
Abstract
Cardiovascular tissues exhibit architecturally complex extracellular matrices, of which the elastic matrix forms a major component. The elastic matrix critically maintains native structural configurations of vascular tissues, determines their ability to recoil after stretch, and regulates cell signaling pathways involved in morphogenesis, injury response, and inflammation via biomechanical transduction. The ability to tissue engineer vascular replacements that incorporate elastic matrix superstructures unique to cardiac and vascular tissues is thus important to maintaining vascular homeostasis. However, the vascular elastic matrix is particularly difficult to tissue engineer due to the inherently poor ability of adult vascular cells to synthesize elastin precursors and organize them into mature structures in a manner that replicates the biocomplexity of elastic matrix assembly during development. This review discusses current tissue engineering materials (e.g., growth factors and scaffolds) and methods (e.g., dynamic stretch and contact guidance) used to promote cellular synthesis and assembly of elastic matrix superstructures, and the limitations of these approaches when applied to smooth muscle cells, the primary elastin-generating cell type in vascular tissues. The potential application of these methods for in situ regeneration of disrupted elastic matrix at sites of proteolytic vascular disease (e.g., abdominal aortic aneurysms) is also discussed. Finally, the review describes the potential utility of alternative cell types to elastic tissue engineering and regenerative matrix repair. Future progress in the field is contingent on developing a thorough understanding of developmental elastogenesis and then mimicking the spatiotemporal changes in the cellular microenvironment that occur during that phase. This will enable us to tissue engineer clinically applicable elastic vascular tissue replacements and to develop elastogenic therapies to restore homeostasis in de-elasticized vessels.
Introduction
The quantity and quality of elastogenesis that can be achieved via different tissue engineering strategies are determined by concurrent impact of culture parameters on (1) the expression and synthesis of various proteins and subunits that constitute elastic fibers, and (2) the balance of cellular matrix metalloproteases (MMPs) and tissue inhibitors of matrix proteases (TIMPs) that directly influence integrity of generated matrix at three distinct stages of elastic matrix synthesis: (i) mRNA expression of the proteins, (ii) their translation into protein synthesis, and (iii) extracellular transport of precursor molecules and assembly of matrix structures. The proteins that constitute the elastic matrix primarily include tropoelastin, fibrillin, and fibulin, while crosslinking is mediated by the enzyme lysyl oxidase (LOX). The quality of the synthesized matrix is evaluated relative to that found in healthy vessels in vivo based on the structure, orientation, dimensions, and continuity of elastic fibers, and the extent of desmosine crosslinking. This article focuses on a critical analysis of the different strategies that have been attempted to tissue engineer vascular conduits containing a robust elastic matrix mimicking that in native blood vessels, which these conduits are expected to replace.
This review article starts with a description of the current challenges that exist in the field of elastic matrix engineering, specifically for vascular applications. Next, specific targets and processes that impact elastogenesis are introduced with a brief description of the steps involved in elastic fiber assembly during fetal development. This information is centric to understanding how specific culture parameters and techniques impact elastogenesis. The parameters described in this review include characteristics of the native ECM microenvironment as well as cues from tissue engineering approaches (e.g., growth factors, mechanical stimuli, scaffold chemistry, and the use of alternative cell sources). Finally, the article concludes with a review of current progress toward achieving robust elastogenesis within tissue engineering vascular grafts aimed at clinical use.
Biocomplexity of Elastic Matrix Assembly
The development of elastogenic technologies requires familiarity with the physiologic process of cellular elastin synthesis and matrix assembly. This process involves several steps of coordinated intracellular and extracellular activities directed by vascular smooth muscle cells (VSMCs) and has been described in more detail elsewhere.7,8 Broadly, the elastin protein is secreted by VSMCs as the tropoelastin precursor molecule into the extracellular space. There, the tropoelastin precursors bind with the elastin binding protein (EBP), a chaperone that protects them from degradation, via their hydrophobic amino acid sequence VGVAPG. 7 The EBP is part of a larger cell-surface elastin-binding complex that contains two membrane proteins of similar sizes (55 and 61 kDa) that link the extracellular space to the cytoskeleton. The EBP simultaneously binds galactosugars, at its galactolectin binding site, and the cell membrane. Linear arrays of microfibrillar glycoproteins in the extracellular space present galactosugars that engage the galactolectin binding site, reducing binding affinity of the EBP for tropoelastin and the cell membrane and resulting in the release of bound tropoelastin and the dissociation of the EBP from the cell membrane. The tropoelastin is thus able to congregate on the microfibrillar scaffold as amorphous elastin and then undergo crosslinking. 9 The microfibrils are composed of several different glycoproteins, most prominently fibrillin, and serve to maintain the integrity of elastic fibers and enable VSMCs to interact with these structures. Crosslinking of elastin is initiated by oxidation of lysine amino acid side chains on the elastin molecules by a Cu2+-dependent enzyme, LOX. 10 The microfibril scaffold provides the necessary spatial coordination/alignment for the congregation of tropoelastin and its crosslinking into elongated fibers. The mature fibers, typically 300 nm to more than 1 μm in diameter, contain a central core of electron-dense crosslinked, amorphous elastin, surrounded by microfibrils.11,12 Provided that alignment and crosslinking progress appropriately, adjacent elastin molecules within these fibers are linked by desmosine and isodesmosine moieties, which impact the ability to stretch and recoil as a single unit. Finally, elastic fibers interconnect to organize into primarily three structural forms, namely, fibers, discrete fiber networks, and denser, fibrous membranes or sheets. 13 Due to such crosslinking and organization into higher order structures, elastic fibers exhibit mechanics, particularly elasticity, which is significantly different and unique compared to other structural ECM components found in vascular tissues. For example, collagen fibers exhibit a more viscoelastic response, resulting in interfibrillar slippage that must be repaired by the surrounding cells. 14 The long-range deformability, passive recoil, and low hysteresis of elastic fibers are all characteristic of the elasticity. The individual contributions of collagen and elastic fibers to vessel mechanics have also been quantified from the different regions of the stress–strain curve. Armentano et al. have found an elastic modulus of 0.49 MPa for elastic fibers and 131 MPa for collagen in the thoracic aorta of dogs. 15
The repair of existing protease-degraded elastic matrix in diseased tissue can proceed through a similar process. Although the fibrillin scaffold is already present, new tropoelastin is deposited and crosslinked at the site of the existing matrix. Another process proposed by Berk et al. is that some degraded desmosine and isodesmosine crosslinks can be reformed or salvaged, as determined through a radioactive assay. 16 The exact mechanism for this reaction is not currently known. In addition, the extent of remodeling that can occur has been shown to depend on the degree of elastic matrix degradation. 17 Finally, since matrix provides a reservoir for growth factors, the presence of growth factors in pre-existing matrix is the other notable difference with diseased tissue.
