
Abstract
Gene transfer has been used experimentally to promote chondrogenesis and cartilage regeneration. While it is controversial to apply gene therapy for nonlethal conditions such as cartilage defect, there is a possibility that the transfer of therapeutic transgenes may dramatically increase the effectiveness of cell therapy and reduce the quantity of cells that are needed to regenerate cartilage. Single or combination of growth factors and transcription factors has been transferred to mesenchymal stem cells or articular chondrocytes using both nonviral and viral approaches. The current challenge for the clinical applications of genetically modified cells is ensuring the safety of gene therapy while guaranteeing effectiveness. Viral gene delivery methods have been mainstays currently with enhanced safety features being recently refined. On the other hand, efficiency has been greatly improved in nonviral delivery. This review summarizes the history and recent update on the gene transfer to enhance chondrogenesis from stem cells or articular chondrocytes.
Introduction
A
To solve the dilemma and limitations of autologous chondrocyte implantation, several strategies were developed. The first strategy is the use of matrix materials, such as hyaluronic acid and collagen, to maintain the chondrocyte phenotypes.5–8 Another concept is the use of other cell sources including stem cells. 9 Numerous experimental studies have been performed with the goal of realizing the latter concept because stem cells have the advantages of multilineage differentiation and proliferation capability. Inadequate differentiation and hypertrophy are main obstacles in the successful chondrogenesis from adult stem cells. 10 Various growth factors and biomaterials have been employed to resolve these problems. In addition to these measures, genetic modification has also been explored to drastically alter the characteristics of stem cells into chondrogenic phenotypes.11–13 Gene transfer was also performed in dedifferentiated chondrocyte to restore the chondrogenic phenotypes lost during expansion culture.14,15
While the use of gene therapy for nonlethal conditions such as cartilage defect is debatable, there is a possibility that the transfer of therapeutic transgene may dramatically increase the effectiveness of cell therapy and reduce the quantity of cells needed to regenerate cartilage. This review summarizes the history and recent updates on the gene transfer to enhance chondrogenesis from stem cells or articular chondrocytes.
Overview of Gene Transfer
Gene transfer involves introduction of genetic material into cells to alter cellular function or structure at the molecular level. When gene transfer is performed to treat a disease, it is called gene therapy. 16 Gene transfer can be done in vivo or ex vivo. In in vivo gene transfer, a vector containing the therapeutic gene is directly injected into the recipient. During ex vivo gene transfer, cells receive transgenes in vitro before introduction in the recipient. While the ex vivo approach is more time-consuming and expensive, it enables gene delivery to a specific cell population and screening of the cells before implantation. 17 Therefore, most of the studies introduced in the review were performed using ex vivo gene transfer.
The goal of gene therapy is the resolution of clinical problem by the translation of therapeutic protein from the transferred gene. As the mammalian cell membrane acts as a barrier to DNA, a vector that can carry the genetic material into the target cells is necessary and essential for gene transfer. Only after the therapeutic gene gets inside the cells is the transcription of genetic information to mRNA and translation from mRNA into the protein possible. 18 Vectors are generally categorized as nonviral or viral. Nonviral gene transfer is termed transfection and viral gene transfer is termed transduction. 19
The main advantage of viral gene transfer is high efficiency in transferring of the genetic payload to the target cells they infect. To develop vectors for gene transfer modifications are made to remove pathogenic components from the viral genome, which increases safety and creates genetic space for transgene introduction. 20 Certain viral vectors only infect dividing cells, while other viral vectors infect both dividing and nondividing cells. Some vectors incorporate into the genome of host cells, while other vectors remain episomal. 21 Not surprisingly, safety issues are concerns in viral gene therapy, even though viral genomic structures have been modified to reduce pathogenicity. 19 Adenoviral vectors induce an immune response that can limit their clinical efficacy. 18 Retroviral vectors can cause oncogenesis following integration into the host genome. These risks arising from the use of viruses are not justified for nonlethal musculoskeletal diseases.
In nonviral gene transfer, physical or chemical methods are used to facilitate cellular uptake of DNA. The former comprises ultrasound, electric pulse (electroporation), and gene gun technology. The latter utilizes a carrier, such as a liposome or other polymer, which associates or complexes with DNA. Nonviral vectors generally fall behind viral vectors in efficiency, magnitude, and duration of transgene expression. Nevertheless, recent advances in the nonviral gene transfer technology have markedly enhanced the transfer efficiency of nonviral vectors.
