by SHED was further observed using porcine cells. The efficacy of porcine autologous SHED in regenerating ortofacial bone defects was recently tested in miniature pig, where the critical-size bone defects were produced in the mandible. The cells were seeded onto a β-tricalcium phosphate scaffold and cultured for 7 days before transplantation into the defect. Six months after transplantation the porcine SHED were able to regenerate bone and to repair critical-size mandibular defect. 50
Human IDPSCs were also used to reconstruct critical-sized cranial bone defects in rats. Two symmetric full-thickness cranial defects in each parietal region were surgically completed. The cells were seeded onto collagen membrane (CM) at high density one day before transplantation. Two months after IDPSC transplantation, mature and well-formed bone was observed on the side of CM with IDPSCs, while in control side, less-mature tissue was detected. The IDPSC transplantation was performed without immunosuppression, and graft rejection has not been observed. 46
Bone tissue recovery in an ovine model with induced osteonecrosis of the femoral head was evaluated following transplantation of human IDPSCs and ovine BM-MSCs. Both cell types were stably transduced with LacZ in order to visualize the cell engraftment. After injection into femoral head, human IDPSCs and ovine BM-MSCs were able to proliferate within injured site and provided bone tissue fabrication. The histological analysis and B-galactosidase staining suggested that bone regeneration was more efficient (produced more mature bone) when IDPSCs were used. 51
DTSCs in Ocular Surface Reconstruction
It is known that DP and eye tissues originate from neural crest cells. This suggests that stem cells isolated from both organs could possibly form both of these tissues. Our group demonstrated that, indeed, IDPSCs shared similar properties with limbal stem cells (LSCs). IDPSCs express a set of specific LSC markers, such as integrin β-1 (CD29), vimentin, Delta N-p63 (tumor expressing protein p63), ABCG2 (ATP-binding cassette, subfamily G, member 2), Connexin 43 (gap junction protein), and K12 (cytokeratin-12). 52 In vivo, IDPSCs similar to LSCs have the capacity to reconstruct the entire corneal epithelium in total LSC deficiency in a rabbit model of chemically injured corneal surface. This study demonstrated that in vitro–fabricated IDPSC sheet, which was transplanted into chemically injured rabbit eye (xenotransplantation). The cells were able to survive, proliferate, and differentiate within rabbit eye. A new corneal surface was composed of IDPSCs, which expressed specific human proteins of the tissue in an appropriate and functional manner. 53 Therefore, IDPSCs formed a corneal epithelium that is equivalent to the natural cornea.
DTSCs in an Animal Model of Muscular Dystrophy
Golden retriever muscular dystrophy (GRMD) dogs are an animal model for preclinical therapeutic studies of human duchene muscular dystrophy. The repetitive injections of allogeneic canine mesoangioblasts performed under immunosuppressive protocol led to reduction of the disease severity by restoring canine dystrophin expression in GRMD dogs. 54 Human IDPSCs, which exhibited robust muscle differentiation in vitro, were also tested in GRMD dog's model, in order to evaluate safety, efficiency, and capacity to restore dystrophin expression. 55 IDPSCs were transplanted by local or systemic injection comparing the efficiency of unique and consecutive early transplantation. Local injection of the cells showed localization in connective tissue (Fig. 3F), while after systemic injection, the cells were localized in dog muscles (Fig. 3G) and/or distributed among other tissues. In muscle tissue, IDPSCs contribute to the formation of human/dog chimeric muscle fibers. Although significant engraftment of cells in GRMD dog muscles was observed, human dystrophin expression was limited to a few muscle fibers (Fig. 3H). Such difference between two studies can be explained by the cell type (allogeneic mesoangioblasts or IDPSCs) and/or protocol (with or without immunosuppression) used in these studies. The immunosuppressive, immunomodulatory, and antiinflammatory properties of MSCs as well as DPSCs are well documented.8,56–58 Therefore, nine consecutive applications of IDPSCs without immunosuppression did not induce an immunological response in dogs. 55 It is worth mention that GRMD dog, which received nine applications of IDPSCs, is currently alive and does not showed any type of tumor formation after 4 years.