Challenges to In Vitro Engineering and In Vivo Regenerative Repair of Elastic Matrix
Considering the complexity of elastic matrix assembly, as described above, it is easy to appreciate the challenges that undermine biomimetic generation of elastic matrix structures by healthy patient-derived VSMCs or even less elastogenic, diseased VSMCs. Perhaps the biggest impediments to engineering elastic tissues within controlled cellular microenvironments in vitro are the progressive destabilization of tropoelastin mRNA, 18 its diminished translation in most postneonatal cell types, 19 with exceptions such as SMCs of the bladder 20 and female reproductive tissues, 21 and inefficient recruitment and crosslinking of tropoelastin into an elastic matrix. 22 There is currently a lack of knowledge of materials (e.g., tissue engineering scaffolds) and methods to promote assembly of precursors into mature elastic fibers 23 and further organize these fibers into structural and functional mimics of native vascular elastic matrix superstructures. In some diseased conditions (e.g., atherosclerosis) the cellular deposition of amorphous amyloid elastic fibers has been reported 24 ; however, these disorganized pseudo-fibers actually contribute to the pathology and are easily degraded by proteolytic enzymes due to their limited crosslinking. This reinforces the importance of crosslinking matrix elastin deposited by cells within tissue engineering constructs. Even more complex is the prospect of affecting regenerative repair of elastic matrix in situ within proteolytically degraded matrix microenvironments (e.g., aortic aneurysms) not associated with genetic defects in elastic matrix assembly proteins or machinery. Here, there are additional deterrents to elastic matrix synthesis, which include (1) switch of a subset of SMCs to a diseased phenotype characterized by poor elastogenicity compared to even healthy SMCs, 25 (2) diminished production of crosslinking enzymes and/or their impaired action, and (3) enhanced local production and activity of elastolytic enzymes by both recruited inflammatory cells and SMCs. 26 Together, these events reduce net accumulation of new elastin deposits and disrupt new and pre-existing elastic matrix structures, which would serve as nucleation sites for growth of new elastic fibers. 13
ECM Microenvironment Influences Elastogenic Outcomes
It is clear from existing literature that the composition of the extracellular microenvironment can critically influence cell phenotype and behavior, with potentially adverse repercussions to elastic matrix synthesis. SMCs, which typically exhibit a contractile phenotype within healthy, intact vascular tissues, switch to a more synthetic (i.e., proliferative and ECM-generating) phenotype when isolated and cultured in vitro.27,28 This phenotypic switch occurs when the SMCs are cultured on either tissue culture polystyrene or within scaffolds created from one of the many natural (e.g., fibrin 29 ) or synthetic (e.g., polyglycolic acid [PGA] 30 ) polymers. While VSMCs in contact with the above scaffolds have been found to increase their production of tropoelastin precursors, 31 both at expression of elastin mRNA and its translation into protein, they abundantly and preferentially synthesize and deposit collagen, beginning at a fairly early stage in culture. Such collagen fiber deposition is favorable from the standpoint of replicating the 3D collagenous matrix within vascular tissues. However, there is also significant evidence that a collagenous microenvironment promotes a more quiescent, contractile phenotype among SMCs, compared to other scaffolding materials such as PGA and fibrin, and limits cellular synthesis of elastin precursors and assembly of elastic matrix structures.30,32–34 The same issue is highlighted in diseased vascular tissues where accumulation of a disorganized collagen matrix, as part of a fibrotic injury response, can inhibit inherent or induced elastic matrix deposition by cells. Recently, Mitts et al. 35 showed the steroid aldosterone to induce collagen synthesis by human dermal fibroblasts through activation of mineralocorticoid receptors (MRs) and elastogenesis via MR-independent mechanisms. Concurrent exposure of the cells to an MR inhibitor, spironolactone, was found to inhibit collagen synthesis while improving elastogenesis. A similar strategy applied at the onset of in vitro culture of VSMCs may prove useful to achieve robust elastic matrix deposition over collagen matrix generation.
