
Abstract
Articular cartilage repair and regeneration provides a substantial challenge in Regenerative Medicine because of the high degree of morphological and mechanical complexity intrinsic to hyaline cartilage due, in part, to its extracellular matrix. Cartilage remains one of the most difficult tissues to heal; even state-of-the-art regenerative medicine technology cannot yet provide authentic cartilage resurfacing. Mesenchymal stem cells (MSCs) were once believed to be the panacea for cartilage repair and regeneration, but despite years of research, they have not fulfilled these expectations. It has been observed that MSCs have an intrinsic differentiation program reminiscent of endochondral bone formation, which they follow after exposure to specific reagents as a part of current differentiation protocols. Efforts have been made to avoid the resulting hypertrophic fate of MSCs; however, so far, none of these has recreated a fully functional articular hyaline cartilage without chondrocytes exhibiting a hypertrophic phenotype. We reviewed the current literature in an attempt to understand why MSCs have failed to regenerate articular cartilage. The challenges that must be overcome before MSC-based tissue engineering can become a front-line technology for successful articular cartilage regeneration are highlighted.
Introduction
U
Our group and others have been studying chondrogenesis and basic aspects of cartilage repair for at least three decades. While substantial progress has been made toward understanding the cellular and molecular control of the chondrogenic program, the current state of the art of TE cartilage repair still falls short of clinical expectations. Although chondrocytes have been extensively studied and used in TE cartilage, the introduction of culture-expanded mesenchymal stem cells (MSCs) generated a considerable interest in their use within the orthopedic community. Cartilage constitutes one of the potential differentiation lineages that MSCs can follow. Therefore, MSCs are considered a cell source for cartilage repair. It is necessary to rely on external factors (mainly inductive morphogens) to push MSCs to develop into specific phenotypes, and even then, the resulting tissue differs significantly from articular cartilage in terms of structure, chemical composition, cell phenotype, and function. Abundant experimental evidence from several groups suggests that a transient cartilage typical of endochondral processes, such as bone formation and fracture healing, rather than hyaline articular cartilage, is the default lineage intrinsic to marrow-derived MSCs.3–5 In this article, we focus on MSC-based cartilage TE and review the major unresolved issues that underlie the shortcomings in current articular cartilage TE. Principally, over and above, producing simple hyaline cartilage, the natural three-dimensional subarchitecture of articular cartilage, must be achieved. The goal of this review was to identify the shortcomings in current cartilage TE, so that we can continue to progress toward clinically relevant technologies.
Significance
MSCs do not make hyaline cartilage
Whether MSCs can function therapeutically as an exogenous cell source or can mobilize endogenous chondroprogenitor cells that function to resurface, repair, or regenerate joint tissue is still uncertain. Since the work of Johnstone et al., Barry et al., and Pittenger et al., it is known that bone marrow-derived MSCs (BM-MSCs) can form a cartilage-like tissue in vitro under the guidance of specific cocktails of growth factors.6–9 The resulting differentiated tissue can be classified as cartilage in that it expresses many biomolecules typical of hyaline cartilage, such as type II collagen and the proteoglycan, aggrecan. However, the proportion of the chemical constituents tends to be wrong for true weight-bearing hyaline articular cartilage. For example, in TE cartilage, collagen content is generally less than 50% of what is found in native healthy adult cartilage, with negative implications for tensile strength and load-carrying capabilities.10–15
Similarly, the stratified ultrastructure and spatial organization of native cartilage are completely absent, resulting in poor mechanical properties. While it was once thought that articular chondrocytes were a uniform population, it is now obvious that there are differences in morphology, proliferative capacity, and rates of extracellular matrix (ECM) synthesis controlled by where the chondrocyte resides in the depth of the cartilage tissue. 16 Native articular cartilage has a stratified ultrastructure with biochemical composition and biophysical properties specific to each of its zones. Articular cartilage typically comprises three horizontal zones: superficial, middle, and deep.16,17 Each zone has a distinct matrix composition, local oxygen tension, and biomechanical capacities. For example, superficial zone chondrocytes are flattened discoid and secrete surface zone protein (SZP), a proteoglycan that decreases the friction between the two articular plates. 18 The middle zone is characterized by a seemingly random fibrillar structure and matrix producing-rounded chondrocytes.19,20 The deep zone makes up about 30% of the cartilage volume and consists of large diameter collagen fibrils oriented perpendicularly to the articular surface. Chondrocytes in the deep zone are larger, elongated, and tend to arrange themselves in columns. 21
Several decades-old clinical treatments for articular cartilage defects are based on the known capacity of MSCs to differentiate down the chondrogenic lineage. For instance, microfracture or marrow stimulation relies on creating a passage between the articular space and the underlying bone marrow that would allow the migration of MSCs into the cartilage defect. Even with direct access to MSCs, this technique typically results in fibrocartilage formation (which often temporarily reduces clinical symptoms), which is not the correct articular cartilage phenotype. It is possible that local microenvironmental conditions at the defect may interfere with the natural healing capacity of the MSCs and their differentiation potential.22,23
In MSC-based aggregate-culture chondrogenesis, a wide range of hypertrophy-associated genes are expressed at the RNA and protein levels, including type X collagen, alkaline phosphatase (ALP), and matrix metalloproteinases (MMPs), and cellular hypertrophy is evident as well.6,24–30 The expression of these genes in MSC-derived chondrocytes mimics a hypertrophic phenotype and is a potential limitation to their application in articular cartilage TE.9,25,29–33 Joint articular cartilage does not normally undergo hypertrophy, except under severe pathological conditions.34–40 In fact, this sequence of events and the morphology of the resulting cartilage are more reminiscent of the early phases of endochondral ossification as seen in the skeletal development or fracture repair, where it is a prelude to programmed cell death and mineralization.9,25,29–33,38,41–43 Completing the terminal differentiation motif, mineralization of the cartilage ECM has been noted in in vitro aggregate cultures, 43 and Pelttari et al. showed that, after ectopic implantation in SCID mice, MSC-derived cartilage aggregates mineralize, vascularize, and form ossicles, further supporting an intrinsic endochondral bone formation behavior of MSCs. 29 Overall, this is undesirable from a cartilage TE/repair perspective, and maintaining a stable, prehypertrophic cellular phenotype is therefore essential for the long-term stability of a TE product in the joint in vivo.
