
Abstract
Tissue-engineered grafts for cardiovascular structures experience biochemical stimuli and mechanical forces that influence tissue development after implantation such as the immunological response, oxidative stress, hemodynamic shear stress, and mechanical strain. Endothelial cells are a cell source of major interest in vascular tissue engineering because of their ability to form a luminal antithrombotic monolayer. In addition, through their ability to undergo endothelial to mesenchymal transition (EndMT), endothelial cells may yield a cell type capable of increased production and remodeling of the extracellular matrix (ECM). ECM is of major importance to the mechanical function of all cardiovascular structures. Tissue engineering approaches may employ EndMT to recapitulate, in part, the embryonic development of cardiovascular structures. Improved understanding of how the environment of an implanted graft could influence EndMT in endothelial cells may lead to novel tissue engineering strategies. This review presents an overview of biochemical and mechanical stimuli capable of influencing EndMT, discusses the influence of these stimuli as found in the direct environment of cardiovascular grafts, and discusses approaches to employ EndMT in tissue-engineered constructs.
Introduction
T
EndMT plays an important role in embryonic development and remodeling of adult heart valves. 1 During embryonic development, semilunar valves emerge from endocardial cushions that bulge into the cardiac outflow tract. This happens in response to a tightly regulated combination of growth factors, microRNAs, matrix proteins, and mechanical stimuli 2 that will be discussed further in this review. As they undergo EndMT, endothelial cells lining the cushions migrate inward from the endothelial monolayer into the cardiac jelly, becoming a matrix-producing and matrix-remodeling cell type with myofibroblast-like characteristics. 2 These cells, which are responsible for producing and maintaining the interstitial cardiovascular tissue, will continue to populate the adult heart valve and are responsible for life-long remodeling and mechanical functioning of the valve leaflets. 3
In addition, EndMT plays a role in the response of the adult vasculature to hypoxia or injury. In addition to hypoxia-inducible factor 1 alpha (HIF1α)-induced proliferation, migration, and adhesion of endothelial cells,4,5 EndMT enables the remodeling and expansion of the existing vessel network through transdifferentiation of the endothelial cells into smooth muscle cells and pericytes in the newly sprouted vessels.2,4 In this process, EndMT is triggered by growth factors, microRNAs, matrix proteins, and mechanical stimuli, and involves signaling factors that are highly similar to developing heart valves. These factors, which will be discussed further in this review, may be applied in a stepwise approach to vascular tissue engineering for both heart valve replacement grafts and vascular replacement grafts (Fig. 1). While replacement heart valves and blood vessels are distinct structures that will require separate approaches, both are dependent on an endothelial monolayer and a population of ECM-producing cells. EndMT, therefore, is of great interest to tissue engineering strategies for both heart valve replacement grafts and vascular replacement grafts.
