
Abstract
Recently, dental stem and progenitor cells have been harvested from periodontal tissues such as dental pulp, periodontal ligament, follicle, and papilla. These cells have received extensive attention in the field of tissue engineering and regenerative medicine due to their accessibility and multilineage differentiation capacity. These dental stem and progenitor cells are known to be derived from ectomesenchymal origin formed during tooth development. A great deal of research has been accomplished for directing osteoblastic/cementoblastic differentiation and neural differentiation from dental stem cells. To differentiate dental stem cells for use in tissue engineering and regenerative medicine, there needs to be efficient in vitro differentiation toward the osteoblastic/cementoblastic and neural lineage with well-defined and proficient protocols. This would reduce the likelihood of spontaneous differentiation into divergent lineages and increase the available cell source. This review focuses on the multilineage differentiation capacity, especially into osteoblastic/cementoblastic lineage and neural lineages, of dental stem cells such as dental pulp stem cells (DPSC), dental follicle stem cells (DFSC), periodontal ligament stem cells (PDLSC), and dental papilla stem cells (DPPSC). It also covers various experimental strategies that could be used to direct lineage-specific differentiation, and their potential applications in tissue engineering and regenerative medicine.
Introduction
To date, various stem cells, such as embryonic stem cells (ESCs), bone marrow-derived stem cells (BMSCs), adipose tissue-derived stem cells (ADSCs), and other tissue-specific somatic stem cells, have been extensively used in tissue engineering. Manipulation of culture condition and microenvironment for directing desired cell lineage differentiation to a certain extent has also been extensively studied.7–14 These stem cells have been used to generate three-dimensional (3D) tissue-like structures by combining the cells with suitable biomaterials and cultivation systems.15–19 However, due to the lack of accessibility, autologous implantation, multilineage differentiation capacity, and issues related to aged cell populations, there is great limit for using these cells in practical applications.20,21 For instance, the isolation of BMSC and ADSC need surgical procedure to procure tissues such as bone marrow aspiration process and liposuction process. For these reasons the search for new alternative cell sources that require noninvasive or minimally invasive procurement process is ongoing.
Recently, the presence of multi-potent stem cells in periodontal tissues, such as dental pulp, dental follicle, dental papilla, and periodontal ligament, and in exfoliated deciduous teeth was reported.22–27 The recent finding of stem cells in periodontal tissues has suggested the use of these dental stem cells as a potential cell sources for tissue engineering (Fig. 1). Dental stem cell banks have been pioneered by private companies worldwide, and the dental stem cells are cryopreserved at stem cell bank for their potent availability in near future. Pulp, periodontal ligament, and papilla tissue can be procured from the discarded teeth, and dental stem cells can be isolated from the collected tissues by collagenase treatment, 22 which provides noninvasive procurement process. In addition, it has been well known that functional ability and self-renewal ability of somatic stem cells show a significant reduction with increasing donor age.20,21,28 In other hands, wisdom teeth and baby teeth developing from age 6 to age 20 after birth undergo organogenesis, and immature periodontal tissues in them are evident to have a pool of stem cells with high regeneration ability, which might be a good source for tissue engineering and regenerative medicine. After isolation, dental stem cells are expanded well in culture, and interestingly a cell population with stem cell characteristics appears to increase with passage during their expansion, but the underlying biology of this increased stem cell population during expansion needs to be further elucidated. 29 Benefits of this source for tissue engineering application include relatively easy accessibility, multilineage differentiation capacity, and possible autologous implantation. There has been extensive research on forming mineralized or neural tissue using dental stem cells in the field of regenerative medicine. In this review, we summarize recent research relating to multilineage differentiation, especially toward osteoblastic/cementoblastic and neural differentiation, of dental stem cells such as dental pulp stem cells (DPSC), dental follicle stem cells (DFSC), dental papilla stem cells (DPPSC), and periodontal ligament stem cells (PDLSC), and their potential applications to tissue engineering with comments on the progress of this approach and future trends of the field.