Elevated production of galactosugar-containing glycosaminoglycans (GAGs) such as chondrotin sulfate (CS), and CS-presenting proteoglycans (PGs; e.g., versican) has been shown to inhibit elastic fiber formation. 7 These GAGs are typically produced by non-neonatal VSMCs in culture and in diseased vascular microenvironments (e.g., restenotic lesions). Conversely, neonatal VSMCs produce little or no versican but abundantly deposit fibrous elastic matrix. 36 The inverse relationship suggests that the presence or production of versican interferes with elastic fiber assembly. Studies by Hinek et al. 7 have shown that the CS side chains in the versican molecule engage the galactolectin domain of the EBP on the cell surface, resulting in diminished tropoelastin affinity for the complex, displacement of the EBP from the cell surface, and impairment of elastic fiber assembly (Fig. 1). In addition, a series of studies by Joddar and Ramamurthi37,38 indicated that long-chain forms of the nonsulfated GAG hyaluronan (HA; MW > 104 Da) encourage tropoelastin mRNA expression, protein synthesis, recruitment, and crosslinking, but interfere with higher level elastic matrix assembly. In fact, upregulated production and accumulation of long-chain HA, such as within restenotic lesions, 36 appear to correlate with enhanced deposition of elastic matrix. The enhanced accumulation of elastin deposits in the matrix may be due to high anionicity of GAGs, which may serve to coacervate soluble tropoelastin and facilitate LOX-mediated crosslinking into an insoluble matrix, as certain studies have implied. 39 Despite these benefits, the presence of long-chain HA at the cell surface also encourages binding of versican via its HA binding motif, followed by engagement of the EBP by its CS chains as described above. As an alternative, recent work by Merrilees et al. 40 has shown that V3, a splice variant of versican lacking the β-galactosidase domain that contains CS, competes with versican for binding to HA, thereby preventing CS-mediated interference with elastic fiber assembly. Likewise, culture of rat SMCs with exogenous, soluble, 37 or substrate-immobilized 41 HA oligomers (4 and 6 mers) was found to both induce multi-fold increases in elastic matrix yield as well as significantly improve fiber formation. This may potentially occur through competitive inhibition of the binding of cell-synthesized long-chain HA to cell surface CD44 receptors and subsequent deterrence of versican binding to the long-chain HA.

Inhibition of elastic fiber formation by versican and chondrotin sulfate (CS). Adult smooth muscle cells (SMCs), but not neonatal SMCs, in culture and in diseased microenvironments, produce abundant amounts of the CS-bearing proteoglycan, versican, which binds to long-chain hyaluronan (HA) in the extraellular space
Elastogenic Stimulation of Cells Using Growth Factors
Controlled growth factor delivery to cells continues to be a popular strategy to improve elastic matrix deposition within tissue-engineered constructs. Cells perceive growth factors from their environment through cell surface receptors, which trigger changes in intracellular signaling proteins and pathways when activated or inhibited. This signaling leads to physiological outcomes such as enhanced or inhibited elastic matrix synthesis. Prior studies have shown that elastin matrix synthesis is regulated by numerous biochemical factors that may be broadly classified as elastin synthesis inhibitors (e.g., ascorbic acid, 42 basic fibroblast growth factor 43 , cyclic AMP 44 , and monensin 45 ) and elastin synthesis stimulators (e.g., cyclic GMP, 44 fetal calf serum, 46 insulin-like growth factor 1 [IGF-1],47,48 and transforming growth factor β1 [TGF-β1] 49 ). The most prolifically studied factors to date for elastogenic stimulation of VSMCs are TGF-β and IGF-1, which have been shown to increase elastin synthesis both by upregulating cellular mRNA expression of tropoelastin and by correspondingly increasing protein translation and matrix deposition. Additionally, TGF-β1 is shown to improve matrix assembly by increasing Lox mRNA expression and enzyme activity, 50 and preventing proteolysis of existing matrix by downregulating Mmp 2 and 9 and upregulating Timp 1, 2, and 3.51,52 Kothapalli and Ramamurthi 53 and Kothapalli et al. 54 showed these elastogenic effects to be enhanced synergistically when TGF-β1 and IGF-1 were provided to healthy VSMCs, both rat and human, together with the HA oligomers (HA-o) discussed above. Together, these cues suppressed SMC proliferation, enhanced synthesis of tropoelastin and matrix elastin protein, and significantly improved the mass fraction of total elastin synthesized and organized into a matrix. This may occur through a combination of mechanisms, including observed increases in elastin mRNA, coacervation of tropoelastin precursors by HA for more efficient crosslinking, and enhancement of Lox mRNA expression, protein synthesis, and crosslinking activity. The effects of growth factors such as TGF-β have been shown to be concentration-dependent and biphasic, 55 with adverse events, such as matrix mineralization, triggered by excessively high doses. 56 Thus, careful optimization of factor doses on a per-cell basis is required. It is noteworthy that the elastogenic effects of aldosterone on human skin fibroblasts, as discussed earlier, have been shown to act via IGF-1 receptor signaling. 35 The steroid may also potentially be employed to enhance elastic matrix synthesis within engineered vascular constructs incorporating VSMCs. The elastogenic effects of ascorbic acid/ascorbate may also be dose dependant. Franzblau and co-workers57–60 have shown that withdrawal of ascorbic acid/ascorbate improves elastic matrix deposition and crosslinking. However, Kiere et al. 61 showed that engineered vascular constructs cultured under ascorbate-free conditions, following an initial period of culture with ascorbate, exhibit increased burst strength and elasticity. Finally, studies from the 1990s have also shown that retinoic acid supplements modestly enhance (i.e., two- to threefold increases) elastin gene expression, insoluble elastin deposition, and elastic fiber formation by VSMCs 62 and fibroblasts. 63
Increased bioavailability of elastogenic factors appears to also benefit regenerative repair of elastic matrix by SMCs of a diseased phenotype in matrix-disrupted vascular microenvironments in vivo. Transient paracrine secretion of TGF-β1 by endovascularly seeded SMCs in rat abdominal aortic aneurysms (AAAs) was found to increase matrix production, though mostly of collagen, by various cell types (e.g., SMCs and fibroblasts) in situ. LOX production, which is critical for collagen and elastin crosslinking and fiber organization, was also enhanced. Dai et al. 51 used gene therapy techniques to overexpress TGF-β1 within already-formed rat AAAs and qualitatively showed improved elastin stabilization and endoluminal reconstruction, as evidenced by the generation of a collagen- and elastic-fiber-rich intimal lamina. In the context of inducing regenerative elastic matrix repair in vivo, it is important to emphasize a critical need to deliver the correct elastogenic signals to the right place and at the ideal time. 64 This is necessary to evoke the desired outcome, particularly since spatial heterogeneity and temporal changes in cellularity, cellular phenotype, pre-existing ECM composition and architecture, and paracine/autocrine cell signaling within the disease tissue site can influence cell responses, including those pertinent to elastogenesis.