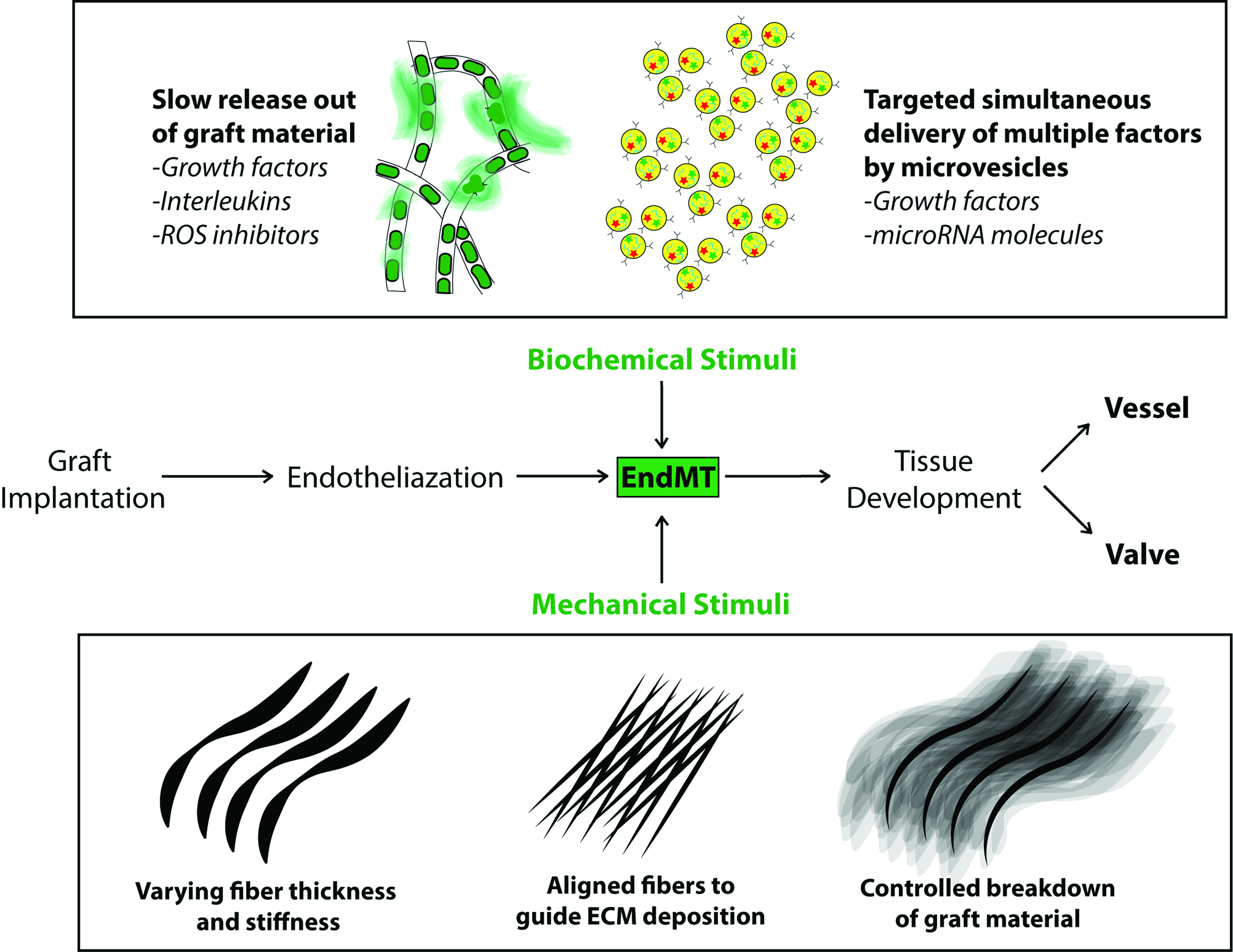
Endothelialized cardiovascular tissue-engineered grafts experience environmental stimuli influencing tissue development. To influence this environment, local delivery of biochemical stimuli such as growth factors, interleukins or ROS inhibitors by slow-release mechanisms, or simultaneous delivery of biochemical factors and microRNA molecules by microvesicles can help guide tissue formation. Additionally, the mechanical properties of the graft material itself can help guide cellular development by locally varying material stiffness, fiber alignment, and the controlled breakdown of fibers allowing the newly formed tissue to take over the mechanical load. ROS, reactive oxygen species. Color images available online at www.liebertpub.com/teb
Heart valve or vascular graft scaffold material may be preseeded with endothelial cells or cell-free material may be designed to self-endothelialize with host-derived cells after implantation.2,6 Following implantation, endothelial cells can be stimulated to undergo EndMT to give rise to an autologous cell population that produces and maintains sufficient ECM to achieve functional tissue. It should also be taken into account that accumulation of ECM and, in particular, collagens may be associated with fibrosis in vivo.3,7 Fibrosis can potentially lead to stenosis and subsequent graft failure. The stiffness of fibrotic material differs from that of healthy tissue and can impair tissue function. 8 A mechanically sturdy tissue, however, is required for mechanically loaded replacement grafts. A balance, therefore, between EndMT induction and inhibition, is necessary to harness EndMT for tissue engineering.
Understanding how the environment regulates EndMT in endothelial cells is crucial and studies should focus on appropriate cell sources. Endothelial cells from a wide variety of sources have been investigated in this context, such as human umbilical arterial endothelial cells (HUAECs), human umbilical vein endothelial cells (HUVECs), and endothelial colony-forming cells (ECFCs) (see Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/teb). In particular, ECFCs represent an easily accessible candidate cell source for preseeded tissue-engineered constructs as they can be cultured out of the peripheral blood and can produce large colonies of monolayered endothelial cells. 9 In addition, ECFCs are considered a model cell type for circulating endothelial progenitor cells. 10 Moreover, ECFC have been shown to be capable of undergoing EndMT. 11
This review aims to present an overview of the influence of biochemical stimuli and mechanical forces in the direct environment of vascular and valvular grafts on EndMT, and discuss approaches to employ EndMT in tissue-engineered constructs for cardiovascular application.