The type of gene transfer depends on the specific clinical situations, either systemic or local. While genetic disease such as enzyme deficiencies may require lifelong expression of a transgene, posttraumatic conditions like segmental bone defects require only the transient expression of the transferred genes. 22 On the other hand, it is still undetermined whether transient or permanent transgene expression is necessary to treat chronic degenerative diseases like osteoarthritis. The local increase in anabolic factors or presence of a catabolic factor antagonist can promote regeneration or arrested destruction of cartilaginous tissue. However, excessive and prolonged expression of anabolic factors can be detrimental, as seen in the formation of osteophytes with adenovirus-mediated transfer of transforming growth factor (TGF)-β1. 23 Expression of therapeutic factors for a limited period of time may provide a reasonable choice. In gene transfer for cartilage regeneration, long-term in vivo observation that includes monitoring of transgene expression in large animal models will be necessary to determine the most appropriate period of transgene expression for each gene transfer method.
Cell Sources, Carrier Material, Vectors, and Reporter Gene Expression in Ex Vivo Gene Transfer to Promote Chondrogenesis
Articular chondrocytes and mesenchymal stem cells (MSCs) have been investigated as options for implantation therapy to treat human articular cartilage defects. While articular chondrocytes and MSCs are usually transplanted,24–30 fibroblasts,31,32 perichondrial cells, 33 periosteal cells,34,35 and muscle-derived cells (MDCs) 36 have been also applied. Fibroblasts expressing active TGF-β1 were injected into the knee joints of rabbits with artificially made cartilage defects. Four to 6 weeks after the injection, the cartilage defect was completely filled with newly grown hyaline cartilage. 31 NIH3T3 cells were transfected with expression plasmid vectors containing the beta-galactosidase (LacZ), human insulin-like growth factor (IGF)-1 gene, or both human IGF-1 and fibroblast growth factor (FGF)-2 genes, and transplanted into osteochondral defects in rabbits. After 3 weeks, transplantation of the co-transfected IGF-I/FGF-2 implants improved articular cartilage repair, which were better than with IGF-I alone or when compared to LacZ implants. 32 Plasmids carrying LacZ, parathyroid hormone-related peptide (PTHrP), and TGF-β1 genes were transfected into rabbit perichondrial cells. When polylactic acid scaffolds seeded with transfected perichondrial cells were implanted into osteochondral defects in rabbit knees, the transformed cells continued to overexpress transgenes in vivo for the entire test period of 7 days. 33 Rabbit periosteum-derived cells were infected with a recombinant adeno-associated virus (AAV) expressing LacZ gene (AAV-LacZ). Collagen gel containing the LacZ-transferred periosteum-derived cells was transplanted into a full-thickness articular cartilage defects in rabbits. The LacZ-positive cells were recognized at the transplantation sites in 8 out of 10 knees. 34 Rabbit MDCs and chondrocytes were transduced with a retrovirus encoding for the LacZ. The cells were embedded in type I collagen gels, and grafted to the full-thickness osteochondral defects. The repaired tissues in the MDC and chondrocyte groups had similarly better histology than those of control groups. Repaired tissues in the MDC group were mainly composed of type II collagen, as in the chondrocyte group. 36
Implantation of ex vivo gene-transferred cells to heal cartilaginous defect in vivo has been performed using cell aggregates 37 or cells encapsulated in a carrier material like alginate,38–40 agarose,41,42 and fibrin or type-I collagen gels,26,43–45 seeded in synthetic biodegradable scaffolds,46,47 or as tissue-engineered cartilage. 48
Both nonviral and viral approaches can be used to transfer therapeutic genes to articular chondrocytes and MSCs, which can be then transplanted into articular cartilage defects in vivo to produce clinically relevant sustained levels of therapeutic protein. The first report used a retroviral vector to transduce chondrocytes and implant cells into an articular cartilage defect in vivo. 27 Subsequent studies used nonviral,28,30,33 adenoviral,26,37,49 retroviral,27,29,35,36,50 and recombinant AAV (rAAV) vectors to deliver reporter genes to defects via ex vivo approaches.28,34,51–58 While most of the evaluations were carried out in small animals including rats and rabbits, large animal models such as caprine or equine models provide data that would more closely predict human results.