As important as increased elastin synthesis is the need to enhance yield of elastic matrix, which is necessary to ultimately generate sufficiently large elastic tissue constructs for human use. The LOX family of copper-dependent proteins, which include up to five close variants designated as LOX-like proteins that promote elastic fiber renewal, 10 play a critical role in tropoelastin crosslinking. Kagan and co-workers 65 showed that tropoelastin generated by recombinant DNA techniques can be crosslinked by exogenous LOX. Kothapalli and Ramamurthi 66 have also shown that exogenous LOX supplemented to rat aortic SMC cultures improves tropoelastin crosslinking significantly and enhances elastic matrix deposition beyond that achievable by endogenous LOX enzymes alone. Copper ions (Cu2+) enhance extracellular transport of endogenous LOX and functional activity of endogenous and exogenous LOX, by enabling electron transfer from oxygen to facilitate oxidative deamination of lysyl groups in elastin/collagen. 10 For this reason, Kothapalli and Ramamurthi 67 supplemented rat aortic SMC cultures with copper sulfate and found it to improve LOX synthesis and elastin crosslinking activity. Electrostatic interaction of Cu2+ with anionic, exogenous HA was found to localize these crosslinking effects. A critical problem that was, however, encountered with delivery of soluble copper salts to cells was cytotoxicity at higher doses (i.e., 0.1 M in the above study). 68 A follow-up study by the same investigators showed that more controlled, gradual release of Cu2+ ions from copper nanoparticles (CuNP) does not induce cytotoxicity and also benefits elastin crosslinking activity in the vicinity of the cells by settling of the CuNP and interaction with endogenous or exogenously supplemented GAGs (Fig. 2). 11
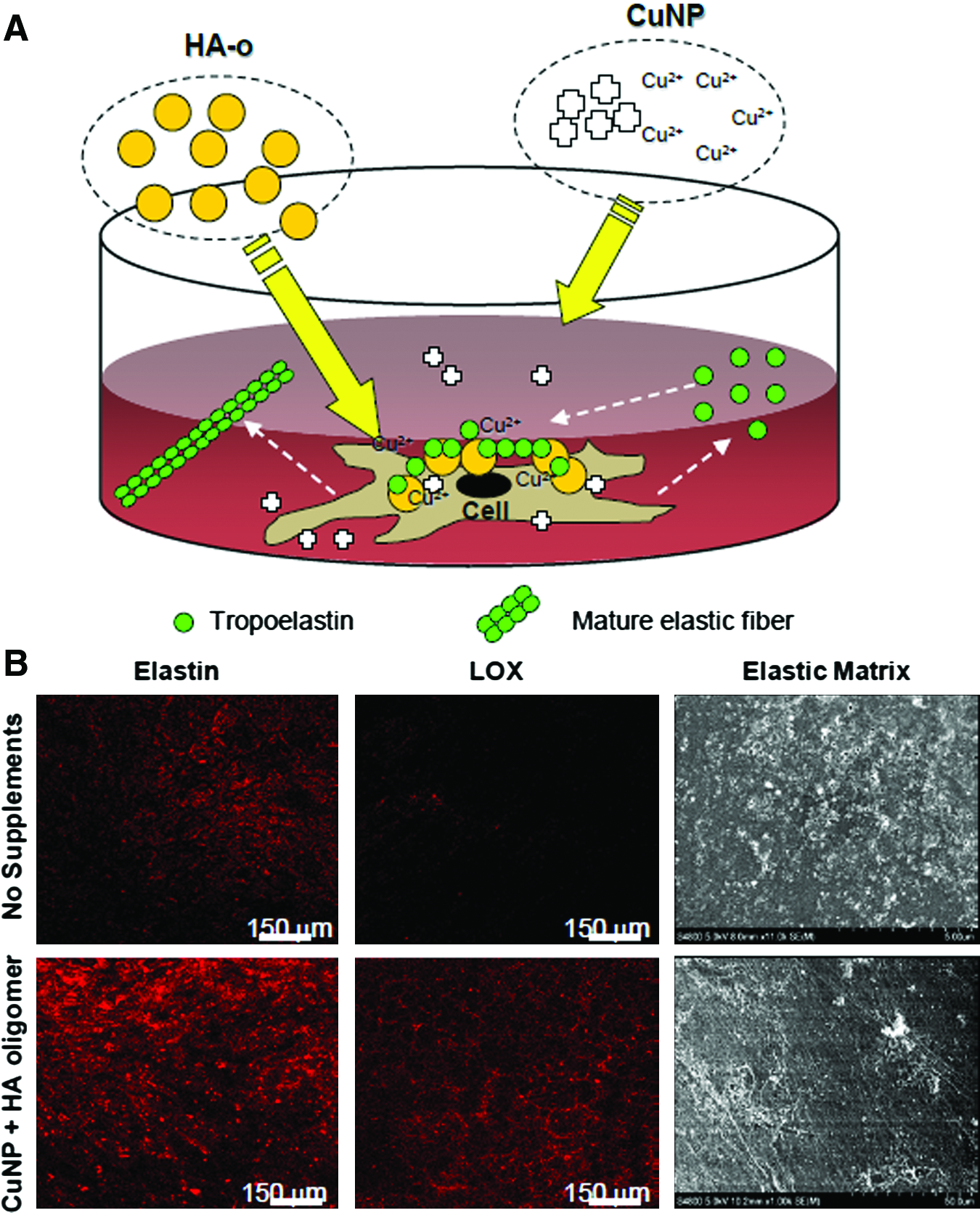
Hypothesized mechanism for improvement of tropoelastin crosslinking and fiber formation by copper nanoparticles (CuNP) and HA
Scaffolding Characteristics Impact Elastogenic Outcomes
Cell phenotype and matrix synthesis are affected by the substrate in which they are grown upon or within. The physical and mechanical properties of a substrate depend on the type of macromolecule (i.e., natural or synthetic) and the processing techniques (e.g., phase inversion, salt leaching, and electrospinning). As previously mentioned, SMCs' microenvironment (e.g., biomaterial substrate) influence their phenotype, with the cells generally assuming a more synthetic, ECM-generating phenotype when isolated and cultured on most biomaterial substrates in vitro.27,28 However, the specific responses are dependent on the substrate properties, with both the chemical composition of cellular substrates and their physical attributes (e.g., mechanics and topography) known to influence cell behavior. Thus, careful selection of cellular scaffolds that facilitate elastogenesis, and yet appropriately match vascular tissue compliance and mechanics, is important. Table 1 highlights recent advancements in vascular tissue engineering approaches that have shown promise in generating elastic matrices using various scaffolding materials and other factors. Since engineering an elastic matrix has not been the primary focus of most tissue engineering groups so far, it is important to note that most studies have not comprehensively analyzed the quality and quantity of cell-generated elastic matrix and its components; in most cases, elastogenic assessment has been limited to reverse transcription–polymerase chain reaction of tropoelastin mRNA expression and qualitative histology.
SMCs, smooth muscle cells; RASMCs, rat aortic smooth muscle cells; PGA, polyglycolic acid; PHA, polyhydroxyalkaonate; PCL, poly(ɛ-caprolactone); SPC, smooth muscle progenitor; SOCs, smooth muscle outgrowth cells; TGF-β1, transforming growth factor β1; HA-o, HA oligomers; 2D, two-dimensional; 4-HB, 4-hydroxybutyrate; LOX, lysyl oxidase; EC, endothelial cells.