Several recent studies have focused on delineating culture conditions and factors that control the terminal hypertrophic progression of MSC-derived chondrocytes.44–47 Transforming growth factor beta (TGF-β) is necessary to induce chondrogenesis in human MSCs (hMSCs) and, as in growth-plate chondrocytes, 48 it inhibits hypertrophy for at least 7 weeks in MSC-based aggregates. However, chondrogenic MSCs under TGF-β stimulation still express many hypertrophy-related genes, such as type X collagen, ALP, MMP13, VEGF, PTHR1, and RUNX2.30,49,50 Varghese et al. showed that the presence of chondroitin sulfate (CS) in the scaffold material prevents or delays the lineage progression to the hypertrophic phenotype. 44 The mechanism remains unclear, but direct interactions between the cells and the CS, or CS effects on growth factor storage/binding or presentation, may play a critical role.
While it is plausible that the hypertrophic phenotype constitutes the default differentiation pathway of bone marrow-derived MSCs, it may also be that biochemical cues, such as TGF-β, must be very tightly controlled, both quantitatively and temporally. These dynamics are predicated on developmental events; however, the actual process of formation and maintenance of stable articular cartilage in adults is still not well understood. Biomarkers that correlate with cell hypertrophy, vascular invasion, matrix calcification, and bone formation in vivo will all become potentially powerful tools to monitor this progression when these markers are delineated.9,29
Realistically, we are still far from achieving the complexity that is associated with the generation of new cartilage, especially considering that the generation of stable articular cartilage during embryonic and postnatal development is far from being understood at the cellular and molecular levels. Hyaline cartilage has been widely studied, given its important function during development as a framework for future skeletal elements that arise following endochondral ossification (transient cartilage) and its function at the joint interfaces (permanent cartilage). However, other types of cartilage are present throughout the body at specific locations (Fig. 1), such as fibrocartilage (e.g., between intervertebral discs) and elastic cartilage (e.g., external ear). Hypertrophic cartilage represents a lineage stage in differentiation or disease in each cartilage subtype that will be replaced by vasculature and marrow and eventually bone tissue. 51 This cartilage classification is based on molecular and structural differences. 52 The controlling details of how these different cartilages are formed are not fully understood (Fig. 2).
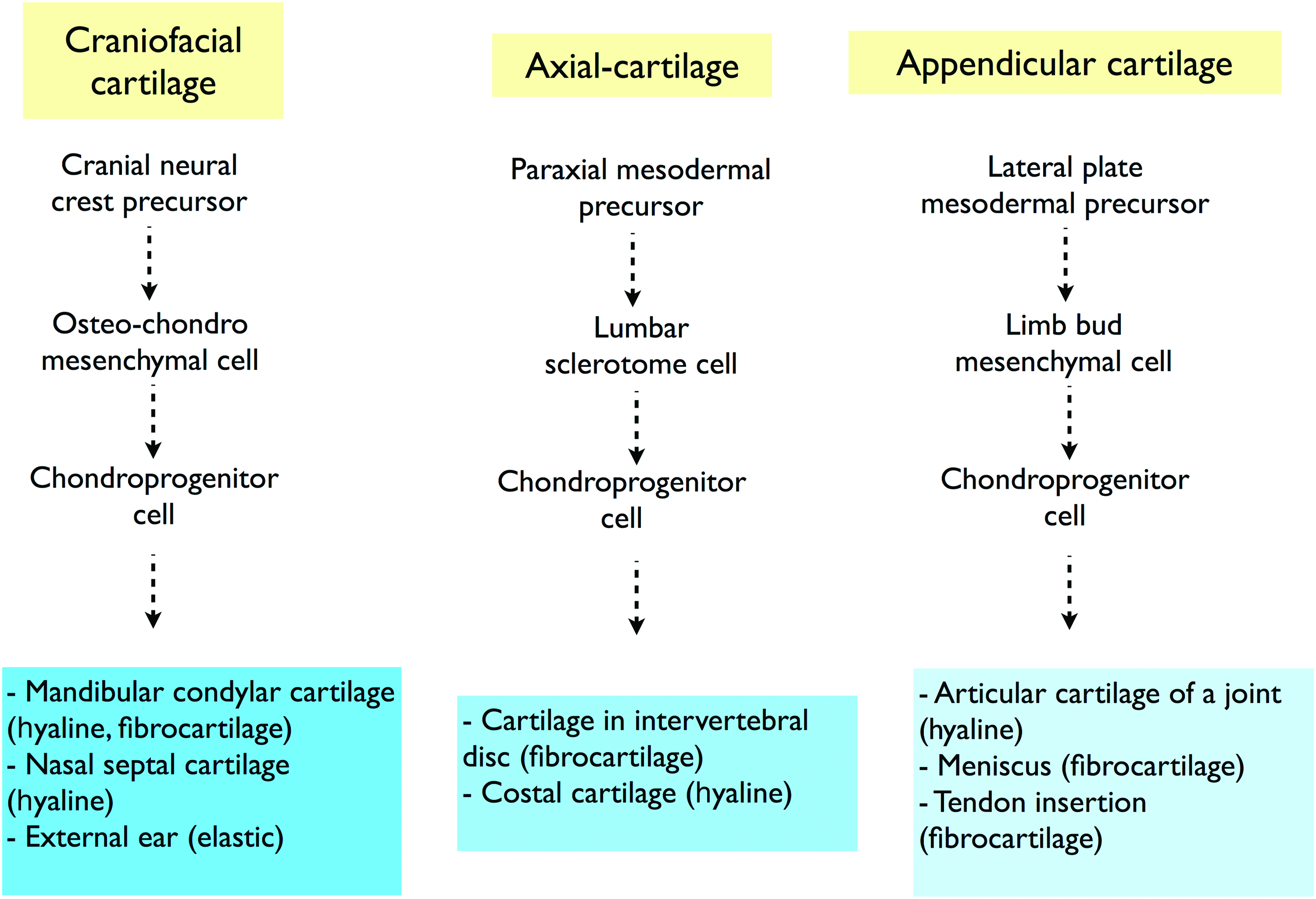
Cartilage development. Oversimplified scheme of cartilage development highlighting the generation of different types of cartilages derived from the cranial neural crest (craniofacial cartilage), paraxial mesoderm (axial cartilage), and lateral plate mesoderm (appendicular cartilage). Cartilage diversification may not be entirely dependent on tissue development; it seems probable that environmental factors are involved in cartilage determination. For example, hyaline cartilage can be derived from three different developmental lineage paths, whereas its functional specification is determined by its specific anatomic location. Thus, we suggest that these hyaline cartilages are influenced by site-specific and different physiological and physical or mechanical signals. Color images available online at www.liebertpub.com/teb

Chondrogenic lineage progression of mesenchymal stem cells (MSCs). MSCs appear to be limited to follow an endochondral ossification program, which will end in vascular penetration, marrow deposition, and ossification of this cartilaginous tissue. Differentiation toward the stable hyaline cartilage phenotypes is theoretically restricted. However, it is possible that reproducing the complex environmental signals that are required for pathway switching has not been discovered. Bypassing these restrictions may be possible using reprogramming technology (induced pluripotent stem [iPS]) or embryonic stem cells (ESCs) as a new start state. Dashed arrows indicate possible routes, although evidence is lacking about the detailed differentiation pathway. Figure made with images available at Servier Medical Art (www.servier.fr). Color images available online at www.liebertpub.com/teb