Tracking of implanted cells is necessary to monitor the transduction efficiency and to detect the localization of transgenes after gene transfer to target cells. Green fluorescence protein (GFP) reporter gene has been most popularly used to track gene transfer in both viral and nonviral systems. 11 The efficiency of gene delivery varies with cells. An AAV vector to deliver GFP to primary human chondrocytes achieved a high efficiency of 93.7% at 7 days after transduction, gene expression being maintained for up to 28 days after transduction.26,59 On the other hand, AAV vector-mediated GFP delivered to MSCs showed a maximum transduction efficiency of 65%. 60 Our group had transfected adipose stem cells (ASCs) with GFP using electroporation, reaching a tranfection efficiency of 83.7%. 61
Gene Transfer of Growth Factors to Promote Chondrogenesis and Cartilage Regeneration
The rationale for gene transfer of growth factors in cell therapy is to make implanted cells produce active recombinant peptide that will generate a local environment more favorable for, and thus further enhance articular cartilage repair. 11 Cell type and the efficiency of gene transfer critically affect the amount of a secreted therapeutic protein. The duration of transgene expression depends on individual method for gene transfer. The choice of therapeutic genes is based on the strategy selected for improving articular cartilage repair. Genes that encode products that stimulate chondrogenesis or inhibit the degeneration of articular cartilage may be used depending on the therapeutic intent. Table 1 summarizes studies that have used gene transfer of growth factors to enhance chondrogenesis.
AAV, adeno-associated virus; ASC, adipose stem cells; BMP, bone morphogenetic protein; ECM, extracellular matrix; FGF, fibroblast growth factor; hMSCs, human mesenchymal stem cells; IGF, insulin-like growth factor; IHH, Indian hedgehog; MDSCs, muscle-derived stem cells; MSCs, mesenchymal stem cells; PCL, poly(ɛ-caprolactone); rAAV, recombinant AAV; TGF, transforming growth factor.
The TGF-β superfamily of growth factors induces chondrogenic differentiation of MSCs and maintains the chondrogenic phenotype. 62 They have been considered as candidates for gene therapy to enhance chondrogenesis. TGF-β1 is included in classical chondrogenic differentiation medium 62 although TGF-β3 is being increasingly used. 4 In one study, adult human MSCs (hMSCs) were transduced with AAV-GFP or AAV-TGF-β1 and implanted into osteochondral defects of 21 athymic rats; improved cartilage repair was observed at 12 weeks. 57
hMSCs were transduced with adenoviral vectors carrying genes for TGF-β1 (AdTGF-β1) and/or IGF-1 (AdIGF-1). The transduction of AdTGF-β1, but not AdIGF-1, demonstrated robust chondrogenic differentiation. 63 Adenoviral delivery of TGF-β1 reportedly enhanced chondrogenesis of hMSCs, with the secretion of ∼5 ng/mL of the peptide into the culture medium, while unmodified cells required 10–24 ng of exogenous TGF-β1 to achieve the same degree of chondrogenesis. Namely, constant expression of TGF-β1 through gene delivery was more effective than intermittent addition of growth factor peptide with a short half-life. 63
When autologous bone marrow clots containing AdTGF-β1-transduced cells were implanted into partial thickness chondral defects in sheep, improved histological, biochemical, and biomechanical parameters were evident 6 months postoperatively. 64 Nonviral delivery of TGF-β1 using plasmid DNA (pDNA) immobilized on a chitosan-based scaffold also successfully resulted in chondrogenic induction of MSCs. 65 When plasmid-borne TGF-β1-transfected MSCs were seeded into polylactide scaffolds and implanted on full-thickness articular cartilage defects in rabbits, improved extracellular matrix (ECM) formation and reconstitution of subchondral bone, and inhibited inflammatory responses were observed after 24 weeks. 66 TGF-β3 was also delivered to human bone marrow MSCs using lentiviral vectors immobilized on poly(ɛ-caprolactone) woven scaffolds. Transduced MSCs undergoing chondrogenic differentiation produced similar levels of ECM production when compared to untransduced MSCs that received recombinant TGF-β3 protein. 67 TGF-β3 has also been transduced to ASCs using a baculoviral vector. Collagen II and matrix protein expression were enhanced in TGF-β3-transduced cells and improved biomechanical properties. 68