2D versus 3D substrates
The phenotype of SMCs is distinctly different when cultured on 2D surfaces compared to porous, 3D scaffolds, even when both substrates exhibit identical chemistries (Fig. 3). 85 Cells within 3D scaffolds by and large exist in microenvironments that provide spatial cues that more closely mimic native tissue. In 3D cultures, VSMCs tend to assume a more contractile, less proliferative,86,87 and poorly collagen generating phenotype, although some articles have reported overall cell numbers to be much greater within porous 3D scaffolds, potentially due to the greater surface area they afford for cell adherence and proliferation. 82 An inverse relationship has been observed between SMC proliferation and collagenous ECM synthesis on 2D surfaces and 3D matrices.28,86 In a recent study, Bashur and Ramamurthi 82 showed that rat aortic SMC proliferation on smooth, 2D films of poly(ɛ-caprolactone) (PCL) was far more limited than within 3D electrospun PCL fiber meshes, while collagen matrix deposition by the cells was higher. Of note, opposing trends in collagen and elastic matrix generation were observed in this study and in others.42,82,88 Despite increased elastic matrix deposition on the 3D electrospun PCL scaffolds described above, compared to 2D tissue culture plastic, no corresponding differences were noted in the mRNA expression of matrix proteins. This suggests that substrate chemistry, surface topography, and surface compliance (i.e., the ability to be deformed) can in certain cases influence post-transcriptional mechanisms of tropoelastin recruitment and crosslinking without impacting elastin gene expression itself. 82

Illustration of cellular cues presented by 3D scaffolds with
Scaffold compliance and dynamic stimulation
Cellular phenotype and elastic matrix deposition is also affected by the stiffness of the scaffolding substrate. This has been related to the ability of cell traction forces to deform the substrate, a mechanism that has been proposed by Crapo and Wang, 84 and others.89,90 Work by Engler et al. has shown that the differentiation of mesenchymal stem cells (MSCs) cultured on collagen-coated acrylamide gels was directed by the hydrogel stiffness. 91 Gels prepared with stiffnesses corresponding to different types of tissue (i.e., brain E=0.1–1 kPa, striated muscle E=8–17 kPa, and collagenous bone E=25–40 kPa) coaxed MSC differentiation into cell lineages typical of the parenchyma of those tissues. It is important to note that these moduli are at the surface, or what the cells would feel, rather than conventional tensile moduli. Since fully differentiated cells (e.g., VSMCs) also exert traction forces on their surfaces, it can be expected that substrate stiffness also affects the behavior and matrix production of these cells. It has been found that poly(ethylene glycol) hydrogels exhibiting higher tensile moduli (i.e., 424 kPa) promote SMC proliferation and co-localization of calponin, a calcium binding protein associated with a contractile SMCs, with the α-SMA cytoskeleton. 92 It has also been found that excessive/rapid shrinkage of scaffolding materials, as encountered with scaffolds based on PGA, may adversely impact elastin biosynthesis by VSMCs. 93 However, Crapo and Wang found that baboon SMCs cultured on porous, elastomeric poly(glycerol sebacate) scaffolds produced more insoluble matrix elastin than on the stiffer poly(lactic-co-glycolic acid) (PLGA) constructs. 84 These results indicate the importance of surface stiffness/compliance on SMCs, and also that an ideal compliance for VSMCs elastic matrix production still needs to be established.