Mechanisms of EndMT
The capacity of endothelial cells to undergo transition into a mesenchymal cell type resembles that of epithelial cells, which can undergo epithelial to mesenchymal transition (EMT). EMT plays a role in embryonic development and wound healing, but also organ fibrosis and its pathways are well studied. 12 EMT and EndMT have many regulatory pathways in common, and both yield a cell type capable of producing and remodeling ECM. EndMT in cardiovascular development comprises migration and the production and remodeling of ECM. Endothelial cells that originally form a monolayer reduce their adhesion to the surrounding cells and develop the ability to migrate.13–15 As cells begin to migrate away from a monolayer, their cortical cytoskeleton is rearranged to enable cell motility by developing actin-rich projections in sheet-shaped lamellipodia or spike-shaped filopodia.16–19 Loss of endothelial markers (vascular endothelial cadherin [VE-Cadherin]; platelet endothelial cell adhesion molecule [PECAM-1, or CD31]) corresponds to loss of cell–cell adhesion, migration, and marks endothelial cell dedifferentiation. As cells move away from the monolayer and deeper into the ECM-rich environment they develop the ability to increase production and remodeling of ECM2,20 Increased expression of collagens and elastin, but also ECM-degrading enzymes, such as matrix metalloproteinase 2 (MMP2) and MMP9, are important and functional indicators of EndMT. The gain of myofibroblast markers (fibroblast-specific protein 1; FSP1, alpha smooth muscle actin; α-SMA, collagen type 1) corresponds to a newly acquired ability to produce and remodel ECM. Intracellular signaling proteins, such as Snail, Twist, Slug, and zinc finger E-box-binding homeobox 1 (ZEB1),5,21 are important regulators of EndMT and are commonly used as markers in EndMT research. The pathways that induce EndMT converge on several signaling proteins, including Snail, which downregulates the expression of proteins involved in maintaining cell–cell adhesion such as integrins and focal adhesion kinases and upregulates proteins involved in migration and ECM production and remodeling such as collagen type 1, elastin, and MMPs.4,22–24 Besides Snail, EndMT also requires a downregulation of GSK3β to decrease Snail degradation, which can be achieved by transforming growth factor β2 (TGFβ2)-induced PI3K signaling. 14 This is different from EMT where Snail alone is sufficient for induction. 25 Factors such as TGFβ2, therefore, are of major interest to tissue engineering approaches focused on EndMT.
Biochemical Factors Regulating EndMT
TGFβ is a major regulator of EndMT
The major inducers of EndMT are members of the TGFβ super family, which, among others, contains bone morphogenic proteins (BMPs), growth and differentiation factors, and three mammalian types of TGFβ: TGFβ1, TGFβ2, and TGFβ3. 26 The crucial role of TGFβs in cardiovascular development is apparent from knockdown experiments. Loss of TGFβ115 and 220 results in severe malformation of vasculature and absence of cellular migration into the cushions of the developing heart valves17–19 due to inhibition of EndMT, underlining its role in valvular development. TGFβ3, rather than inducing EndMT in cardiac development, appears to play a role in EndMT-related processes in lung tissue. 27 Endothelium-specific knockdown of TGFβ receptors has been shown to specifically inhibit the migration of mesenchymal cells into the cardiac cushions. 28 Another clinical example demonstrating the importance of TGFβ-signaling is Loeys–Dietz syndrome, 29 which is caused by autosomal dominant mutations in SMA and MAD 3 protein (SMAD3) and TGFβ receptor-encoding genes. Dysfunctional TGFβ receptors prevent EndMT leading to reduced structural integrity of connective tissue and a higher chance of developing aortic aneurysms.30,31 There are two types of TGFβ receptors, TGFβR type I (activin receptor-like kinase 5 (ALK5) and ALK1) and TGFβR type II, which upon ligand binding, combine to form a heterotetrameric receptor complex and phosphorylate receptor-regulated SMAD proteins (R-SMADs). In addition, binding to coreceptors endoglin and beta-glycan can further modulate the response to TGFβ. Downstream of TGFβ signaling, phosphorylated R-SMADs will form a complex with the co-SMAD SMAD4 and move into the nucleus where they bind DNA and regulate the transcription of many of the TGFβ target genes. 32 The signaling between TGFβ receptors and SMAD proteins is elaborate and enables a fine-tuning of the cellular response. 33 This indicates that TGFβ signaling is regulated by multiple pathways that may be targeted to control EndMT. For example, a high level of TGFβ will mainly regulate transcription by signaling through ALK5, whereas low levels of TGFβ will bind to a complex of two type I receptors, ALK5 and ALK1. Therefore, high levels of TGFβ will lead to the phosphorylation of Smad2 and Smad3 by ALK5, which will induce EndMT. 