The question is: do different cartilages develop from distinct chondroprogenitor subtypes, which are developmentally restricted, or are their molecular, morphological, and functional differences determined by the site-specific environment (e.g., load bearing), or both? Furthermore, are some of the components present during in vitro MSC chondrogenesis restricting their differentiation potential or is the end-stage phenotype restricted by developmental factors (e.g., cell history), which will eventually preclude their use as articular cartilage precursors? These unanswered questions strongly suggest that cells that differentiate from MSCs have a potential limitation of only being able to contribute to the endochondral ossification process (Fig. 2).
In contrast, it is possible that MSCs may retain developmental plasticity such that they could be guided toward the desired chondrogenic phenotype. To date, we and others have not been able to discover the required microenvironmental conditions in vitro needed to control this hypothesized plasticity. Supportive of the importance of the proposed environmental component during cartilage development is the fact that different types of cartilages are derived through different developmental routes (Fig. 1). For example, hyaline cartilage emerges both from the cranial neural-crest (e.g., nasal septal cartilage) and from lateral plate mesoderm (e.g., articular cartilage), and most likely, their distinctive features are acquired later due to anatomic site-specific and different physiological and physical signals. The hypothetical restricted potential of MSCs to differentiate toward a stable hyaline cartilage-like chondroprogenitor phenotype is illustrated in Figure 2. Alternately, we could hypothesize that this differentiation pathway can be achieved by specific microenvironmental signaling. If marrow MSCs are, indeed, restricted from molecularly differentiating toward the joint-type hyaline cartilage phenotype, it may be possible to bypass these restrictions using reprogramming technology or an embryonic stem cells (ESCs)-based approach (discussed in the “from pluripotency towards multipotency” section). Other complexity issues to manage to obtain site-specific stable cartilage from MSCs are the molecular and cellular differences exhibited by each anatomic cartilage. These different site-specific cartilages may require quite different environmental signals to generate one or the other cartilages (Fig. 1). We would argue that we are closer to knowing how to generate fibrocartilage from MSCs than articular cartilage. In vitro and in vivo studies have shown that the tissue elaborated by chondrogenically differentiated MSCs has features characteristic of fibrocartilage.53,54 Although this fibrocartilage is different from, for example, the one found in meniscus, Vangsness et al. recently reported on meniscus regeneration upon intra-articular injection of MSCs. 55
MSCs are heterogeneous
MSCs are an attractive source of cells for cartilage TE due to their ease of isolation and high capacity of in vitro expansion. MSCs have the potential to differentiate into chondrocytes, osteoblasts, myocytes, tendinocytes, ligamentocytes, and other mesodermal cell types. 56 Furthermore, they have immunoregulatory properties and can escape immune recognition while depressing immune mechanisms (reviewed in Ghannam et al. 57 ). However, what is currently termed an MSC, especially from bone marrow, is not a homogeneous population of cells, which may, in itself, affect our ability to obtain optimal reproducible results in cartilage TE.