DTSCs in Lupus Erythematous–Associated Disorder
Immunomodulatory properties of SHED were studied in mice model of (MRL/lpr) Inbred lymphoproli-feration strain with systemic lupus erythematous (SLE)—associated disorder. SLE is an immune disease, which provokes the destruction of targeted organ tissues and the accumulation of autoreactive lymphocytes and immune complexes. The disease induces multifaceted immune modulation, including both deficiency and hyperactivity of the immune system. Immunomodulatory properties of these cells were studied in vitro, which showed significant inhibition of T helper 17 (Th 17) cells. After transplantation in MRL/lpr mice, the SHED exhibited a high efficiency in reversing SLE. Transplantation of SHED in MRL/lpr mice contributed to the reconstruction of trabecular bone and appeared to inhibit osteoclast activity. It appeared that SHED use provided an enhanced therapeutic effect when compared with BM-MSCs. The upregulated level of the ratio of regulatory T cells (Tregs) and Th 17 cells suggests that SHED can provide a treatment for immune disorders via improved immunomodulatory properties. 59
DTSCs in Neuronal Injury
Neuronal differentiation of SHED multicellular population was studied after their injection into the dentate gyrus of the mice hippocampus. The data demonstrated that SHED were able to survive for more than 10 days in mice hippocampus and to express NFM, suggesting their appropriate differentiation in the homing site. 29
With the aim to improve spinal cord regeneration after trauma, IDPSCs have been used in a mouse model of compressive spinal cord injury. IDPSCs cultivated in vitro express mRNA transcripts of several neurotrophic factors, such as nerve growth factor (NGF), brain-derived neurotrophic factor, glial-derived neurotrophic factor, NT-3, and NT-4/5 (neurotrophic factors). Young adult female C57/Bl6 mice were subjected to laminectomy at T9 and compression of the spinal cord with a vascular clip, for 1 min. The IDPSCs were introduced at 7 days or 28 days after this lesion. Animals that received IDPSCs showed better white matter preservation than the group treated with only culture medium. Higher values of human trophic factors' expression and better tissue organization were observed with many axons showing myelination by either Schwann cells or oligodendrocytes. The presence of some healthy-appearing intact neurons with synapse contacts on their cell bodies has also been observed. IDPSCs were able to express the astrocyte marker GFAP and the Schwann cell marker S-100 after homing in recipient tissue. The functional analysis using global mobility test that evaluates the distance that the animal is able to walk in an open field and the speed that it reaches after 1 min of observation also showed locomotor improvement in IDPSC-injected mice but not in control animals. Additionally, this study suggests that IDPSCs can be applied in spinal cord injury treatments at 7 days or 28 days after the lesion. 60
iPS Cells Derived from DTSCs
iPS cells have been considered as an alternative to human ES cells in order to avoid the ethical problems of embryo destruction and to produce patient-matched pluripotent cells. The iPS cells can be generated by reprogramming of terminally differentiated cells or stem/precursor cells by means of overexpression of specific transcription factors, such as Oct3/4, Nanog, Sox-2, c-Myc, and Klf-4, which are responsible for pluripotency. 61 Such reprogrammed human cells are comparable with human ES cells, especially in respect to their differentiation capacity. The efficiency of reprogramming varies significantly between different starting cell types. It is lower in terminally differentiated cells, such as fibroblasts, whereas less-differentiated or undifferentiated adult cells undergo reprogramming process more rapidly and efficiently. 62 Two recent studies demonstrated derivation of iPS cells from SHED cells and IDPSCs (Fig. 3I).63,64 These cells present significant difference in protocol and efficiency of reprogramming. SHED required several attempts and two rounds of viral transduction for reprogramming, while IDPSCs demonstrated formation of small colonies, which express pluripotent stem cell markers at 5 days after transduction. After 11 days the first IDPSC-derived iPS cell colonies can be harvested. The SHED-derived iPS cells need mouse embryonic fibroblast (MEF) feeder layers for iPS cell isolation and growth. IDPSCs can be easily reprogrammed into iPS cells in the absence of MEF. SHED-derived iPS cells also require a longer time for formation of teratomas 63 in comparison with IDPSCs. 64 Both cell types, SHED and IDPSCs, demonstrated significantly higher efficiency of reprogramming than in fibroblasts. Establishment of iPS cells from DTSCs could be useful for the studies of pediatric diseases and disorders.