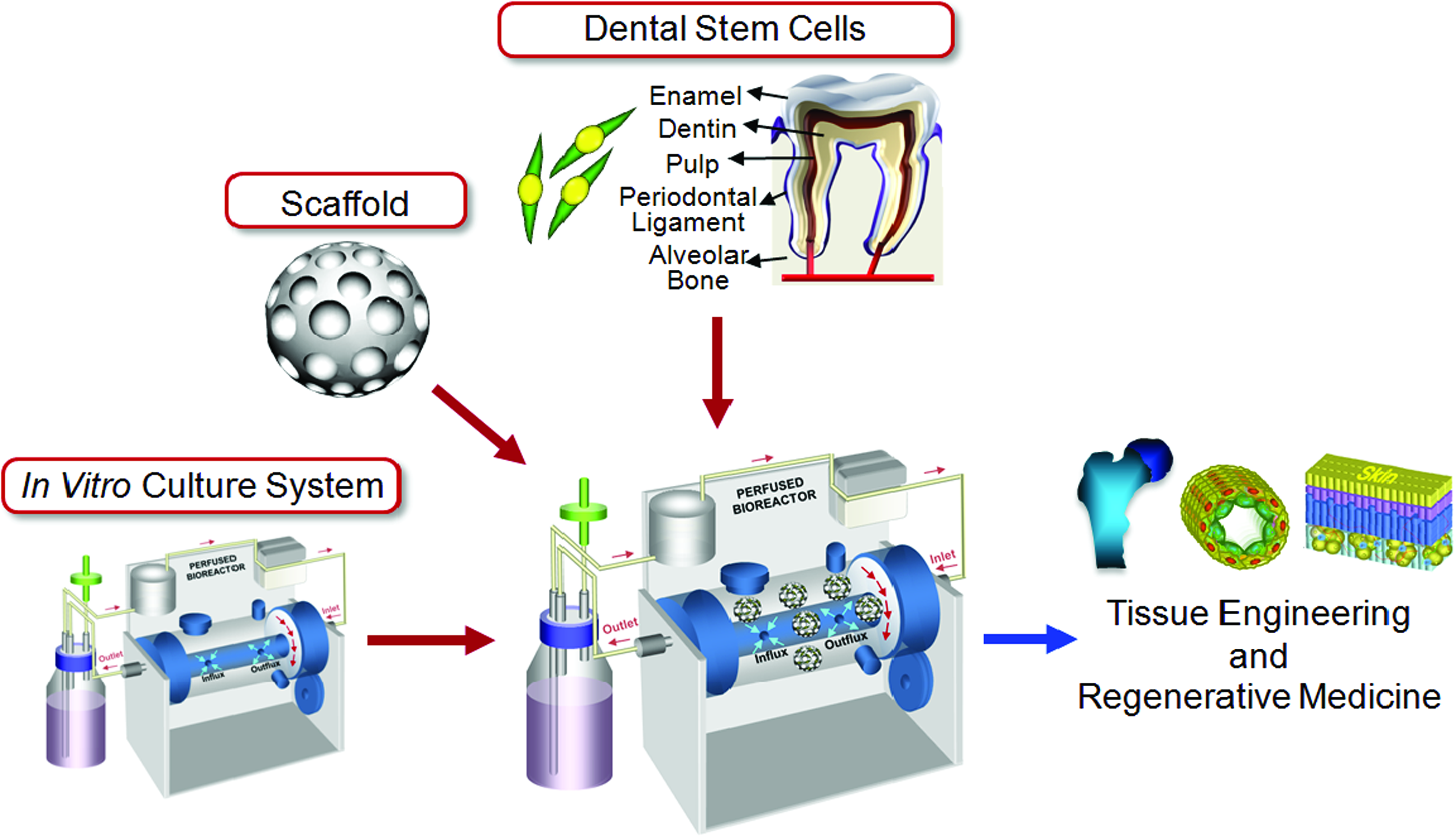
Dental stem cell-based tissue engineering. In vitro 3D tissue-engineered construct can be developed by combining dental stem cells with proper 3D cell carrier and bioreactor culture system, and can be applied to tissue engineering and regenerative medicine. 3D, three-dimensional. Color images available online at www.liebertonline.com/teb
Tooth Development, Dental Stem Cell Origin, and Multilineage Differentiation
To comprehend the stem cell characteristics of periodontal tissues, it is necessary to understand the biological processes that induce and regulate tooth development and the origin of dental stem cells. Tooth development involves the complex back and forth signaling between epithelial and mesenchymal tissues. Teeth are derived from oral ectoderm and neural crest-derived mesenchyme. 30 As shown in Figure 2, the process of tooth development is mainly composed of five stages. The first stage is the thickening stage, which involves the thickening of oral epithelium at the site of tooth formation. This is followed by the condensation of ectomesenchymes, which are derived from neural crest-derived mesenchyme, around the bud. During the second stage, also known as the bud stage, two distinct cell lineages, peripheral basal cells and stellate reticulum, are separated from dental epithalium to form two tissue layers providing the stem cell niche in growing teeth. The next two stages are the cap stage and the bell stage. During these stages, the cervical loop structure is developed, and the inner enamel epithelium is formed within the loop bordering the dental papilla. Also, during this stage the dental follicle is formed at the interface of the outer enamel epithelium. This cervical loop structure is maintained in developing teeth. During these stages, the dental papilla is surrounded by dental epithelium. The cells at the interface of the epithelial-mesenchyme within the cervical loop structure give rise to ameloblasts and odontoblasts, which produce a mineralized matrix of enamel and dentin. The pulp is formed from dental papilla, and cementoblasts and periodontal tissues are formed from the peripheral dental follicle. The growing enamel knots within the cervical structure regulate the size and shape of the tooth crown during tooth development. During the fifth and final stage, the secretory stage, the developed tooth erupts through the gums. 30

Developmental process of tooth morphogenesis. Tooth development is initiated by the interaction of ectomesenchyme and oral ephitelium and is composed of six sequential steps such as thickening, bud, cap, bell, secretory, and eruption. Color images available online at www.liebertonline.com/teb
The mature tooth is mainly composed of enamel, dentin, pulp, and periodontal ligament tissue. Human teeth can be mainly categorized into baby teeth and permanent teeth according to age. In addition, wisdom teeth (third molar teeth) are generally developed in people aged from 16 to 20 or older. All of these types of teeth have dental stem cells. Recently, the presence of multi-potent stem cells has been reported within various periodontal tissues. Yamazaki et al. demonstrated the presence of the neural crest-derived dental mesenchyme in developing tooth by tracing genetically modified neural crest-derived cells in a mouse model. 31 They also reported the potential of these cells to differentiate down multiple pathways into odontoblasts, chondrocyte-like cells, and osteoblast-like cells. 