The cells vary in number in the tissue, for example, in bone marrow, the frequency of MSC colony-forming units-fibroblast is on the order of 1 cell in 104–105 mononuclear cells. 58 However, numbers vary among donors, with donor age and health and with the tissue source. This becomes a key issue, as large numbers of cells are typically needed for TE, and side effects of in vitro expansion include significant phenotypic drift and rapid senescence.
Bone marrow and adipose tissue (termed ASCs, reflecting behavioral differences) are the main sources for therapeutic MSCs, with BM-MSCs being considered the gold standard source for musculoskeletal TE approaches. However, MSCs have been derived from a variety of tissues, including synovium, periosteum, dental pulp, umbilical cord vein, placenta, and others. 59 Many studies have shown the feasibility of MSCs from different tissue sources to generate cartilage.60–64 However, it is important to consider that MSC from those different sources are not necessarily equivalent. Indeed, they have shown significant functional differences,65,66 and most certainly require specific optimal chemical and temporal protocols for their chondrogenic differentiation.
Other progenitor cells with potential applications for cartilage TE are related to MSCs, for example, the multipotent adult progenitor cells or marrow-isolated adult multilineage inducible (MIAMI) cells. These cells have been described as pluripotent subpopulations within the bone marrow that can be isolated using complex cell culture methods, providing a new level of local heterogeneity.67,68 There is substantial donor-to-donor variability in the proliferation, differentiation potential, metabolic demands, and biosynthetic activity even with cell preparations obtained from a given tissue source.69,70 There is also intrinsic heterogeneity that results from different harvest sites or different cell differentiation stages, which affects (often reducing) the efficiency of the chondrogenic induction process. Heterogeneity of MSC cultures (even the in vivo heterogeneity) may dictate the differentiation behavior of distinct subsets of MSCs, thus reflecting the intrinsic capabilities of generating the appropriate cartilage matrix by the whole cell population. This may be the result of the in vivo microenvironmental histories at the source tissue that control their functions (pericytes vs. stem/progenitors), as we discuss below. In summary, MSCs from different anatomic locations, or even subpopulations within the same tissue, most likely have very dissimilar proliferation and differentiation potentials and differing requirements for, and responses to inductive stimuli, making it absolutely necessary to optimize culture conditions and differentiation protocols for each cell source, especially for the chondrogenic lineage. For emphasis, there are many different chemical and mechanical properties of cartilage throughout the body; it is not logical that MSCs could lineage-progress to fabricate these diverse cartilaginous tissues without unique and sequential signaling customized for each class of MSCs. For example, although TGF-β alone can induce marrow-derived MSCs, fat-derived MSCs require both TGF-β and bone morphogenetic protein-6. 71
Regarding pericytes as MSC progenitors, one possibility is the presence of cellular and chemical components in their perivascular niche that may have an inhibitory effect on the differentiation properties of MSCs. Thus, the purification of distinct subsets of MSCs may enhance their particular capabilities by eliminating interfering cells with limited potential, or even cells with inhibitory activities. We, and others, have shown that MSCs grown in the presence of fibroblast growth factor-2 (FGF-2) are specifically primed for subsequent chondrogenic differentiation and massive ECM formation; this effect may be related to a preselection of a subpopulation of cells that have increased chondrogenic potential. 72 We have unpublished data showing upregulation of Sox9 mRNA and protein in cells exposed to FGF-2. Thus, some of the intrinsic variability of MSCs can be reduced by growth factor treatment.73,74
New Dynamic and Developmental-Based Approaches for MSC Chondrogenic Differentiation
A review of current cartilage TE practice suggests that the most frequently used approach is to use a single culture medium formulation to isolate and expand MSCs, followed by a single chondrogenic medium formulation used to drive the entire differentiation process in vitro. This “fire and forget” approach to in vitro chondrogenesis, expected to carry differentiation from induction to implantation-ready tissue, has clearly reached its limits. This simple approach likely induces a very general (albeit unnatural) cartilage differentiation pathway in MSCs, resulting in their failure to form the highly specific hyaline cartilage phenotype. Moreover, the hyaline cartilage of the hip, knee, and ankle joints are all quite different, thus requiring a site-specific cartilaginous tissue. It is thought that the condensation step of endochondral bone formation is being mimicked as the initial step of MSC chondrogenic differentiation in micromass or pellet cultures, 75 but attempts to recapitulate embryonic development and lineage progression in in vitro chondrogenesis have been unsuccessful so far.
