
Abstract
Articular cartilage has a very limited capacity for repair after injury. The adult body has a pool of stem cells that are mobilized during injury or disease. These cells exist inside niches in bone marrow, muscle, adipose tissue, synovium, and other connective tissues. A method that mobilizes this endogenous pool of stem cells will provide a less costly and less invasive alternative if these cells successfully regenerate defective cartilage. Traditional microfracture procedures employ the concept of bone marrow stimulation to regenerate cartilage. However, the regenerated tissue usually is fibrous cartilage, which has very poor mechanical properties compared to those of normal hyaline cartilage. A method that directs the migration of a large number of autologous mesenchymal stem cells toward injury sites, retains these cells around the defects, and induces chondrogenic differentiation that would enhance success of endogenous cartilage repair. This review briefly summarizes chemokines and growth factors that induce recruitment, proliferation, and differentiation of endogenous progenitor cells, endogenous cell sources for regenerating cartilage, scaffolds for delivery of bioactive factors, and bioadhesive materials that are necessary to bring about endogenous cartilage repair.
Introduction
A
Microfracture procedure is one of the traditional surgical strategies that employ the concept of bone marrow stimulation in the regeneration of cartilage. In the procedure, subchondral bone is perforated in several locations to allow the migration of endogenous progenitor cells into cartilage defects. 6 These cells migrate to the microfracture site, stimulated by chemotactic signals from the microfracture site. However, the tissues generated are mostly fibrous cartilage, which has very poor mechanical properties compared to those of normal hyaline cartilage.7–9
The reasons for the failure could be attributed to an inadequate number of stem cells remaining in situ on the defect and the inability to trigger robust cell migration from underlying bone marrow. This point is substantiated by a study which showed that exogenous transplantation of autologous uncultured bone marrow-derived mononuclear cells enhanced the results of microfracture. 10 A method that directs the migration of a large number of autologous mesenchymal stem cells (MSCs) toward injury sites, retains these cells around the defect, and induces chondrogenic differentiation that will enhance the success of endogenous repair.11,12 This review focuses on the molecular and materialistic aspects of enhancing endogenous cartilage repair that mobilizes, retains, and induces chondrogenic differentiation of endogenous stem cells (Fig. 1).
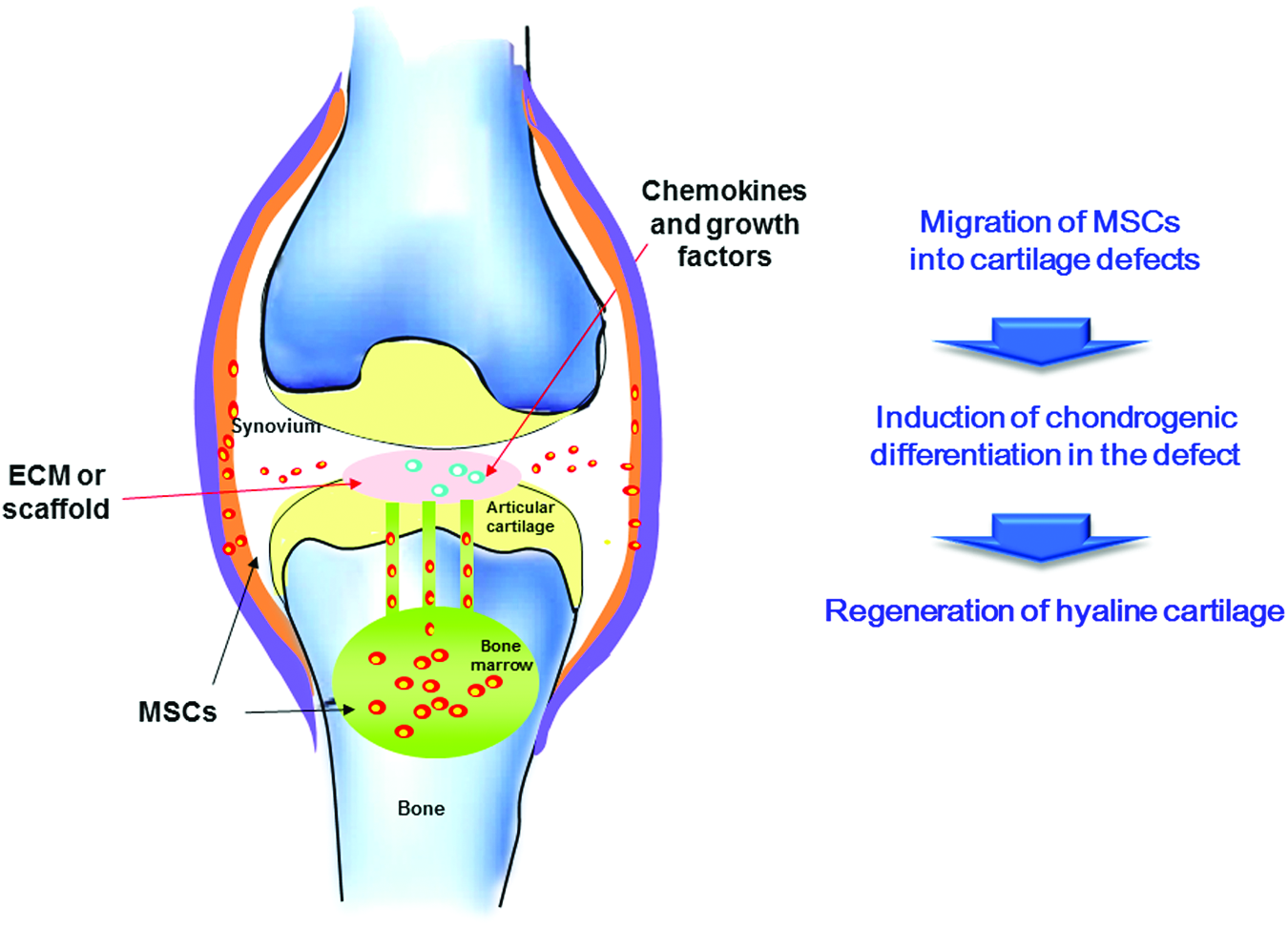
The general concept of endogenous cartilage regeneration without cell implantation. Color images available online at www.liebertpub.com/teb
Chemokines and Growth Factors for Recruitment, Proliferation, and Differentiation of Endogenous Progenitor Cells
Numerous factors are involved in the recruitment, proliferation, and differentiation of cells during cartilage repair. The presence of these factors at the local defect site promotes endogenous cartilage repair. Knowledge of the in vitro and in vivo effects of these factors is essential before employing them for the purpose.