31 It has also been reported that multi-potent stem cells, expressing mesenchymal stem cell marker such as STRO-1 and CD146/MUC18, are present in periodontal ligament tissue extracted from human third molar teeth. These PDLSCs could be isolated by single colony selection and an immunomagnetic isolation method using anti-STRO-1 antibody. These stem cells showed the multi-differentiation capacity to form both cementoblast-like cells and adipocytes. 26 Gronthos et al. isolated a clonogenic and highly proliferative stem cell population from adult human dental pulp tissue by enzymatic digestion. 22 It has been shown that the typical immunoreactivity profile of these stem cell populations shows a very similar pattern as bone marrow stromal cells. These DPSCs can also produce dentin/pulp-like structures on hydroxyapatite (HA)/tricalcium phosphate (TCP) scaffolds after transplantation in immunodeficient mice in conjugation with HA/TCP powder, indicating their differentiation potential. 22 In addition to PDLSC and DPSC, DPPSCs and DFSCs have also been isolated from papilla and follicle tissue from third molar teeth by collagenase treatment. These stem cells had a fibroblastic morphology and stained positive for various mesenchymal stem cell or putative stem cell markers, such as CD29, CD44, CD105, or Notch-1.24,25 In vitro differentiation studies of those stem cells have reported potent osteogenic differentiation capacity in the presence of osteogenic supplements.24,25
To date, in addition to osteogenic differentiation, several differentiation studies of dental stem cells have reported on their capacity to differentiate into chondrogenic, adipogenic, neural, and endothelial cell lineages.23,26,32,33 For instance, adipogenic differentiation was induced by the addition of 0.5 mM isobutyl-methylxanthine, 1 mM dexamethasone, 10 mM insulin, 200 mM indomethacin, and 50 mg/mL of gentamicin into alpha minimum essential medium (α-MEM) containing 10% fetal calf serum (FCS). 32 Such adipogenic differentiation capacity was also reported in PDLSC differentiation studies.23,26 In addition, despite that chondrogenic differentiation activity was relatively lower in comparison with BMSC, chondrogenic differentiation of dental stem cells could be achieved by the micromass culture of DPSC in the presence of 10% FCS, 200 μg/mL ascorbic acid, and 10 ng/mL of TGF-β3 after 6 weeks. 34 In addition to skeletal lineage differentiation of dental stem cells, different lineage differentiations such as neural and endothelial differentiation could be induced under certain culture conditions.32,35 Neural differentiation was induced by the culture in the presence of 10 mM β-mercaptoethanol, 2% dimethyl sulfoxide, and 200 mM butylated hydroxyanisole without serum. 32 Nakasima et al. reported in vitro and in vivo endothelial (endothelial cell lineage) differentiation capacity of DPSC. CD31−/CD146− side population cells and CD 105+ cells were isolated from dental pulp tissue, and these cells generated vascular-like branching structures on Matrigel and generated newly formed vasculature after transplantation. 35 Dental stem cells from exfoliated deciduous teeth differentiated into vascular endothelial cells in an implantation study. 36 In view of their stem cell properties and multidifferentiation activity, there is a growing interest in the use of dental stem cells isolated from various periodontal tissues in the field of stem cell biology and tissue engineering. The sources of dental stem cells and their multi-differentiation strategy are illustrated in Figure 3.