It seems likely that a more sophisticated approach will be needed, one based on an understanding of the molecular events involved in chondrogenesis induction, lineage progression, and maintenance of the chondrocyte phenotype. It also appears likely that a study of events during embryogenesis could be informative, in which the developmental phenomena can provide empirical basis for the design of MSC differentiation protocols. Moreover, most protocols that induce MSCs to differentiate into chondrocytes are based on static culture conditions that have been optimized in terms of morphogen concentration and cell density, which drastically differs from the ever-changing embryonic milieu; a true mimic methodology, a dynamic, changing microenvironmental approach is required. It seems likely that a choreographed temporal sequence of stimuli will be necessary to induce a functional and phenotypically stable chondrocytes and the formation of adult articular cartilage tissue.
Such methods will depend on how much of the data from cartilage developmental biology can be translated to an in vitro setting; insufficient information is currently available to allow the control of this intricate and dynamic process that would result in the generation of a stratified hyaline cartilage tissue. 76 Restoration of the zonal organization of native articular cartilage is now considered by some to be critical for the success of the cartilage TE approaches.17,20 There are enormous differences in the zonal organization and structure of human articular cartilage, depending on the different microenvironmental factors to which the cells are exposed and the cartilage zone where they reside. Therefore, an engineered construct should have a strict localization-based design, which considers the importance of chondrocyte properties, such as cell physiology, morphologies, orientation, and biosynthetic activities.17,77,78 It seems reasonable to suggest that location-specific chondrocytes would be optimal to reconstruct a specific cartilage zone. 78 This requirement includes a new challenge to MSC-based TE cartilage in which subtle differences in cell properties found within the different layers of the cartilage should be mimicked to recreate a functional stratification. Attempts have been made to use different chondrocyte populations to mimic cartilage regional cellular variation.78,79 Using a multilayered hydrogel system, and a method to isolate chondrocytes from three different layers of articular cartilage, Kim et al. showed that recreation of the stratification of native articular cartilage is possible. 78 This method could be useful for an MSC-based technology; for example, using multilayered constructs in combination with multiple subpopulation of MSCs (even MSCs from different tissue sources). In this regard, it has been suggested that MSCs derived from synovium, infrapatellar fat pad, and muscle are promising cell sources for creating a superficial zone of articular cartilage by TE, since they can be induced to secrete SZP.80–82 As SZP plays a critical role in articular cartilage physiology, the localization of SZP-secreting cells at the surface of a TE cartilage construct may be critical for a successful biomimetic approach. 83 In this regard, Lee et al. demonstrated that MSCs isolated from synovium and infrapatellar fat pad produce SZP after chondrogenic induction and can maintain this activity after passaging. Therefore, these MSCs could be used as a potential source for superficial zone cartilage TE, allowing controlled localization of SZP. 81
Overall, although it may be possible to provide culture conditions that are complex enough to, in part, recreate the composition of the in vivo milieu at a given time, the dynamic complexity needed to recreate the events during articular cartilage development is far from being understood. As we gradually incorporate new data into our understanding of cartilage development, the site-specific efficiency of the biomimetic process can be perfected. We still need to understand how microenvironmental cues, such as soluble factors, ECM molecules, physical stimuli, and cell–cell and cell–ECM interactions, control the lineage progression and the eventual stem cell fate.
Sequential addition or removal of growth factors are already being considered in an attempt to exploit cascade effects, for example, exposure to a first set of culture conditions alters the cell's receptor profile priming the cells for exposure to a second or subsequent set of conditions.84,85 We have found that a sequential exposure of hMSCs to members of the FGF family of morphogens has a significant cascade effect on cell expansion, chondrogenic differentiation, and hypertrophic differentiation, allowing for control of the chondrogenic differentiation program (unpublished data). The persistent effect of FGF-2 exposure in the expansion medium on chondrogenesis, even after withdrawal of the FGF-2, is one example of a cascade effect that crosses over between expansion and differentiation conditions. 74 Finally, combining bioactive factors with an instructive scaffold may be required.
Cell Optimization
In the marrow space, MSCs can be at various stages of differentiation, or even adopting different transient phenotypes depending on their physical location and function.86,87 It is currently clear that the typical, in vitro starting population of isolated MSCs is highly heterogeneous. Overcoming this heterogeneity may be a useful way of optimizing the cellular starting material for TE. Using specific cell surface markers, characteristic of specific and different phenotypes, it may be possible to obtain enriched populations with desirable traits using a cell-sorting approach. Thus, the cells could be presorted according to their differentiation potential before beginning dynamic chondrogenic induction. Several possible sorting strategies are emerging.