DTSCs Versus DPSCs
DPs from deciduous and permanent teeth differ both anatomically and functionally and hence could differentially determine stem cell fate. The gene expression profile determines tissue and stem cell capacity to differentiate toward a specific lineage. This analysis also allows a composition of the properties of different stem cell populations isolated from the same or different sources. DNA microarray analysis revealed 4386 genes with a differential expression between DPSCs and DTSCs (SHED) by 2.0-fold or more. In particular, higher expression in DTSCs was observed for the genes related to pluripotency (OCT4, SOX2, NANOG, and REX-1), cell proliferation, and extracellular matrix, including several cytokines such as fibroblast growth factor and tumor growth factor β. The most prominent upregulation was observed in expression of collagens (Col I, III, VII, and XIII) and proteoglycans (glypican and versican). Both of these studies infer that DTSCs retained, whereas DPSCs lost, their plasticity through passaging. Taken together these results provide an additional evidence of the divergence in stem cell properties between DTSCs (deciduous teeth) and DPSCs (adult teeth) and indicated that DTSCs could be useful for cell-based regeneration therapy not only for oral disease but also for systemic diseases.65,66
DTSC Banking
Several technical and economic issues are relevant with respect to DTSC banking. The loss of baby teeth occurs naturally and they can be removed with minimal discomfort to the patient during a routine visit to the dentist. DP can be cryopreserved and the cells can be obtained later after thawing. Therefore, frozen DP does not require cell culture conditions until the donor needs these cells for a clinical treatment. Additionally, several DPs from the same individual can be cryopreserved.67–72
Recent reports demonstrated that DTSC isolation seems to be related with physiological process of the root resorption. The authors failed to isolate DTSCs from the DP of patients, which did not show any visible root resorption. Only DP from the teeth, which presented advanced levels of root resorption, was able to generate DTSCs. 73 This data partially confirmed our previous unpublished observations. Our group succeeded to isolate DPSCs from DP without visible root resorption. In this case, outgrowth of the cells from DP is delayed at least for one week when compared with DP at advanced level of root resorption. Following further in vitro cultivation, these DPSCs became indistinguishable with respect to proliferation rate and immunophenotype from that of DP without visible root resorption. 4 From a practical standpoint, this result supports DTSC isolation during normal eruption, thus precluding earlier dental intervention.
Considering that currently BM-MSC isolation occurs from volunteers, who are genetically distant, the advantage of DTSCs is that they can possibly be used by the parents, who share 50%, or by grandparents, who share 25%, of the genetic background with the donor, also minimizing the distribution of unknown genetic elements present in the human population. It is important to note that families that have a history of genetic, cancer, or other types of diseases should avoid the use of stem cells from closely related family members. Aging of stem cells in organism should also be taken into consideration. A striking decrease of BM-MSCs per nucleated marrow cell was previously observed: 10-fold decrease from birth to the teenage years and another 10-fold decrease from the teenage years to old age. 34 In contrast, DPs are an abundant source of young stem cells. DTSCs can benefit the oldest members of the family. Once stored, DPs can be constantly renewed within the family avoiding long-term cryopreservation of DP tissue or DTSCs.
Conclusions
Isolation of stem cells from human adult teeth has been reported for the first time in 2000, attracting the attention of researches to this promising source.5,6,13,69,74 Even less time has passed from the moment of the discovery of DTSCs.1,2 In 8 years, substantial progress has been achieved in investigations of these cells and it has been shown that (1) the explant culture of DP led to harvesting of a relatively pure cell population of DTSCs; (2) DTSCs express pluripotent stem cell markers (Oct3/4, Nanog, Rex-1, and Sox2); (3) based on gene expression profile different populations of DTSCs can be isolated: MSC-like population, epithelial stem cell–like population, and mixed populations of ectomesenchymal stem cells in different proportions; (4) DTSCs are multipotent cells with high differentiation potential that are able to contribute to all embryonic germ lineage formation in pretermed chimeras; (5) in vivo, in animal models, DTSCs present tissue regeneration capacity in response to the cellular milieu; (6) immune rejection and teratomas formation was not observed after DTSCs; therefore, they can be used in parental allogeneic stem cell transplantation; (7) iPS cells can be easily obtained from DTSCs; (8) DTSCs are more immature cells in comparison with DPSCs; (9) DTSCs are almost unlimited source of young stem cells with easy access.
Footnotes
Acknowledgments
The authors are very grateful to Prof. Dr. Alexandre Kerkis (CELLTROVET Company Brazil) coauthor of all publications, for his intellectual contribution, and to all technicians and students of the Laboratory of Genetics for their contribution to this project. We thank David A. Carrino for his editorial assistance. We thank FAPESP, CNPq, NIH, and the Virginia of David Baldwin Research Found for financial support.
Disclosure Statement
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.