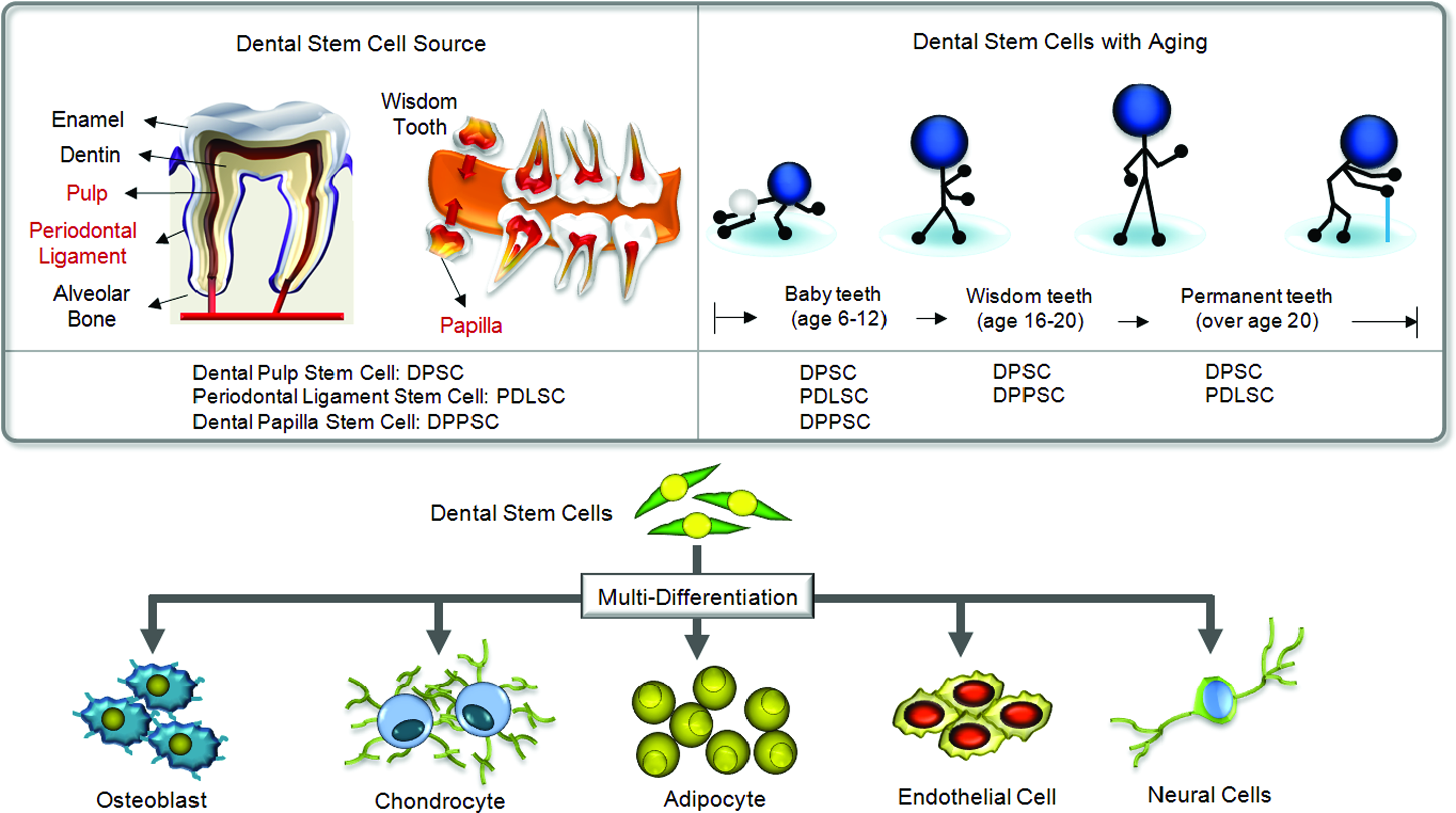
Dental stem cell sources and their multilineage differentiation capability. Various dental tissues can be used for dental stem cell's isolation according to age. Color images available online at www.liebertonline.com/teb
Osteoblastic/Cementoblastic Differentiation of Dental Stem Cells
The use of stem cells for forming hard tissues such as bone has received a great deal of attention, and various stem cells such as BMSCs, ADSCs, and ESCs have been widely used in osteogenic differentiation studies and bone tissue engineering. Along with these stem cells, dental stem cells have also proven themselves to be a potent cell source for fabrication of 3D bone tissue structures. This is due to their good osteogenic activity potentially as a result of their biological function to form hard mineralized tissue during tooth development. For example, it has been shown that c-kit+/CD34+ cells (dental pulp stem cell, DPSC) could be purified from dental pulp tissue, and differentiate into CD44+/RUNX-2+ osteoblast precursors and subsequently into osteoblasts in α-MEM containing 20% fetal bovine serum (FBS), ascorbic acid-2-phosphate, and

In vitro osteogenic differentiation of DPSCs. Stem cells were isolated form dental pulp tissue by purifying STRO-1-positive cell population with MACS system, and allowed to differentiate into osteogenic lineages to form mineralized nodules under proper osteogenic culture condition.