Our unpublished data and a recent report by Gharibi and Hughes suggest that FGF-2 stimulation during cell expansion alters the distribution of CD146+ cells suggesting that CD146 could serve as a sorting criterion to select cells with maximized chondrogenic potential. 88 Rada et al. showed that CD105+ and CD29+ MSCs have greater chondrogenic potential over other MSC subpopulations. 89 Moreover, Arufe et al. found that the CD271+ subpopulation expressed the highest level of type II collagen and aggrecan at 28 days of chondrogenic induction compared with other MSC subpopulations. 60 Moreover, a recent report showed that CD271+ MSCs have greater chondrogenic potential, both in vitro and in vivo, compared with the expanded MSC culture. 90
All these findings confirm that hMSC derived from different tissue sources constitutes a heterogeneous cell population comprising several subpopulations, each one having particular phenotypic and functional characteristics. The distinctive phenotypes may parallel the stage of differentiation and, more importantly, determine functional aspects related to the cells' physical location in the bone marrow and their ultimate functional fate. Collectively, this information supports the use of selection protocols that query the actual cellular phenotype before their use as cartilage matrix building units.
Hybrid Cultures
The study of the molecular control of in vitro chondrogenesis relies on the addition of purified or recombinant proteins to the culture medium. However, this mode of delivery guarantees inhomogeneous penetration and distribution of these additives within the growing cell aggregate. Additionally, the effects of cell-to-cell communication during differentiation events, while likely critical, are not fully understood. MSC-adult hyaline chondrocyte coculture has been shown to provide chondro-instructive and other signals both by a trophic effect and by direct cell-to-cell contact. These interactions bidirectionally modulate the biology and phenotype of both cell types: chondrocytes proliferate and maintain a differentiated phenotype with enhanced matrix synthesis, whereas MSCs are pushed into the chondrogenic pathway. 91 Interestingly, the trophic effect exerted by MSCs seems to be independent of the tissue source for the cells. 92 Coculture-mediated chondroinduction is also influenced by cellular interaction with the ECM via integrins and by cell-to-cell contact via gap junctions or transmembrane proteins, such as cadherins. 93
Using different coculture systems, various groups have identified intercellular communication through soluble diffusible factors that promote chondrogenic differentiation of MSCs and enhance the chondrogenic potential of dedifferentiated chondrocytes. The newly differentiated chondrocytes may further induce chondrogenic differentiation of resident MSCs, resulting in the formation of more chondrocytes. 93 These systems include cartilage explants, 45 freshly isolated chondrocytes, 94 and chondrocyte-conditioned medium, 95 as the source of the bioactive factors. Thus, Ahmed et al. investigated phenotypic stability of MSCs in a coculture study where the presence of rat cartilage pieces suppressed type X collagen gene expression in rat MSCs. 45 Similarly, Giovannini et al. showed that MSCs cocultured with articular chondrocytes, but in the absence of dexamethasone and TGF-β1, did not express hypertrophy markers even after 6 weeks. Interestingly, in the presence of dexamethasone and TGF-β1, articular chondrocytes failed to downregulate hypertrophy markers. 46 Bian et al. showed that hMSCs and human chondrocytes, cocultured at a ratio of 4:1 and encapsulated in hyaluronic acid hydrogels, decreased their expression of cell hypertrophy markers, such as type X collagen. 96 Another study provided evidence that chondrocyte-secreted PTHrP mediated an antihypertrophic effect in MSCs. 97
Despite the fact that coculture methods have been developed to study in vitro chondrogenesis, the mechanisms underlying these cellular interactions are only beginning to be understood, in part, due to technical difficulties related to determining the specific relation between the acting factors and the observed response. It is, in particular, not always clear which direction of interaction between MSCs and chondrocytes dominates and how the cell-type ratios influence chondrocyte function and chondrogenic differentiation of MSCs. Although these phenomena are understudied, they appear important for future improvement of cartilage TE strategies.
In Search of the True Articular Hyaline Cartilage Progenitor
As stated above, it is not clear that bone marrow is the ideal source of MSCs for cartilage repair. This is rooted in the intrinsic endochondral ossification potential of BM-MSCs, which results in an undesirable cellular phenotype (hypertrophic) that is not useful for articular cartilage repair. Therefore, despite the many theoretical advantages of MSCs, the search for a true cartilage stem or progenitor cell continues. Such a progenitor would be useful for understanding the molecular and genetic signature that a true hyaline cartilage progenitor should possess. Furthermore, if current techniques allow us to isolate and expand these cells, it would then be possible to design proper cell therapy approaches based on cells that have well-defined intrinsic properties. However, the identity and physical location of this progenitor cell remains a subject of study.
Adult intra-articular tissues (cartilage, synovium, etc.) provide a potential reservoir of chondroprogenitors. There is strong evidence of a cartilage stem cell in the superficial layers of hyaline cartilage.98,99 The superficial layer of the cartilage is the first that is lost in degenerative joint disease, which has implications for understanding the progression of the disease, but unfortunately, it also curtails the supply of regenerative cells for research. The deep zones of cartilage have also been shown to be a possible reservoir of chondroprogenitor cells, based on identification of cells that have proliferative capacity and can rebuild cartilage tissue. 100 These studies have shown that isolation of native cartilage progenitors can be accomplished by a combination of techniques, which relies on differential progenitor properties, such as colony-forming capabilities, differential adhesion properties, and specific marker expression.98,101 Recently, equine chondroprogenitors from the superficial zone have been isolated and compared with BM-MSCs, showing that these superficial cells have superior capabilities for cartilage repair, principally in terms of lacking the expression of hypertrophic markers, such as Runx-2 and type X collagen. 49 The presence of chondroprogenitor-like cells has also been described in other locations within the joint, such as the perichondrium,102,103 synovium,60,64,104 perichondrial groove of Ranvier, 105 and infrapatellar fat pad.106–108 All these cells resemble MSCs, suggesting that these locations could be reservoirs of MSC-like cells, which may have useful physiological functions within the joint.