Bone morphogenetic proteins (BMPs) are other members of the TGF-β superfamily. BMP gene transfer has been used to improve chondrogenesis. A strong interaction between TGF-β and BMPs during developmental chondrogenesis has been described. 69 BMPs also induce chondrogenesis from stem cells. As chondrogenic induction from BMPs is considered an intermediate step to bone formation, BMPs should be applied, tuned in accordance to the multitude of other stimuli present. BMP-2 is the most commonly investigated BMP.70,71 Retroviral BMP-2 transduction is highly effective at inducing a chondrocyte phenotype in multipotential mesenchymal C3H10T1/2 cells in vitro. 72 Adenoviral transduction of MSCs from various tissues with BMP-2 can increase cell proliferation and collagen II matrix protein secretion. Perichondrium-derived MSCs show the highest chondrogenic capacity, followed by bone marrow MSCs and adipose-derived MSCs. 73 pDNA delivery of BMP-2 enhances chondrogenesis by increasing Sox-9 expression. 74 It has been proposed that the effect of BMP-2 is dependent on the Sox-9 status of the cell, with high Sox-9 leading to BMP-2 driven chondrogenesis, while low Sox-9 leads to BMP-2-driven osteogenesis. 72 This finding bears on the application of BMP-2 for chondrogenic enhancement in a clinical setting, as constitutive expression of BMP-2 leads to chondrocyte hypertrophy followed by osteogenic differentiation. 72 Adenoviral transduction of human bone marrow MSCs, harboring the genes for BMP-2 or BMP-4, has demonstrated the equal effectiveness of the BMP types in provoking chondrogenesis in a pellet culture. However, this triggered chondrogenesis is prone to hypertrophy. 75 Application of retroviral BMP-4-transduced muscle-derived stem cells on full-thickness chondral defects of nude rats using fibrin glue has demonstrated significant improvement in histologic repair compared with control defects. 76 Adenoviral transduction of BMP-7 in equine chondrocytes can induce earlier articular cartilage matrix generation than untransduced control when transplanted in vivo. Articular cartilage defects implanted with transduced chondrocytes reportedly allowed a better appearance of the repair tissue after 4 weeks, although no differences were found between two groups after 8 months. 25 In another study, C3H10T1/2 mesenchymal progenitor cells were transduced by an adenoviral construct containing hBMP-13 or hBMP-2. BMP-13 supported chondrogenesis, but not terminal differentiation, whereas BMP-2 stimulated endochondral ossification. hBMP-13, while stimulating chondrogenesis, failed to support differentiation to hypertrophic chondrocytes and endochondral ossification similar to hBMP-2. 77
IGF-1 enhances proliferation and differentiation of articular chondrocytes. 78 IGF-1 simultaneously stimulates type-II collagen and proteoglycan synthesis, and cell proliferation. 74 In one study, bovine articular chondrocytes were transfected with a plasmid vector containing the cDNA for human IGF-1 using lipofection. Transplantation of IGF-1-transfected articular chondrocytes onto the surface of articular cartilage led to thicker tissue containing more cells and glycosaminoglycans (GAGs) than the control. 74 These results suggest that the transfection of IGF-1 might simultaneously promote chondrogenesis and shift cartilage homeostasis in an anabolic direction. When equine chondrocytes transduced by an adenovirus vector encoding IGF-1 were arthroscopically implanted on chondral defect, increased IGF-1 mRNA and ligand production was observed in repair tissue in an equine model for up to 9 weeks following transplantation. Collagen type II expression was significantly increased in IGF-1-treated defects. Adenovirus vector encoding IGF-1 improved early (4–9 weeks) and, to a lesser degree, long-term cartilage repair in chondrocytes over control defects in vivo. 79 Nonviral delivery of IGF-1 in alginate gels into rabbit articular chondrocytes also improved articular cartilage repair and accelerated the formation of the subchondral bone in vivo compared with the control. 40 IGF-1 pDNA immobilized on collagen II-GAG scaffolds has been delivered to canine articular chondrocytes. The sustained IGF-1 overexpression resulted in significantly higher amounts of chondrocyte-like cells, GAG accumulation, and type II collagen production compared with control scaffolds. 80
FGF-2 is a mitogenic factor that may be useful for increasing the pool of cells in a cartilage defect responsive to chondrogenesis. Liposome-mediated transfection of hFGF-2 stimulated cell proliferation in bovine articular chondrocytes without effect upon matrix GAG synthesis. 81 AAV-hFGF-2-transduced monolayer and aggregate cultures of hMSCs led to enhanced, dose-dependent cell proliferation compared with the control. While chondrogenic differentiation (proteoglycans, type II collagen, and Sox-9 expression) was successfully achieved without significant difference between conditions, application of AAV-hFGF-2 reduced the expression of type I and type X collagen. 82 FGF-2 has also been delivered to rabbit chondrocytes by transfection of pDNA using liposome. FGF-2 gene transfer enhanced type II collagen expression and individual parameters of chondrogenesis including the cell morphology and architecture of the new tissue. 39