4 Low levels of TGFβ, however, will activate Smad1, Smad5, and/or Smad8 through ALK1. Within the heterotetrameric complex, the ALK1 kinase will inhibit ALK5 signaling and, therefore, inhibit EndMT. 4 Combined, this indicates that including the timed release of either high or low doses of TGFβ in scaffold materials may allow for increased control over EndMT. Synthetic materials for vascular grafts that are capable of controlled local release of factors have been described and would allow a single material to release both high and low doses of TGFβ with control over the timing of release. 34 This material has previously been used to enhance the development of tissue-engineered small-diameter vessels by timed release of vascular endothelial growth factor and platelet-derived growth factor. 35 An alternative method to deliver multiple factors simultaneously may be found in extracellular vesicles. Extracellular vesicles are lipid membrane vesicles that are involved in intercellular communication, 36 including TGFβ signaling. 37 The formation of extracellular vesicles occurs in the endocytotic pathway when intraluminal vesicles are formed in the late endosome, thereby forming the multivesicular body.36,38,39 Importantly, this means that the contents of extracellular vesicles are dependent on the cytosolic contents of the cell. The physiological role of extracellular vesicles is intercellular communication either by direct interaction with cell surface receptors, by fusing with the target cell membrane, or by endocytosis by target cells. Extracellular vesicles have been shown to be able to present TGFβ on their surface and effectively deliver it to fibroblast to trigger differentiation into myofibroblasts.37,40 The contents of extracellular vesicles can be manipulated.41,42 To optimize extracellular vesicles for tissue engineering approaches they could be harvested from cells cultured under specific conditions such as hypoxia, which increases the content of ECM-remodeling proteins in extracellular vesicles. 41
MicroRNAs can modulate EndMT by targeting ECM expression
MicroRNAs (mirs) are small noncoding RNA molecules that strongly bind to the 3′ UTR of mRNA and inhibit their translation. Many mirs have been reported to be involved in the induction or inhibition of EndMT. Mir23, for example, inhibits TGFβ-induced EndMT in the embryonic heart and restricts cardiac cushion formation. 43 Overexpression of Mir155 inhibits EndMT by targeting Ras homolog gene family member A (RhoA). 44 In contrast, mir21 overexpression induces EndMT. 45 Multiple cellular programs that play a role in EndMT are regulated by mirs. For instance, mir29a, mir29b, and mir29c target the expression of collagen type 1 and elastin and are lowered by TGFβ signaling. 46 Mir29b (along with mir195) plays a role in the development of aortic aneurysms through regulation of ECM expression. 47 Importantly, TGFβ1 only appears to lower the expression of mir29b, and thus will not lead to a complete silencing of these mirs. 48 TGFβ2, however, has been shown to lower all three of these mirs and lead to an increased expression of collagen type 1 and elastin even in haploinsufficient fibroblasts. 49 This means that TGFβ2 may be more effective in the targeting of mirs to manipulate ECM production. The ability of mirs to fine-tune ECM expression may be of value in developing implanted grafts (Fig. 1). Determining which mirs guide the ECM reconstitution of native vascular structures will point out novel candidates. Either mirs (e.g., mir21) or mir inhibitors (e.g., against mir29) may be incorporated in delivery vehicles and released as the graft cells undergo EndMT, and guide the development of the ECM to avoid a predominantly fibrotic development and instead lead to a mixed ECM composition that includes elastin and resembles native vasculature. Here, too, extracellular vesicles are a potential delivery vehicle, capable of delivering mirs in addition to growth factors. Extracellular vesicles from a variety of cell sources were shown to contain mir-29, mir-192, and Let-7 mirs which are all implicated in EMT-mediated metastasis.50–53 Let-7 also plays a major role in fibroblast growth factor (FGF)-induced EndMT. Disruption of FGF reduces Let-7 expression and leads to an increase of TGFβ ligands and receptors, ultimately resulting in EndMT. 54 Therefore, extracellular vesicles containing mirs such as Let-7 may target multiple mechanisms driving EndMT. Control over the mirs within extracellular vesicles can be achieved by using overexpression vectors applied to the cells producing the vesicles.55,56 In addition to enriching extracellular vesicles it is possible to reduce the presence of specific mirs by transfecting cells with antisense single RNA molecules. 57 The possibility to tailor the contents of extracellular vesicles makes them a versatile tool for cardiovascular tissue engineering.