The application of mechanical stimuli is another tool that applies a force to the cytoskeleton and employs mechano-transductive signaling to impact SMC matrix production. 87 In general, the goal is to replicate the stimuli found in native tissue, which for VSMCs is cyclic mechanical stretch. Kim et al. found that rat aortic SMCs exhibited increased collagen and elastin synthesis when stretched to 20% strains for 4 days (frequency of 0.05–0.25 Hz), but they also found that the results were also dependant on the matrix that the cells were attached to. 93 Li et al. found that the magnitude of strain impacts SMC matrix production. Specifically, they found that collagen production was higher for 10% strain compared to 5%, but no further increases were observed up to 20% strain. Cyclic strains are shown to induce matrix synthesis by SMCs in culture through a tyrosine-kinase dependent TGF-β1 pathway.94,95 It has also been established that excessive mechanical stimulation (e.g., high strains) can induce negative cellular responses, including apoptosis. 96 Wernig et al. established that SMC apoptosis acts through an integrin signaling pathway. 96 Together, these findings emphasize the importance of both an appropriate stretch regime and cellular interaction with other aspects of its microenvironment.
Scaffolding chemistry
There is evidence that SMC cultures within biological, 3D scaffolds based on naturally occurring ECM components are superior to those cultured within 3D synthetic scaffolds or on 2D monolayer cell cultures,97,98 since they better simulate the chemical and physical environment of vascular tissues. However, they do require some degree of crosslinking to prevent enzymatic degradation in vivo. Cells within these scaffolds are able to bind to ECM via integrins, 99 allowing interaction with cell adhesion molecules, growth factors, and mechanical stimuli similar to that within native vascular tissues. Moreover, the ECM is known to serve as a reservoir for growth factors that further affect cell behavior. Since collagen is a major matrix component of vascular tissues, collagen scaffolds have been extensively studied in the context of growing vascular replacements. A number of reports, including those by Weinberg and Bell, 32 who used bovine aortic SMCs; L'Hereaux et al., 33 who studied human umbilical cord vein SMCs; and Hirai et al., 100 who studied canine jugular SMCs, have demonstrated poor elastin synthesis within constructs containing mostly collagen types I or III. Long and Tranquillo 29 showed that gels containing fibrin alone, or collagen I and fibrin together, encourage robust production of native elastin-like fibers by neonatal SMCs. However, even these cell types, which are much more elastogenic than adult VSMCs, 18 produced little elastic matrix when seeded within collagen constructs. Ahmann et al. 101 later showed that fibrin degradation products generated by VSMC remodeling of fibrin gels directly stimulate increased elastin and collagen matrix synthesis. Xie et al. 75 showed that rat peripheral blood (PB) smooth muscle progenitor-derived smooth muscle outgrowth cells cultured on silk fibroin-modified poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) scaffolds were able to synthesize elastin protein. However, the deposition of an elastic matrix and the formation of elastic fibers were not investigated. Hydrogels based on GAGs have also been very useful to generate elastin-rich matrices. Ramamurthi and Vesely 76 showed that hydrogels containing crosslinked long-chain HA are too anionic and hydrophilic to even support cell adherence. However, surface irradiation of these materials with UV light, which generated shorter HA fragments, not only promoted adherence of neonatal and adult rat VSMCs, but also stimulated their deposition of a thick layer of fibrous and sheet-like elastin at the cell–gel interface. 38 As previously mentioned, Joddar and Ramamurthi found HA oligomers (4 and 6 mers) to stimulate elastin precursor synthesis, elastic matrix deposition, crosslinking, and fiber formation by adult rat VSMCs. 38 Higher molecular weight HA (i.e., MW > 104 Da) did not induce elastin precursor synthesis; however, it did appear to coacervate elastin precursors via opposite charge interactions, to enable more efficient crosslinking, and thus generate higher fractional yields of elastic matrix. 37 Therefore, including both these forms of HA into biomaterial scaffolds may be a useful strategy to enhance elastic matrix deposition by VSMCs. Ibrahim et al. 102 further showed that chemical derivatization and photocrosslinking of bioactive HA-o and long-chain HA improves their half-life and does not adversely impact their elastogenic effects on VSMCs. Since certain sized HA fragments can incite inflammation under specific circumstances, it must be emphasized that presentation of HA oligomers on and within scaffolding materials should be carefully controlled and cellular activation (in vitro) or inflammatory responses (in vivo) evaluated. Corroborating these findings, Zavan et al. 103 demonstrated the usefulness of vascular grafts based on HYAFF, a HA derivative, to guide the development of a well-functioning neoartery when implanted end-to-end within rat aorta, characterized by presence of layers of elastic fibers. Contingent on drastically improving their generally poor handling properties and their proclivity for enzymatic breakdown, or developing methods to present them on other mechanically stronger scaffolds, HA-based biomaterials may be adopted more widely for elastic matrix tissue engineering.