Indian hedgehog (IHH) protein is also known to induce chondrogenesis from hMSCs. 83 When autologous bone marrow coagulates with adenoviral vector particles encoding BMP-2, IHH, or GFP control were delivered to osteochondral defects in the trochlea of rabbit knees, IHH treatment resulted in superior histological repair cartilage quality than controls in all of the assessed parameters while BMP-2 treatment led to substantially variable results, including severe intralesional bone formation in two of six joints after 13 weeks. 84
Delivery of Multiple Factors
More than two factors that have different mechanism of action can be used to enhance chondrogenesis. An example is the co-application of FGF-2 and IGF-1, which targets cell proliferation and anabolic function, respectively. Table 2 summarizes studies that used gene transfer of multiple factors to enhance chondrogenesis.
Cbfa1, core binding factor alpha1; shRNA, short hairpin RNA; siRNA, small interfering RNA.
Adenoviral transduction of IGF-1 and FGF-2 in ovine ASCs showed synergistic effects of IGF-1 and FGF-2 in chondrogenic differentiation, showing higher significant expression levels of aggrecan, biglycan, cartilage matrix, proteoglycan, type II collagen, and very low expression of type I collagen or hypertrophic marker type X collagen. 85 In another study, NIH 3T3 cells were transfected with expression plasmid vectors harboring cDNA containing human IGF-1 gene and/or human FGF-2 gene and implanted into osteochondral defects in the trochlear groove of rabbits. Transplantation of the co-transfected IGF-1/FGF-2 implants increased the DNA content of the repair tissue, accelerated the formation of the subchondral bone, and improved articular cartilage repair more than observed with IGF-I alone. 32 Lapine articular chondrocytes were also transfected with expression plasmid vectors of IGF-1 and FGF-2, encapsulated in alginate, and transplanted into osteochondral defects in the knee joints of rabbits in vivo. Co-overexpression of IGF-1 and FGF-2 by the transplanted articular chondrocytes enhanced the early repair of cartilage defects in vivo and protected the neighboring cartilage from degeneration. 86 Adenoviral co-transduction of human bone marrow MSCs in vitro with IGF-1 and TGF-β1 or BMP-2 had synergistic effects in chondrogenic enhancement with increased collagen II expression. However, a high level of collagen X expression in transduced cells was observed. 87
Growth factor can be combined with transcription factors or RNA inhibitors. When Sox-9 and BMP-2 were nonvirally delivered in a bicistronic plasmid vector into dedifferentiated human chondrocytes expressing a weak chondrogenic phenotype, significantly increased expression of type II collagen and aggrecan was observed compared with individual delivery of the BMP-2 or Sox-9 gene. Six weeks after in vivo transplantation, a synergistic effect with rapid redifferentiation of the chondrocytes was also demonstrated. 14 Bicistronic lentiviral delivery of the growth factor TGF-β3 and a short hairpin RNA targeting type I collagen gene enhanced chondrogenic differentiation of synovium-derived MSCs. 88 Nonviral delivery of Sox-9 pDNA enhance chondrogenesis of human MSCs when co-delivered with small interfering RNA to core binding factor alpha1 (Cbfa1), the osteogenic transcription factor. 89
Retroviral transduction has been employed to deliver transcription factors Klf4 and c-Myc with Sox-9 to reprogram somatic cells into chondroprogenitor cells. Combining transcription factors (Oct-4, Klf-4, C-Myc, and Sox-2) that can reprogram somatic cells into a pluripotent stem cell phenotype along with chondrogenic transcription factor Sox-9, mouse dermal fibroblasts were directly reprogrammed into chondrocytes, bypassing the induced pluripotent stem cell state. 90 Of several combinations, the overexpression of Klf4 and c-Myc with Sox-9 has yielded the highest efficiency in chondrogenic differentiation, resulting in secretion of ECM with physical and histological properties as those of articular cartilage. The combination also worked for human dermal fibroblasts to chondrocytes. 91
Overexpression of Transcription Factors to Enhance Chondrogenesis
The chief advantage in transduction of chondrogenic transcription factors instead of growth factors is that the transcription factors act inside cells and are not secreted from cells unlike growth factors. Functional control of chondrogenic activity is possible, depending on the number of injected cells. Transfer of chondrogenic transcription factors can enhance subsequent expression of target chondrogenic genes, such as collagen II and aggrecan,92,93 which are often lost because of cellular dedifferentiation.