From Pluripotency (ES, Induced Pluripotent Stem Cells) Toward Multipotency
The reason that MSCs from different sources do not make articular hyaline cartilage has not been fully addressed. One explanation could be that the developmental origins of adult MSCs and articular chondrocytes are different. It is possible that authentic articular cartilage chondroprogenitors are only transiently present during development. 109 Recently, Wu et al. described early human cartilage committed cells. They used laser-capture microdissection and microarray analysis of human limbs from 5 to 6 weeks of development to identify cells that have a very specific molecular profile, not found in chondrocytes at later stages. 109 These data are in agreement with the fact that formative chondrogenesis is largely completed before birth. 110 The recent proposition of a neural crest origin for BM-MSCs suggests that the observed chondrogenic potential of these cells might be closer to a craniofacial cartilage. This is structurally and functionally different from the limb joint hyaline cartilage resulting from the embryonic mesodermal chondroprogenitors (Fig. 1).111,112 The developmental restrictions that MSCs and other adult progenitor cells may have to generate a functionally hyaline cartilage may be overcome using less compromised stem cells. Chondrogenesis from pluripotent stem cells, such as ESCs and induced pluripotent stem cells (iPSCs), has been explored extensively. Several protocols have been developed to induce chondrogenic differentiation from ESCs and iPSCs.113–117
The relative success rate in obtaining the correct or native chondrocyte and ECM phenotype using ESCs seems to be greater compared with MSC-based protocols. However, these conclusions depend on how the final outcome is assessed. It seems probable that the final chondrocyte phenotype obtained from ESCs depends more on the culture system rather than on the cell source (embryoid bodies, purified ESCs, ESC-derived MSCs).118–120 Although ESCs as a cell source for cartilage repair have been shown to be promising, their clinical application remains controversial due to ethical concerns related to their isolation, along with a risk of teratoma formation and tissue rejection. 121 This makes application to human therapy unlikely in the short term.
Genetic reprogramming of adult somatic cells (e.g., dermal fibroblast) to restore multidifferentiation capacity is possible by overexpressing transcription factors specifically associated with pluripotency, such as Oct4, Klf4, c-Myc, and Sox2. 122 These iPSCs are emerging as a cell source for cartilage TE, as these cells have the same potentials as ESCs and lack some of their shortcomings. Numerous in vitro approaches have been developed to generate chondrogenic cells from iPSCs based on protocols previously established for ESC differentiation as well as novel approaches.123–125
Interestingly, some of these studies have found that an initial differentiation of ESCs or iPSCs toward an intermediate phenotype, not necessarily an adult MSC-like one, and only then toward chondrogenic differentiation appears to be an efficient approach to generate articular-like chondrocytes.124–126 These observations suggest that the mesodermal stem cell-like phenotype is a crucial stage in cartilage differentiation. It is possible that different developmental potentials and restrictions (or immature intermediate stages) exist within these mesodermal lineages (e.g., in embryonic MSCs vs. adult MSCs).
Protocols for the generation of MSC-like cells from pluripotent cells usually do not describe whether the cells are of mesodermal or neural crest origin, thus these cultures may be composed of several different types of cells. 127 It is important to stress that these progenitors are not comparable to adult hMSCs. Importantly, these embryonic progenitors respond to specific morphogens in ways that adult hMSCs cannot. Indeed an “adult” hMSC has yet to be derived from iPS technology.
Recent studies have shown that somatic stem cells may be more suitable than adult differentiated cells for genetic and nongenetic reprogramming, given that they need fewer reprogramming factors and that the efficiency of iPSC generation is higher. For instance, mouse BM-MSCs have been successfully used to create iPSC lines.128,129 Kunisato et al. showed that iPSC generation from BM-MSCs is more efficient compared with mouse epidermal fibroblasts, whereas Niibe et al. showed that purified BM-MSC cultures generate high-quality iPSCs.129–131 If iPSCs generated from MSCs are to be the biological source of new cartilage, then we must understand and control the intrinsic properties of the initial population to be reprogrammed to an iPSC, that is, we must start from a homogenous and well-characterized cell population. Additionally, the differentiation protocol has to be specified so that iPSCs can generate functional chondrocytes that can coordinate the formation of site-appropriate (i.e., ankle vs. knee articular cartilage) ECM.
Despite the enormous potential of both ESCs and iPSCs, their clinical use is still subject to our ability to control their phenotype. This is related not only to their final phenotype (articular cartilage) but also to the risk of teratoma formation and immune rejection. 132 Thus, although these cells have a great potential to be used for cartilage regeneration, purification and differentiation protocols should be even more highly controlled than for adult stem cells. To circumvent these difficulties, it could be useful to partially reprogram somatic cells toward an intermediate state (i.e., chondrogenic committed cells), to avoid the generation of fully pluripotent cells.