Synthetic polymer scaffolds, although lacking specific cell-binding sequences, are commonly used for tissue engineering and proffer several advantages over biological scaffolds. These advantages include the ability to mass-produce the scaffolds, to tune their biochemical and physical characteristics to achieve desired rates of degradation, to control the type and quantity of their degradation products, and to match the compliance, tensile strength, and size of constructs to the target tissue. The most frequently investigated synthetic, degradable polymers for CV applications have been polyesters such as PGA, poly-
Elastogenic outcomes with polyurethane (PU)-based scaffolds have been mixed. Early work by Robinson et al., 107 who implanted PU-based vascular prostheses intra-aortally in rats, showed that the scaffolds support the formation of a neoartery with elastin. Contradictorily, Grenier et al. 108 reported lack of elastic matrix deposition in 3D PU scaffolds, which was not improved even when the scaffolds were coated with fibronectin or matrigel, even though cells infiltrated and spread. However, the 3D architecture of PU scaffolds renders cells elastogenically responsive to factors such as TGF-β in a manner not seen in cell cultures on 2D PU substrates, 80 and this suggests that phenotypic modulation of cell behavior by scaffolding topography sensitizes them to elastogenic factors. This may circumvent the need to find scaffolding materials that directly coax cells to enhance elastic matrix synthesis. Microfibrous polyethylene terephthalate scaffolds were shown by Moreno et al. 109 to encourage elastic matrix deposition by human aortic SMCs. However, again, elastic matrix amounts were not quantified, nor formation of elastic fibers and superstructures (e.g., lamellae) investigated. Although not all inclusive, the body of work described above lacks uniformity in the studied cell types, test species, cell seeding conditions, and scaffolding parameters that render it difficult and, in many cases, inappropriate to directly compare elastogenic outcomes. However, the studies thus far appear to suggest that 3D scaffolding architecture, surface hydrophilicity, lack of pH-altering byproducts of degradation, and rapid changes to scaffolding compliance are major factors that either directly stimulate VSMCs to synthesize elastic matrix or sensitize them to exogenous elastogenic agents.
Contact Guidance as a Strategy to Improve Elastic Fiber Alignment
While it is clear that cellular elastic fiber deposition may be increased through careful selection of the substrate chemistry, the ability to guide elastic fiber alignment is also necessary. One way to do this is through careful design of the scaffolds' topographical features and use of the phenomenon of contact guidance. It has been shown, for example, that when PLGA scaffolds are surface-treated to incorporate nanometer-sized roughness, bladder SMCs seeded within exhibit increased adherence, proliferation, and deposition of matrix elastin compared to non-surface-treated PLGA. 110 In addition, the previously mentioned fact that SMCs respond differently on 3D surfaces is an indication that surface topography and, potentially, also the associated differences in surface compliance provide cues to direct cell attachment to the scaffold. This results in differences in cell alignment, cell phenotype, and elastic matrix production through mechanotransductive signaling mechanisms. By aligning the surface features, it is possible to also direct the alignment of most anchorage-dependent cells, including VSMCs. Since cells have been shown to be actively involved in directing collagen fibril111,112 and potentially elastic fiber alignment,113–115 aligned scaffold topography is an important tool in elastic matrix engineering. The exact role of cells in elastic fiber alignment is not currently clear, but they are thought to affect microfibril assembly and alignment through interactions with the cell surface. 115 Rossi et al. 113 have shown that on surfaces with aligned grooves, subconfluent endothelial cells align in the direction of the grooves and deposit fibrillin-containing microfibrils in that same direction. Conversely, they found that the same cells cultured on square patches did not align or deposit aligned microfibrils. A similar response can be expected for VSMCs. Elucidating the cellular and extracellular mechanisms that motivate and regulate elastic fiber orientation by VSMCs is therefore emphasized as an area of future research in the field.
Enabling Elastic Matrix Deposition in a 3D Space
It is important that the cells infiltrate and deposit elastic matrix throughout scaffolds, as would be found in medial layer of an artery. This in an important consideration with 3D scaffolds, where the size and interconnectivity of the pores must be large enough to allow cell infiltration and yet provide topographic cues. For cells to infiltrate, rounded cells must be able to either fit within the pores or deform the surrounding features (e.g., fibers), thereby increasing the effective pore diameter. The pore diameters required for specific tissue engineering applications will vary between cell types and potentially the presence/absence of chemotactic factors or oxygen or nutritional gradients. Work by Ju et al. 116 demonstrated that human aortic SMCs did not infiltrate into electrospun PCL scaffolds with ≤1-μm-diameter fibers, with the larger fibers exhibiting larger effective pore diameters. However, increasing infiltration over 4 weeks was observed as the diameter increased to 2.39 and then 4.45 μm. 116 Similar results were found by Bashur et al., 82 with some of the rat aortic SMCs infiltrating within a PCL scaffold with ∼1-μm-diameter fibers, and yet with room for improvement.