Sox-9 is a key chondrogenic factor that directly binds to the promoter region of collagen II and aggrecan, and transcriptionally activates the expression of these genes in both differentiating MSCs and chondrocytes. 92 Viral transduction of Sox-9 has been achieved using adenovirus, 94 retrovirus, 95 AAV,52,96,97 and lentivirus. 92 When these viruses were compared in their ability to deliver Sox-9 to human osteoarthritic chondrocytes and induce chondrogenic phenotype and ECM, lentiviral transduction was confirmed to be the most appropriate vector system for Sox-9 delivery with a transduction efficiency of 85%. 92 The delivery of Sox-9 was also efficiently performed in explant cultures of human cartilage using AAV vectors, showing enhanced expression of ECM components. 52 Adenoviral transduction of Sox-9 delivery into rabbit MSCs has demonstrated the maintenance of ectopic expression when transduced cells were implanted into osteochondral defects in rabbits, resulting in successful integration of the graft tissue. 94
In vitro AAV-mediated Sox-9 transduction in human bone marrow MSCs has also resulted in a significant reduction in hypertrophy markers. 96 Marked and prolonged expression of the transcription factor was noted in undifferentiated and chondrogenically differentiated cells transduced with rAAV-hSox-9, leading to increased synthesis of major ECM components compared with control treatments, but without an effect on proliferative activities. rAAV- hSox-9 delivery reduced the levels of markers of hypertrophy, terminal and osteogenic/adipogenic differentiation in hMSCs with diminished expression of the osteoblast-related transcription factors. 96 The underlying mechanisms appear to be a decrease in β-catenin expression, a mediator of the Wnt signaling pathway known to stimulate osteoblast lineage differentiation, and an increase in PTHrP, an inhibitor of hypertrophy that reduces the gene expression for COL1A1, COL10A1, alkaline phosphatase, Cbfa1, and activates Sox-9 transcriptional activities. 96 Retroviral transduction of Sox-9 enhanced the re-expression of the chondrocyte phenotype in passaged osteoarthritic human articular chondrocytes. 95 Nonviral delivery of Sox-9 was attempted by lipofection in mouse MSCs. Efficient ectopic expression of Sox-9 was demonstrated in vitro and GAG production was evident 4 weeks after in vivo implantation in athymic mice. 93 A tetracycline inducible Sox-9 transfection system using a biodegradable scaffold for chondrogenic stimulation has also been used to enhance the phenotype of rat chondrocytes. 98