A more direct approach has been developed in which chondrogenic cells were generated directly from somatic cells without producing iPSCs. For this, the authors used a combination of reprogramming factors (c-Myc and Klf4) with a chondrogenic factor (Sox9) resulting in chondrogenic cells that formed histologically homogenous hyaline cartilage.133,134 The authors suggested that c-Myc and Klf4 are involved in epigenetic events in the somatic cells, enabling Sox9 to direct cells to the chondrogenic lineage during the induction of the reprogrammed cells. Moreover, the generated cells were nontumorigenic and have stable karyotypes. Overall, these technologies could contribute to the development of efficient approaches for site-specific articular cartilage regeneration. However, we must not ignore the fact that articular cartilage is not a final, homogeneous, and definitive cellular phenotype; rather cartilage is an architectural phenomenon. Thus, obtaining the chondroprogenitor does not guarantee the successful formation of stratified cartilage necessary for a clinical application.
MSC/Pericytes
The concept that a pericyte, a cell embedded within the abluminal basement membrane of blood vessels, 135 possesses the potential to give rise to cells of multiple lineages was first introduced almost two decades ago.136–138 Moreover, in 1965, Urist made the striking observation that a young perivascular connective-tissue cell responded to soluble signals coming from cells invading decalcified matrix (free macrophages), inducing them to differentiate into osteoprogenitors. Furthermore, in a different microenvironmental setting (compaction of cells in closed vascular channel), the same cells could also give rise to chondroprogenitor cells. 139 Besides the known functions of pericytes, the suggestion that they may be resting stem/progenitors cells participating in neoangiogenesis and wound healing was apparent almost 30 years ago. 135 Diefenderfer and Brighton reported data based on molecular analysis that pericytes, besides their osteogenic role, also had a chondrogenic nature based on the presence of aggrecan and type II collagen mRNA. 140 Farrington-Rock et al. later showed that microvasculature-derived pericytes could differentiate into chondrocytes in vitro and in vivo. They, thus, demonstrated for the first time that pericytes express the master chondrogenic transcription factor Sox9, supporting the hypothesis that pericytes may serve as a reservoir of primitive precursor cells that can contribute to neochondrogenesis. 141
It has been suggested, and subsequently shown, that MSCs present in bone marrow and many other organs are a subset of perivascular cells in vivo.140,142–146 The potential of pericytes to differentiate toward mesenchymal phenotypes may be restricted by the perivascular microenvironment; they may become permissive when pericytes exit this restrictive microvasculature ECM and become MSCs based on the microenvironmental conditions they experience in vivo.145,147 How these events are related to articular cartilage repair/regeneration is not known, and whether pericytes directly contribute to chondro-dynamics is still an open question. Moreover, their function may be different in developing tissues compared to their role in adult tissues, taking into consideration that it is unlikely that an adult tissue milieu would provide the appropriate cues to allow a regenerative response compared to a reparative one. It can be envisioned, based on the events during osteochondral development, that unique anatomical locations, together with highly controlled local microenvironmental cues (e.g., matrix and cellular interactions, soluble factors, oxygen tension) determine the precise differentiation phenotype that the MSC/pericyte can acquire in vivo. It has not yet been possible to mimic these conditions reliably in vitro or in different TE and Regenerative Medicine approaches.
Conclusions
The main challenges that remain with MSC-based articular cartilage TE, such as quality and durability of the de novo generated tissue, its resistance to an endochondral ossification program, and its effective integration into the anatomic-specific host tissue, may be due to the intrinsic inability of MSCs to differentiate into articular cartilage or may be a reflection of our poor understanding of the underlying MSC biology. MSCs can be isolated from many human vascularized tissues as they reside as perivascular cells, including various locations within the joint that may be considered as potential reservoir of chondroprogenitor cells. Until now exogenously administered as well as endogenously mobilized MSCs have failed to contribute to durable and phenotypically correct (i.e., anatomic site-specific) articular cartilage regeneration, observations that might cast doubt on the usefulness of MSCs in regeneration of hyaline articular cartilage. This apparent lack in potential of MSCs to give rise to stable cartilage may come from restrictions imposed during embryonic development. Therefore, it is possible that their functions and responses in developing tissues are quite different from adult tissues. ESC and iPSC technology have provided evidence that these restrictions may be circumvented, which would allow us to obtain cells with an intrinsic potential to regenerate site-specific articular cartilage.
Whether adult marrow MSCs are suitable for such site-specific articular cartilage TE or have an intrinsic endochondral bone formation program is still an open question, but the latter seems likely. Moreover, whether adult MSCs have lost their intrinsic ability to differentiate into articular cartilage or whether we are not currently able to recreate the developmental milieu to which they are exposed during embryology are also unanswered and challenging questions. We suggest that, at the very least, regenerating stable articular cartilage using MSCs will require us to implement more rigorous methodologies to precisely mimic morphogenetic events of embryonic development of cartilage, yet note that more information as to how this process successfully guides stem cells to differentiate into a cartilage phenotype is still needed.
Footnotes
Acknowledgments
This work was supported in part by the L. David and E. Virginia Baldwin Foundation, and through a grant from the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health (P01 AR053622). We thank anonymous reviewers of the initial submission for their helpful suggestions, which have in part been incorporated verbatim into this revised version. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Disclosure Statement
No competing financial interests exist.