Inflammatory signaling and oxidative stress promote EndMT
Implantation of a replacement vascular graft will induce an immunological response. 58 The resulting inflammatory environment will contain a myriad of signaling proteins such as tumor necrosis factor α (TNFα), interferon gamma (IFNγ), TGFβ, interleukins (ILs), and reactive oxygen species (ROS). 6 The factors released into this environment have individual effects on EndMT and the combination of factors will dictate the net outcome. To avoid adverse effects of the inflammatory environment on EndMT, grafts may be designed to minimize the impact of inflammation. For example, oxidative stress increases the expression and secretion of TGFβ1 and TGFβ2, 59 which could induce EndMT in the implanted graft (Fig. 2). Montorfano et al. demonstrated that H2O2 can induce the conversion of endothelial cells into myofibroblasts, shown by decreased expression of the endothelial markers VE-cadherin and CD31, and increased levels of fibrotic (FSP1) and ECM proteins (fibronectin). 59 Graft material may incorporate factors to reduce ROS, thereby limiting EndMT in a timely manner. ROS inhibitors, such as astaxanthin, were shown to reduce fibrotic development through SMAD/TGFβ signaling. 60 The intended vascular structure may also be designed with regard to the expected oxygen tension in situ. Hypoxia has been shown to induce Snail expression through HIF1α in mice. 61 With regard to the design of heart valve replacement grafts, the influence of oxygen tension may be highly relevant, considering that in vivo a pulmonary valve experiences a different oxygen tension than an aortic valve. 62
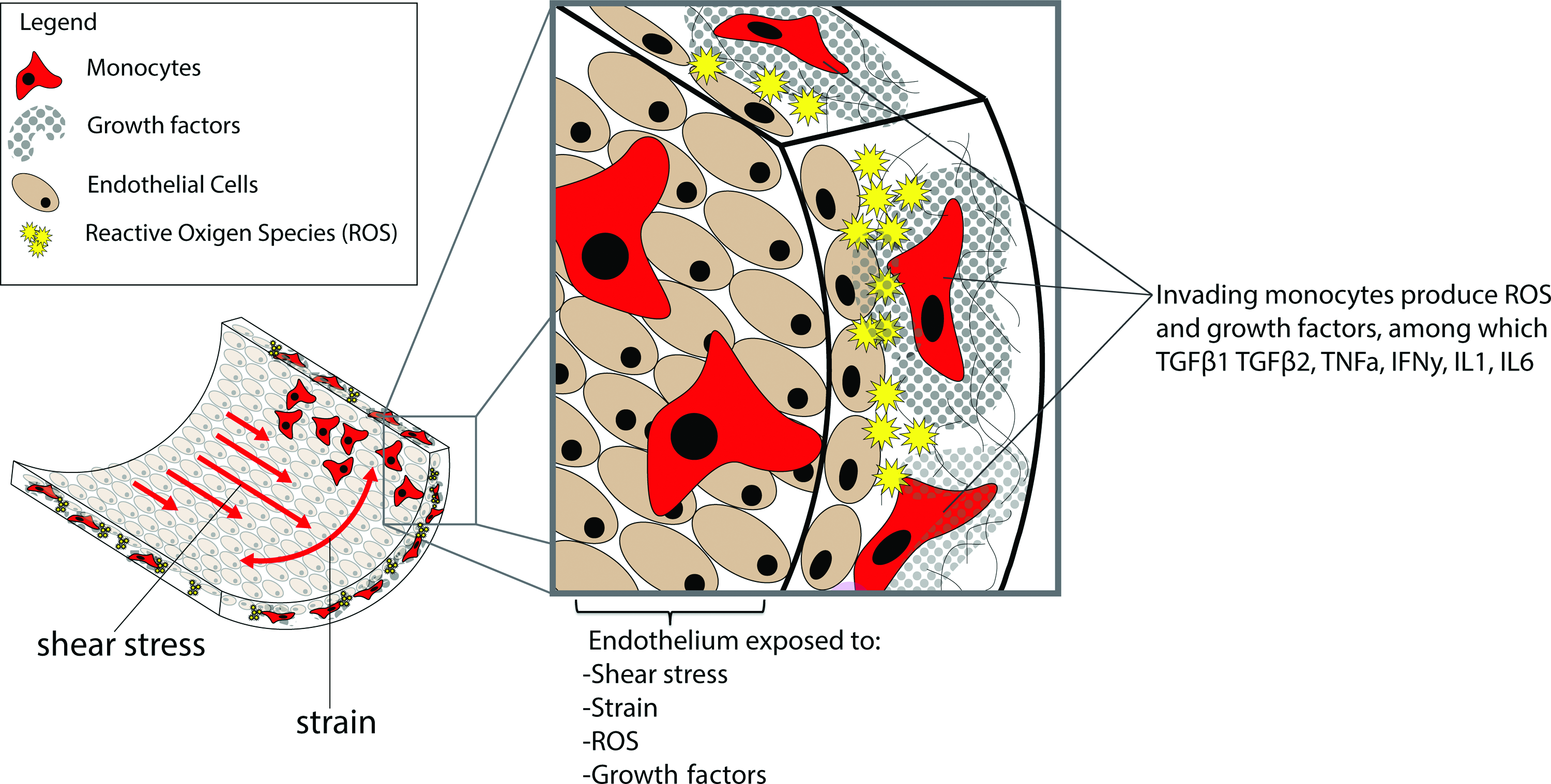