Cell-Based Approaches to Elastic Matrix Engineering
Despite some progress toward being able to enhance elastin and elastic matrix synthesis by adult VSMCs, key challenges remain. These include inability to stimulate these cell types sufficiently to generate large, clinically useful elastic tissue constructs in a limited time period. In this context, an alternative approach may be to investigate the utility of other more elastogenic cell types for engineering or regenerating vascular tissues. For example, bladder SMCs, 20 as well as SMCs within female reproductive tissues (e.g., vaginal wall), 21 are known to retain their elastogenicity in adulthood, quite unlike VSMCs. Vaginal SMCs facilitate the enormous remodeling and regenerative repair of elastic matrix that occur in these tissues, postpartum, to enable resorptive involution. 21 If noninvasive biopsy of these human cells is made possible, their utility in a vascular tissue engineering/regenerative repair milieu may be investigated. However, this will require an understanding of phenotypic differences between vascular and such nonvascular SMC types and an investigation into the possibility of retraining these differentiated cell types to assume a phenotype more typical of VSMCs, while retaining their own inherent high elastogenic capacity. Recently, Campbell et al.117–119 reported on recruitment of macrophage-like cells from peritoneal fluid onto silastic tubes implanted within peritoneal cavities of rats and their production of a fibrous tissue capsule. Although the everted tubular tissue capsule exhibited distinct circumferential layers resembling the vascular tunics (i.e., a luminal mesothelium, an outer collagenous layer, and a layer of myofibroblasts in the middle), little elastin was observed in these constructs. Although a few elastic lamellae-like structures were found within these constructs over several months of maturation after autologous intra-aortal implantation, these were too sporadic to influence vascular mechanics in any tangible manner. Also, the cell types involved in these changes and their source are still unknown. Despite these drawbacks, combining the above concept of an in vivo bioreactor with 3D, fibrous scaffolds and elastogenic growth factors, as described previously, may prove to be a successful strategy to grow elastic vascular tissue constructs on demand.
Stem cells (SCs) may represent a novel source of autologous cells that could be useful for elastic matrix engineering in vitro or for in situ regenerative repair within proteolytically compromised vessels. Since most elastin in the body, including in vascular tissues, is primarily synthesized during fetal and neonatal development, as is evidenced by the far greater elastogenicity of neonatal SMCs relative to adult SMCs, 18 it is not far-fetched to hypothesize that SC-differentiated SMCs may retain the high tropoelastin mRNA expression and efficient ECM crosslinking machinery that adult VSMCs appear to lose with aging. A number of recent studies have demonstrated the feasibility of differentiating adult SCs and progenitor cells isolated from bone marrow,120–123 PB,124,125 and adipose tissues126–128 as well as embryonic SCs (ESCs)129–131 into VSMCs. However, to date, these SC-differentiated SMCs have not been studied in the elastogenic context beyond perhaps demonstrating their ability to express elastin mRNA, thus confirming their identity. Elucidating these aspects may lead to a future cell therapy or elastic matrix engineering approach appropriate for clinical application. Potential challenges include low numbers of adult SC types in respective source tissues and low yields during isolation, 132 which limit the scalable production of the undifferentiated SCs and of SCs differentiated into the desired lineage. In the last 5 years, induced pluripotent SC (iPSC) techniques have emerged as an innovative means of generating targeted and individualized cell lines by reprogramming somatic cells from patient's own tissues. iPSCs, which are pluripotent, can potentially circumvent the above limitations of adult SCs 132 and may eliminate the ethical concerns associated with the use of ESCs.
Clinical Progress and Elastic Fibers in Vascular Constructs
Since Weinberg and Bell first attempted to fabricate a completely tissue-engineered vascular graft over two decades ago, 32 Shin'oka et al. 133 and others have focused on developing the ideal tissue-engineered vascular replacement. However, as highlighted in this review, the lack of a mature elastic matrix in these grafts is one factor that has hindered their long-term clinical use as arterial replacements. Here we focus on two recent engineered vascular grafts developed by the companies Cytograft 134 and Humacyte 135 that are currently in clinical trials for low-pressure arterio-venous conduits or shunts. Cytograft developed a graft based on sheet-based tissue engineering technique where fibroblasts isolated from age- and risk-matched individuals undergoing vascular bypass surgeries were cultured in vitro to produce three concentric layers of cell–matrix sheets, which were then subjected to graduated pulsalite pressures. 136 Humacyte developed grafts by culturing cadaveric SMCs on a PGA felt scaffold under cyclic strains. 137 The PGA scaffold was replaced by a collagenous matrix synthesized by the cells within and were decellularized before implantation. Both these grafts have several advantages such as the absence of intimal hyperplasia, the ability to be stored long-term, and the presence of initial mechanical properties comparable to those of the sapheous vein and internal mammary artery. However, the long-term mechanics of these grafts are yet unknown and both products lack the presence of any elastic matrix during or postimplantation, thereby limiting their application as arterial grafts.
Conclusion
In summary, through the use of these natural proteins, growth factors, and appropriate environmental cues, SMCs may be directed to deposit a mature elastic matrix, which is necessary for CV applications. The use of cell types (e.g., fibroblasts and SC-derived SMCs) other than VSMCs, which are likely to be the focus of future efforts to grow elastic fiber-rich vascular constructs, may, however, provide more impressive outcomes, even without elastogenic stimulation. Further research into the special at temporal cues found to regulate elastic fiber assembly and organization during the fetal and neonatal periods (i.e., when most elastic matrix deposition occurs) may also elucidate new and more potent factors that can be used to promote elastogenesis in tissue-engineered constructs or diseased-tissue microenvironments. In addition, the impact of these cues on all components of elastic fibers (e.g., fibrillin and fibulin) and on the size and ultrastructure of the elastic matrix will be important. Through an appropriate combination of current elastic matrix engineering technologies, and potential discovery of new ones, engineering patient-customized vascular replacements should become closer to a reality.
Footnotes
Disclosure Statement
No competing financial interests exist.