In addition to osteoblastic/cementoblastic differentiation of DPSC, the osteogenic differentiation of PDLSC and DPPSC has been also reported. The STRO-1+ cell population, PDLSCs, was isolated by a magnetic activating cell sorting (MACS) system and allowed to differentiate in the presence of ascorbic acid-2-phosphate. This resulted in the expression of alkaline phosphatase, mineralization-related genes, and the formation of mineralized nodules, thus indicating osteoblastic/cementoblastic cell lineage differentiation. 26 In another study, Ikeda et al. reported in vitro osteoblastic/cementoblastic differentiation of DPPSCs in the presence of ascorbic acid, β-glycerophosphate, and dexamethasone. This differentiation was evidenced by high alkaline phosphatase activity, osteocalcin content, and mineralized nodule formation. 25
Recent hard tissue engineering studies have shown the potential for use of dental stem cells in combination with 3D polymeric scaffolds. For example, a recent report showed the formation of bone-like hard tissue formed by DPSC in conjugation with a 3D porous calcium phosphate ceramic carrier in an implantation study. The STRO-1-selected DPSCs were seeded onto a HA/TCP (HA/TCP ratio of 60/40) Camceram® ceramic disc with a porosity of 90%, and in vitro osteogenic differentiation was induced in the presence of bone morphogenic protein-2, dexamethasone, β-glycerophosphate, and ascorbic acid. After 8 days of in vitro culture, these constructs were implanted into BALB/c immunocompromised mice and analyzed after 12 weeks. Histological analysis showed well-mineralized hard tissue formation with distinct concentric lamellae structure and with partially developed bone marrow-like hematopoietic tissue. 39 In another study, mineralized tissue formation was reported in an autologous implantation study with DPSCs and a poly(lactic-co-glycolic) acid scaffold construct. 40 In the study, DPSCs were isolated from New Zealand white rabbits, and cultured on a 3D scaffold for 2 days before implantation. Afterward, these autologous DPSC/scaffold constructs were transplanted subcutaneously into rabbits, to result in the formation of a mineralized structure containing osteodentin with a well-organized cell arrangement and blood vessels. 40 In another study, dental stem cells were used to generate a functional bioengineered tooth replacement. 41 In this research, a 3D bioengineered molar tooth germ was fabricated by encapsulating a molar tooth germ-derived epithelial cell and a mesenchymal stem cell mixture with collagen gel. This 3D construct was then implanted into the alveolar bone region of a lost tooth. After transplantation, mineralized hard tissue with the correct tooth structure was developed. This bioengineered tooth was functional in a similar pattern with natural teeth, and proved to be responsive to mechanical stress. 41 These studies and findings might suggest the potential use of dental stem cells or dental stem cell-based tissue-engineered 3D construct for hard tissue regeneration. Briefly, in vivo hard tissue formation using dental stem cells are summarized in Table 1.
DPSC, dental pulp stem cells; PDLSC, periodontal ligament stem cells; DPPSC, dental papilla stem cells; HA, hydroxyapatite; TCP, tricalcium phosphate; PLGA, poly(lactic-co-glycolic) acid; BMP, bone morphogenetic protein; 3D, three-dimensional.
Neural Differentiation of Dental Stem Cells
To date, extensive trials to generate neural cells have been done using various adult stem cells, such as MSCs, ADSCs, and neural stem cells (NSCs). The goal of these studies is to use stem cells for nerve regeneration in patients with neurodegenerative diseases. Although several successful approaches have been reported on inducing neural differentiation from these stem cells, there are a number of potential limitations in their medical application in terms of efficiency in differentiation and accessibility of these cell sources. In spite of several reports for neural differentiation of MSC and ADSC,45–47 the precise mechanism of trans-differentiation between MSC or ADSCs and neural lineage cells has not been well elucidated and remains controversial.