Overview of stimuli influencing EndMT in implanted cardiovascular grafts. Shear stress and strain may enhance EndMT depending on the local composition of growth factors that the endothelium is exposed to. This composition, in turn, is dependent on the invasion of inflammatory cells, ROS, and inflammatory signaling. The sum of the inhibitory and stimulatory effects will determine if EndMT will proceed. EndMT, endothelial to mesenchymal transition. Color images available online at www.liebertpub.com/teb
The implantation of graft material will likely trigger a foreign body response mediated largely through ILs. Mahler et al. have shown that an inflammatory signaling environment induces EndMT through IL-6 and TNFα through TGFβ signaling. 63 Further supporting the influence of immune signaling in EndMT, in the context of chronic inflammatory bowel disease, TGFβ, IL-1, and TNFα were shown to induce EndMT in microvessels contributing to intestinal fibrosis. 64 Maleszewska et al. showed that costimulation of HUVECs with IL-1 and TGFβ2 induces EndMT, further indicating that inflammatory signaling can induce EndMT. 65 Interestingly, IFN-α and IFN-γ can either downregulate or respectively upregulate EndMT-related genes in human dermal microvascular endothelial cells, indicating that some modulation of EndMT is possible depending on which IFN types are present. 66 Based on the studies described above, the inflammatory environment provoked by the implantation of tissue-engineered vascular grafts is likely to induce EndMT (Fig. 2). Therefore, incorporating a slow release of immunomodulating factors, such as ILs or growth factors directly in the scaffold material, is a potential approach to guiding inflammatory signaling and mediating EndMT.