Sox-5 and Sox-6 are cofactors of Sox-9. Together these factors constitute the Sox trio. Adenoviral transduction of the Sox trio into fibroblasts is able to transform these cells into chondrocyte-like cells without addition of any growth factors. Our previous data show that retroviral gene transfer of Sox trio to rat ASCs can also enhance the in vitro chondrogenic differentiation of ASCs. ASCs co-transduced with the Sox trio in a fibrin gel act to promote healing of osteochondral defects and arrest the progression of surgically induced osteoarthritis in a rat model. 99 Our group used electroporation to transfect hMSCs and hASCs with the Sox trio. Gene transfer efficiency was high (∼70%). Transfected cells showed high expression of corresponding genes after 21 days, and each Sox protein was detected in ASCs transfected with the corresponding gene. The chondrogenic differentiation of ASCs, as demonstrated by GAG levels (up to sevenfold) and Safranin-O staining, showed significant enhancement following Sox trio co-transfection. The co-transfection enhanced COL2A1 mRNA, while COL1A1 and COL10A1 mRNA remained level or decreased. Type II collagen protein markedly increased and type X collagen decreased with co-transfection of the Sox trio. When pellets were implanted in the subcutaneous pouch of nude mice for 3 weeks, ASCs co-transfected with Sox trio demonstrated abundant proteoglycan and significantly reduced mineralization (Fig. 1). 61 As a further step, we developed a chondrogenic scaffold system in which pDNA containing the Sox trio genes was incorporated into a poly(lactic-co-glycolic acid) (PLGA) scaffold and slowly released to transfect ASCs seeded in the scaffold. The pDNA/PEI-PEG complex-incorporated PLGA/pluronic F127 porous scaffolds were fabricated by a precipitation/particulate leaching method. The transfection rate was around 70%. After 21 days, ASC/PLGA scaffold constructs incorporating each gene individually showed markedly increased expression of the corresponding gene and protein. The GAG and COL2A1 genes and proteins were notably increased in Sox trio pDNA-incorporated scaffolds than in the control, while COL10A1 protein expression decreased. An in vivo implantation study in a rabbit model demonstrated enhanced cartilage regeneration with ASCs/Sox trio pDNA-incorporated PLGA scaffolds implanted in osteochondral defects after 8 weeks (Fig. 2). 100

ASCs transfected with the gene for green fluorescence protein (GFP) (pEGF-C1), SOX-5, SOX-6, or SOX-9, and all three genes (SOX trio) using electroporation in pellet culture 21 days after transfection and pellet culture. Staining with 4′,6-diamidino-2-phenylindole (DAPI) staining (blue, indicating nuclei) was merged with GFP fluorescent imaging (green)
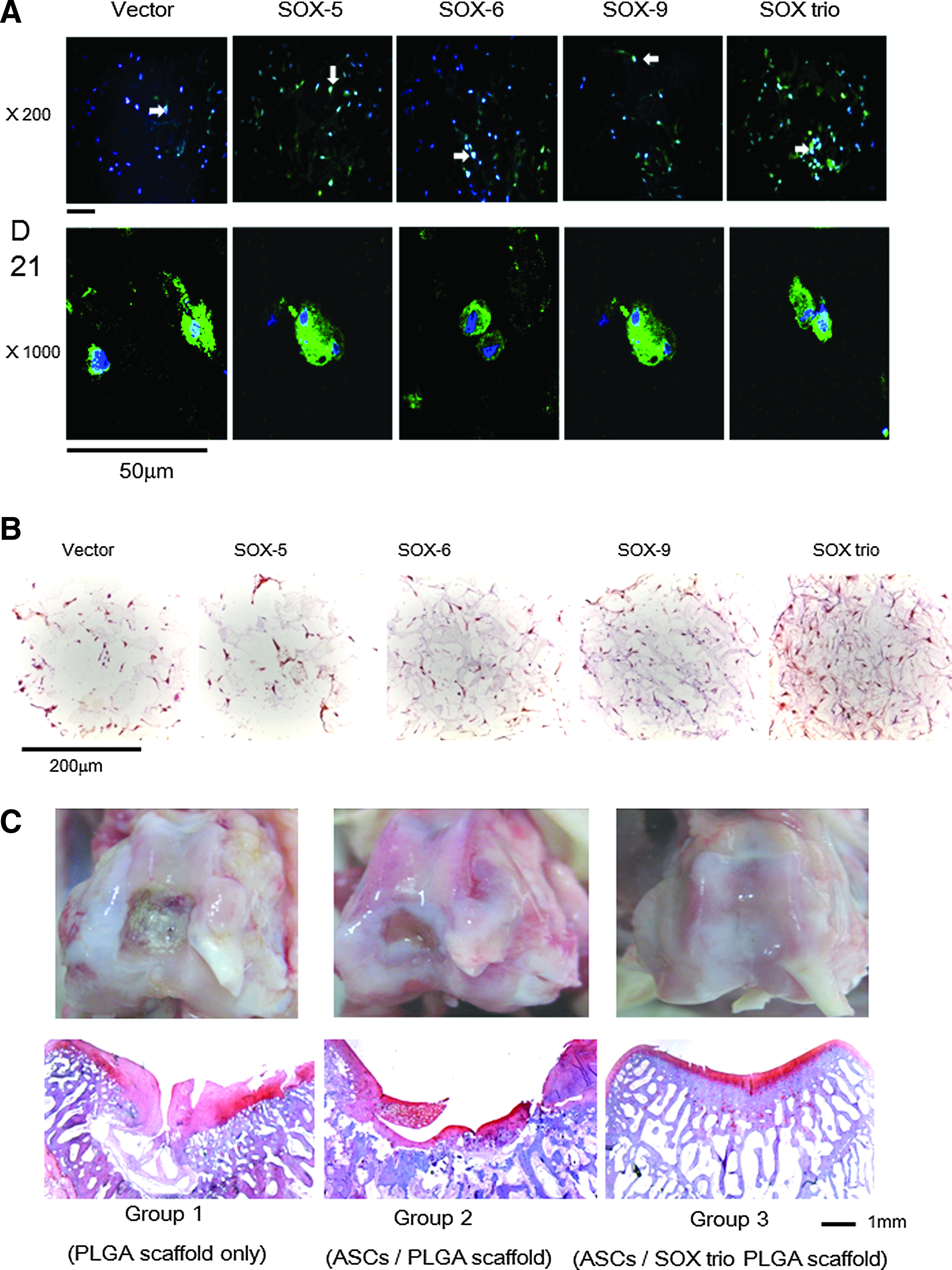