Unlike other somatic stem cells, it has been well documented that dental stem cells, when isolated from immature dental pulp during tooth development, show expression of ESC pluripotency markers, such as Oct-4, SSEA-4, Nanog, and TRA-1–60. These stem cells also display multipotency markers indicating chondrogenic and osteogenic tissue formation in differentiation culture and spontaneous neural differentiation; this represents stem cells of neural crest-derived mesenchyme (ectomesenchyme) origin of the dental papilla. 48 From scientific knowledge of developmental biology, the neural differentiation from dental stem cells can be supposed. A great deal of research has been done on the generation of neural lineage cells from dental stem cells. Dental stem cells express various neural markers upon stimulation with a neural differentiation medium. The neurosphere formation and neural differentiation of DPSC, isolated from teeth by MACS using anti-STRO-1 antibody, under neural culture conditions are shown in Figure 5. The Stro-1-positive stem cell population was allowed to form neurospheres in suspension culture in the presence of fibroblast growth factor 2 (FGF2); the formed neurospheres were replated on laminin-coated culture dishes, and positive expression of neural cell markers such as Ca2+/calmodulin-dependent protein kinases II (Cam kinase II), glial fibrillary acidic protein (GFAP), βIII-tubulin, and tyrosine hydroxylase was detected under neural differentiation condition containing glial cell-derived neurotrophic factor, sonic hedgehog homolog, and FGF 8.
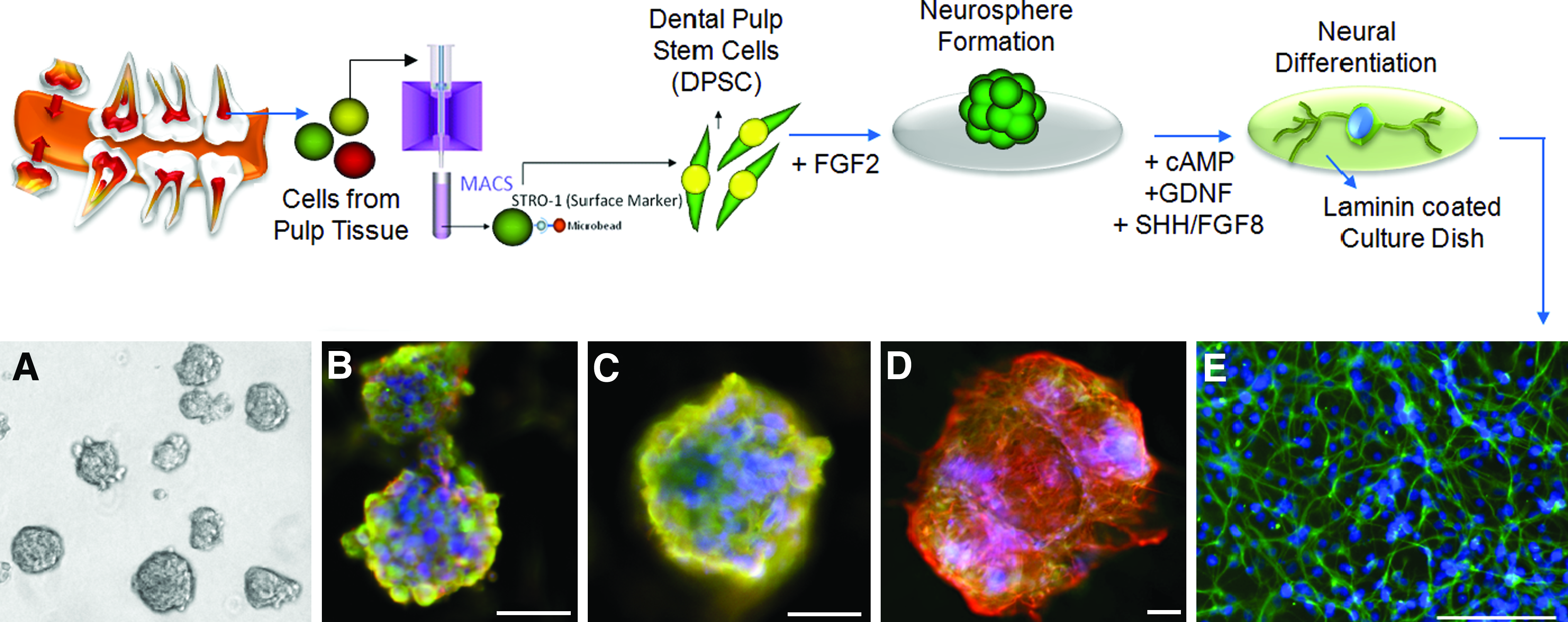
In vitro neural differentiation of DPSCs. STRO-1-positive stem cells were allowed to form neurosphere in the presence of FGF2 under suspension culture condition, and differentiated into neural lineage by plating on laminin-coated culture dishes and culturing in the presence of neurotrophic factors such as GDNF, Shh, and FGF8.