The Mechanical In-Situ Environment of a Vascular Graft Can Modulate EndMT
Hemodynamic shear patterns can inhibit or stimulate EndMT
Endothelium experiences shear stress as blood flows by. Shear stress (τ) relates to the viscosity of the blood (μ), the fluid flow (Q), and the radius of the blood vessel (r) in the following equation33,67: τ = 4 μQ/πr 3 . The shear stress found along the endothelium of the human aorta ranges between 10 and 20 dynes/cm2. 68 Along the surface of the aortic heart valve, however, shear stress ranges between 30 and 1500 dynes/cm2. 69 The patterns of blood flow vary enormously depending on the location within the circulatory system, and endothelial cells are capable of responding to different patterns of flow in a number of ways. Endothelial cells are capable of sensing fluid flow through adhesion molecules, 70 cytoskeletal deformation, 71 nuclear displacement, 72 and cilia; small organelles on the cell membranes capable of translating mechanical stimulation to intracellular signaling.68,73–75 The ciliary state of the endothelium responds to the differing fluid patterns, where a laminar flow with high shear stress results in loss of cilia, and a disturbed flow with low shear stress, such as found in bifurcations and around valves, correlates to ciliated endothelium. 76 The endothelial response to shear stress has been associated with the development of atherosclerosis, and a growing body of evidence suggests that EndMT plays a role in this pathological development. Adult endothelium responds with programs of signaling and development that differ from embryonic endothelium. Egorova et al. have shown that embryonic endothelium is capable of responding to shear patterns with EndMT through TGFβ/ALK5 signaling, suggesting that fluid flow may have a major influence on the developing implanted vascular grafts (Fig. 2). The response of ECFCs to shear stress appears to be similar to that of adult endothelium 77 ; however, it is not fully known what the effect of other mechanical forces, such as strain, will be on EndMT, ECM production, and remodeling in these candidate cells for tissue engineering. In addition, manipulating the ciliary state of ECFCs may influence their tendency to undergo EndMT, where a lack of cilia primes endothelial cells for shear-induced EndMT through TGFβ/ALK5 signaling, 78 but also for calcification through BMP signaling. 79 This is of particular relevance to heart valve replacements, where the entire graft needs to develop homogenously while the ventricular face of the valve leaflet will experience a pulsatile and laminar flow pattern every systole, and the aortic face of the valve leaflets will experience a turbulent pattern of fluid flow. 80 Due to the differences in flow patterns, the ventricular face, therefore, may lack cilia and be more prone to undergoing EndMT compared to the aortic face. Incorporating chemical factors on the aortic face of the graft to chemically induce the loss of cilia 81 may be used to correct this discrepancy.
Mechanical strain influences mechanical integrity through ECM production and alignment
Statically cultured ECFCs have been reported to arrange ECM fibers along the fiber axis of electrospun materials, whereas mature endothelial cells (HUVECs) arrange their fibers perpendicularly, suggesting that ECFCs do not fully behave like mature endothelium. 82 Understanding these effects is particularly important in the context of heart valves and heart valve replacement grafts, since in addition to fluid flow, vascular structures experience considerable strain. For blood vessels, this strain is perpendicular to the blood flow and dependent on the blood pressure and the mechanical properties of the vessel wall (Fig. 2). For heart valves, the strain is distributed along a specific pattern in each valve leaflet, 83 and the ECM of the valve is adapted to the pattern allowing for ideal mechanical support. 84 Underlining how different levels of shear can induce different mechanisms, sheep valve endothelial cells exposed to high and low cyclic mechanical strain have been shown to undergo EndMT through Wnt/β-catenin or TGFβ signaling, respectively. 85 Mechanical strain also has a strong influence on the ECM remodeling behavior of myofibroblasts. 86 Strain, therefore, could both induce endothelial cells to transition to myofibroblast-like cells, and guide their ECM production and remodeling. While it has been previously shown that strain may inhibit EndMT in ECFCs, pretreating the cells with TGFβ1 enables ECFCs to respond to a mechanical environment with increased matrix production.11,84 Synthetic scaffolds may be designed to distribute mechanical stresses along thicker fibers and in such a way that the tissue development is maximally stimulated, for instance, by both guiding the alignment of ECM produced by cells and relieving areas of the graft of maximal strain to avoid inhibition of EndMT.