Fluorescence microscopy appearance of ASC/plasmid DNA (pDNA)-incorporated scaffold after 21 days of in vitro culture. DAPI staining (blue, indicating nuclei) was merged with GFP fluorescent imaging (green). Transfected cells look bright greenish-blue on the magnification ×200 (white arrows), with more detailed appearance provided at a magnification of ×1000. Scale bar represents 50 μm
Besides Sox factors, other transcription factors have been studied in vitro. Smad3 is a key downstream signaling molecule in the TGF-β-mediated chondrogenic differentiation pathway, which functions upstream to Sox-9. Adenoviral transduction of human MSCs with Smad3 strongly enhanced the transcriptional activity of Sox-9 and COL2A1, and upregulated ECM protein secretion. 101 The homeobox transcription factor Barx2 is a regulator of chondrogenesis during embryonic development. Retroviral transduction of mouse embryonic MSCs with Barx2 increased cell aggregation before chondrogenic differentiation. Co-transduction of Barx2 with Sox-9 increased in type collagen II production through Barx2 interaction with Sox-9 (Table 3). 102
GFP, green fluorescence protein; pDNA, plasmid DNA; PLGA, poly(lactic-co-glycolic acid).
Summary and Future Perspective
Various transgene and gene transfer methods have been used to enhance chondrogenic differentiation and hypertrophy in adult stem cells and to reduce dedifferentiation in culture expanded articular chondrocytes. The current challenge for the clinical applications of gene modified cells is ensuring the safety of gene therapy while guaranteeing effective therapeutic levels of the translated factors. Viral gene delivery methods have been mainstays in the literature. Enhanced safety features recently have shown great potential for clinical application of such technologies. On the other hand, efficiency has been greatly improved in nonviral delivery, which also includes the use of various scaffold materials. Considering that cartilage defect is nonlethal, unlike genetic diseases that are rooted in enzyme deficiencies, safety takes precedence over efficiency. In this regard, nonviral delivery is likely to be prevailing mode of clinical application for enhancing chondrogenesis in the future, provided that the efficiency of nonviral gene transfer can be refined to approximate that of viral gene transfer. How long the transgene should be expressed to avoid unnecessary events is unclear. 23 Future investigations are warranted to provide answers to these questions and fine-tune transfer technologies.
Footnotes
Acknowledgments
This work was supported by a grant from the National Research Foundation (NRF) funded by the Korean government (2015R1A2A1A09002793).
Disclosure Statement
No competing financial interests exist.