It has been reported that DPSCs express the neural stem cell marker, nestin, before exposure to differentiation culture, and that the expression of nestin was downregulated upon induction of differentiation. Also, during the course of maturation into neural lineages, mRNA of neural cell markers such as GFAP and βIII-tubulin was upregulated in DPSC differentiation culture in the presence of N-2 plus media supplement, FGF, and epidermal growth factor (EGF), which indicate DPSC differentiation into glial cells and neurons. 49 In addition, it has been reported that dental stem cells form 3D cell clusters, which mimic the so-called neurospheres, when cultured on plates with low cell binding surface or in suspension environments in the presence of EGF and bFGF. These neurospheres, which are self-aggregated cell clusters composed of NSCs, have been widely used for the induction of neural differentiation from dental stem cells. There has been a great deal of research performed to establish more effective culture strategies for neurosphere formation and neural induction.49–52 An immunophenotype study showed a strong positive reactivity of cells within these spheres against antibodies of NSCs markers, such as anti-nestin and anti-CD81 antibodies. The factor bFGF was found to be the most effective means of generating neurospheres from dental stem cells. This factor was even more effective than other growth factors such as BMP2, brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), and others. 50 In another study, Widera et al. developed a well-defined, serum-free medium for the propagation of neurospheres formed from dental stem cells with supplements of EGF and FGF-2. Neurospheres, showing a strong expression of nestin and Sox-2, were formed from PDLSCs in this serum-free condition. Spontaneous differentiation into neuronal lineages was observed when plated on laminin-coated plates in the absence of cytokines. 51
The neural differentiation activity of stem cells isolated from papillar tissue, DPPSC, has also been reported. It is well known that the dental papilla converts to dental pulp tissue during tooth development. This implies that the characteristics of stem cells in dental papilla are closer to ectomesenchyme than DPSC in terms of maturation. 53 DPPSCs were isolated from the apical papilla tissue of human immature permanent teeth, and their proliferation activity was found to be two- or threefold higher than DPSCs. These DPPSCs were found to express a variety of neuronal markers such as nestin, GFAP, βIII-tubulin, and neurofilament M upon induction of neural differentiation with bFGF and EGF. 54 It has also been known that DFSCs are derived from immature dental tissue of wisdom teeth and also possess this multilineage differentiation capacity. 55 It has been reported by the Morszeck group that DFSCs can form neurosphere-like structures when plated onto low-attachment cell culture dishes in serum-free medium containing EGF and FGF-2. They also discovered that DFSC-derived neural cells grown on laminin and gelatin display a relatively long axon-like cell extension, indicating that neural cell morphology and axon-like cell extensions were dependent on cell culture substrates. Also, they reported that the neuronal cell lineage differentiation into dopaminergic or GABAergic neurons was dependent on the time-dependent addition of neural stem cell supplements, such as EGF, FGF-2, and retinoic acid. 55
In addition to studies on the in vitro differentiation of dental stem cells into neural cells, there have been attempts to improve and regenerate nerve tissue via cell transplantation. One recent study demonstrated that implanted DPSCs support host axon guidance within a receptive host nervous system by secreting neurotrophic factors. Human DPSCs were isolated from the third molar teeth. Stable green fluorescence protein (GFP)-positive DPSCs were established by viral transfection with the GFP encoding gene. These cells were then injected into the mesodermal tissue of a developing avian embryo. In this xenotransplantation, it was observed that avian trigeminal ganglion axons were guided toward implanted DPSCs via chemo-attraction between CXCL12, a stromal cell derived factor-1 that is secreted by implanted DPSCs, and its receptor CXCR4. 56 It is also well known that cultured DPSCs produce several neurotrophic factors, such as NGF and BDNF. These neurotrophic factors also serve to promote the survival of injured neurons. 57 In a similar study, DPSCs were implanted into the hippocampus of immunocompromised mice, which increased host neural cell proliferation and stimulated neural differentiation to mature neural cell types by the recruited host neural cells at the site of the graft. Enhanced expression of vascular endothelial growth factor, ciliary neurotrophic factor, NGF, and FGF was observed up to 30 days after implantation at the same site. 58 It has also been reported that maintaining the proper environment of a cell culture can induce the differentiation of DPSCs into functionally active neurons in vitro. This was demonstrated by the actuation of sodium current in DPSC-derived neuronal cells grown in a neuronal inductive media. Moreover, in an avian xenoplantation assay, the in vivo neuronal differentiation of implanted human DPSCs was induced by the endogenous neuronal environment. This differentiation was proved by the morphological change from dental to neuronal cell and by immunohistochemical staining with human specific anti-βIII tubulin and anti-neurofilament M antibody. As a control, there was no morphological change and no immunostaining of implanted human foreskin fibroblast at same injection area. 59
Many in vitro differentiation studies and in vivo animal studies for neural differentiation of dental stem cells showed that dental stem cells have been proved to have neural differentiation capacity under proper culture condition and have inducing activity to regenerate neuronal defect by promoting host neuron's migration or self-differentiation into neurons in implanted animal. These studies suggest that there may be potential uses of dental stem cells in the field of neural tissue engineering and that these stem cells may one day be used for nerve regeneration. However, the identity of dental stem cells and the underlying biology for neural differentiation of dental stem cells has not been elucidated. It was reported that stem cells from dental tissues showed mesenchymal stem cell characteristics, regarding their multidifferentiation capacity into especially skeletal lineage and self-renewal activity. 60 Also, dental stem cells were reported to show another stem cell property resembling ectomesenchymal stem cells having both mesenchymal lineage differentiation capacity and neuroectodermal lineage differentiation capacity 26 regarding their developmental origin is from neural crest-derived ectomesenchyme. 27 Furthermore, nothing has been proved for the presence of neural stem cell within dental tissues and its possible direct differentiation into neural lineage. Hence, it is necessary to gain further insight into the characteristics of postnatal stem cells from dental tissues.