Bioreactors Combine Biochemical and Mechanical Stimuli to Investigate EndMT
To achieve control over EndMT for vascular tissue engineering it is crucial to consider the combined effects of mechanical stresses, biochemical factors, and the immune response. Importantly, in addition to inducing EndMT, it is also likely that some inhibition of EndMT is required in regions of implanted vascular grafts prone to fibrosis or calcification. To achieve a well-timed induction or inhibition of EndMT, in-depth knowledge is required of the combinations of stimuli found in situ. In vitro tools such as bioreactors can allow simultaneous study of mechanical and biochemical stimuli. Mahler et al. developed a bioreactor with the capacity to test 3-dimensional cultures, including cocultures of multiple cell types. 69 Using a single cell type they have reported the development of a migratory phenotype out of an endothelial monolayer up to a depth of 60 μm deep in response to fluid shear, along with an increased expression of α-SMA and Snail, indicating EndMT. Matrix production or remodeling was not measured. Bioreactors such as these may allow the testing of bioactive materials as well as hemodynamic variables. However, in this analysis the endothelial cells were exposed to a constant single shear pattern for 48 h, whereas in vivo the hemodynamic patterns change along with pulsations. Pulsatile flow patterns can be included in bioreactor designs such as described by Smits et al. 87 Egorova et al. used a bioreactor to study the response of ECFCs, HUVECs, and HUAECs to shear stress. All cell types were subjected to low and high shear stress for 5 h using a parallel plate flow chamber, and a similar transcriptional response was found between the cell types pointing to a response of ECFCs to shear similar to adult endothelium. 77 The adhesion of cells to peptide-linked polymers in shear flow can be studied using a radial flow chamber, where the shear rate varies linearly with radial position, allowing for the study of the effect of a range of shear rates. 88
Exposing cell-seeded grafts to appropriate shear and strain stresses may help to develop a mechanically capable vascular graft with not only the proper cell populations, but also with an appropriate ECM fiber orientation level. 84 Emphasizing the importance of the orientation of mechanical force, Balachandran et al. used a bioreactor to apply strain regimes on a VEC monolayer to induce EndMT, and found that when cyclic strain was applied orthogonal, instead of parallel, to the tissue alignment, EndMT increased and the cell microarchitecture was disrupted. 85 The bioreactors described above were mostly used to study mechanical stimulation. Ideally, relevant mechanical and biochemical stimuli would be included in a single bioreactor, allowing for a combined approach. Liang et al. reported on synergistic effects of mechanical strain and TGFβ stimulation on vascular wall function in a bioreactor applying cyclic strain. 89 Such bioreactors would allow for the investigation of EndMT in an environment approaching the intended environment of cardiovascular graft. In addition, bioreactors capable of combining mechanical and biochemical stimuli may be used to guide the development of cellularized grafts in vitro, before implantation in vivo.
Conclusions
A tissue-engineered vascular structure can be designed to exploit the in situ stimuli and guide tissue formation by regulating EndMT. Advanced molecular design of biomaterials and production methods will dictate the possibilities of including these factors. Novel biomaterials can incorporate the timed release of multiple EndMT inducing or limiting factors in controlled concentrations. Provided that a graft contains endothelial cells, these factors may boost cell migration and ECM production to achieve a mechanically sufficient interstitium. Considering that the inflammatory and mechanical environment may combine to push EndMT toward a fibrotic development, inhibitory factors should also be considered. ROS inhibitors such as astaxanthin, for instance, could be administered to halt EndMT after tissue formation is sufficient. In addition, the fine-tuning of the ECM can be aided by regulation by mirs, potentially delivered through microvesicles. The content of these vesicles can be controlled, for instance, by the transfection of expression vectors into the cells producing the extracellular microvesicles. 56 Furthermore, since ECM deposition and remodeling is highly dependent on both the alignment of cells adhering to materials and hemodynamic flow patterns, the structural design of cardiovascular grafts in itself provides options to manipulate EndMT. Additional research, for instance, using bioreactors capable of applying simultaneous mechanical and biochemical stimulation, is needed to accurately predict how candidate cell sources will respond to the complex environment of an implanted vascular replacement graft. This approach may lead to an improved clinical transition of functionalized implanted scaffolds, carefully designed to develop into living, functional cardiovascular replacements.
Footnotes
Acknowledgments
This research forms part of the Project P1.01 iValve of the research program of the BioMedical Materials Institute, cofunded by the Dutch Ministry of Economic Affairs. The financial contribution of the Nederlandse Hartstichting is gratefully acknowledged. The authors acknowledge the support for PHAEDRA (CVON-2012-08) and for 1Valve (CVON-2012-01) from the Netherlands CardioVascular Research Initiative: the Dutch Heart Foundation, Dutch Federation of University Medical Centers, the Netherlands Organization for Health Research and Development, and the Royal Netherlands Academy of Sciences.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.