Future Challenge
Since the discovery of stem cells in periodontal tissues, many researchers have successfully derived lineage-specific differentiation, osteoblastic/cementoblastic, and neural differentiation of dental stem cells. These differentiated cells have been applied to the field of tissue engineering and regenerative medicine. The goal of this research was to successfully regenerate mineralized or neural tissue. In this review, it was suggested that the use of dental stem cells can be an alternative potent source in the application to tissue engineering and regenerative medicine regarding their relatively easy accessibility and multidifferentiation capacity.
However, ultimately, from a therapeutic point of view, there still remain many issues to resolve regarding expansion, differentiation efficiency, and safety in the use of dental stem cells. Dental stem cells have been known to have a great self-renewal activity, and more than 80 passages of DPSC were obtained without losing stem cell characteristics and clear signs of senescence. 61 The successful isolation and subsequent culture to maintain self-renewal activity of dental stem cells could be achieved regardless of dental tissues with aging. Dental stem cells are able to be isolated from adults' dental tissues such as papilla and pulp tissue with various ages, 19–45.37,62 However, age-related changes of differentiation activity of dental stem cells should be taken into account for applications. 37 Osteogenic differentiation capacity was reported to be dependent on age of dental tissues, 63 and some dental tissue-derived cells showed no mineralization activity. 37 Another limit to overcome for dental stem cell application is aging-related dental diseases. Dental pulp diseases such as pulpitis and pulp cavity calcification increase with aging, which means that the limited availability of dental tissue for dental stem cell isolation. It has been known that the small amount of periodontal tissue such as dental pulp and periodontal ligament can be procured from discarded teeth, and a small population of stem cells, between 1% to 10% of the total cell population, within adult (mature) dental tissue was observed.62,64 Considering the paucity of available dental tissues, a more efficient strategy for dental stem cell isolation and in vitro expansion will be needed to generate the large numbers of stem cells that are needed for successful transplantation. In addition, dental stem cells have not yet been extensively studied in terms of age-related cellular change regarding stemness maintenance and differentiation, which should need to be investigated to broaden their therapeutic applications.
The conditions required to induce osteoblastic/cementoblastic or neural differentiation need to be further characterized especially as they apply to dental stem cells collected from different periodontal tissues. Before there can be any successful clinical usage of these dental stem cells, a well-defined FBS-free medium needs to be developed for cell growth and differentiation. This medium needs to be pure in order to remove any possible cross contamination and immune reaction due to factors derived from other species. In addition, although there has been no report for neuroma formation in DPSC transplantation studies for nerve regeneration, the possibility of dental stem cell-derived tumor or cancer formation should need to be investigated further regarding high self-renewal activity and not well-known characteristics of immature tissue (dental papilla)-derived stem cell to confirm biological safety of dental stem cell transplantation in regenerative medicine. Finally, the development of an in vitro 3D culture strategy using suitable 3D biomaterials and a 3D culture system would also constitute an important step toward fabricating a functional 3D tissue like construct for applications to tissue engineering and regenerative medicine. Such a construct would be invaluable for using dental stem cells to replace damaged mineralized and nerve tissue.
Footnotes
Acknowledgments
This research was supported by Grant No. 20090065530 and No. 20100003398 from the Basic Research Program of the Korea Science & Engineering Foundation and by Grant No. A100208 from Korea Health Industry Development Institute.
Disclosure Statement
No competing financial interests exist.