Recruitment factors
Endogenous repair can be enhanced by recruiting more stem cells from their niche. Chemokines are small (8–10 kDa) proteins that chemically attract lymphocytes, neutrophils, and other immune cell types to inflammation sites. Several families of chemokines and their receptors exist, each with different characteristics and abilities. Chemokines attract cells directly along the chemotactic gradients from the local tissue. 13 These factors are produced endogenously by several cell types. Chemokines are regulated to maintain and repair respective tissues. As most chemokines have multiple effects, the exact cellular microenvironment determines their specific action in situ.14–16
MSCs express various receptors for chemokines17,18 and growth factors,11,19 suggesting that the receptor ligands may have a role in MSC migration. 20 These receptors are CXC chemokine receptors 1 and 2 (CXCR1 and CXCR2), CXCR4, CC chemokine receptor 1 (CCR1), CCR2, vascular endothelial growth factor (VEGF) receptor1 (Flt-1), and platelet-derived growth factor receptors a (PDGFR-a) and b (PDGFR-b). Their respective ligands are interleukin (IL)-8, stromal cell-derived factor (SDF)-1, macrophage inflammatory protein (MIP)-1α, monocyte chemoattractant protein (MCP)-1, placenta growth factor (PlGF), and PDGF. On the other hand, chondrocytes express CXCR3, CXCR4, CXCR5, and CCR6, which are receptors for the ligands interferon-γ (Mig), SDF-1α, B cell-attracting chemokine 1, and MIP-3α, respectively.21,22
SDF-1 plays a major role in the mobilization, survival, proliferation, and differentiation of stem cells. It functions as the developmental and postnatal chemoattractant for stem cells. 23 SDF-1 is upregulated at tissue damage sites, 20 and attracts MSCs. 24 Several studies have demonstrated the presence and role of the SDF-1/CXCR4 axis in MSC populations.25,26 SDF-1 has a dose-dependent effect on the migration of human and murine bone marrow-derived MSCs (BM-MSCs), as demonstrated by Boyden chamber chemotaxis assays.17,26,27 SDF-1 induces the migration of murine BM-MSCs to fracture sites in vivo, which was significantly reduced in mice treated with a CXCR4 antagonist. 27 Moreover, SDF-1-sensitive cells affect the migration of cells unresponsive to SDF-1. 28 Local treatment of low concentrations of SDF-1α causes a significant increase in recruited endogenous cells, which remain at the site for several weeks. 29
MIP-1α is the first identified of four members of the MIP-1 CC chemokine subfamily. MIP-1α causes migration of proinflammatory cells to inflammation sites and regulates their transendothelial migration. 30 MIP-1α is induced by inflammatory stimuli, mainly in cells involved in the immune response, but also in other cells, including fibroblasts and epithelial cells. 30 MSCs express CCR1, a receptor for MIP-1α. 18 The ability of MIP-1α to cause in vitro migration of BM-MSCs18,31,32 and increased expression of MIP-1α at injury sites suggests that it may be involved in MSC homing. 20
IL-8, a CXCR1/CXCR2 ligand, draws a dose-dependent chemotactic response from culture-expanded human BM-MSCs (hBM-MSCs) in vitro. Increase of IL-8 at tissue injury sites and the chemotactic response of MSCs to an IL-8 gradient 17 suggest that IL-8 may be involved in MSC migration to tissue damage sites.
MCP-1 also regulates migration and infiltration of monocytes/macrophages. Both MCP-1 and its receptor CCR2 are associated with various diseases. MCP-1 is upregulated during stress. 33
PlGF is a member of the VEGF subfamily. PlGF is a 149-amino-acid-long protein highly homologous to the PDGF-like region of human vascular permeability factor. 34 PlGF is also mitogenic for endothelial cells. 35 PlGF increases mRNA levels of cytokines (tumor necrosis factor [TNF]-α and IL-1ß) and chemokines (MCP-1, IL-8, and MIP-1ß) from cultured monocytes. 36
PDGF is a potent mitogen for mesenchymal cells, including fibroblasts, smooth muscle cells, and glial cells. 37 hBM-MSCs highly express PDGFRs.11,38 PDGF induces migration of hBM-MSCs, which is inhibited by anti-PDGF antibody. 39 PDGF is a more potent MSC chemoattractant than SDF-1 or MCP-1 in vitro. In contrast, the migration capacity of BM-MSCs in response to PDGF is not enhanced by TNF-α pretreatment, unlike chemokines. 38
We have conducted chemotaxis assays for hBM-MSCs in Boyden chambers using 48-well chemotaxis chamber plates with 8 μm pores. Seven growth factors or chemokines (IL-8, SDF-1, MCP-1, MIP-3, PDGF-AA, insulin-like growth factor-1, and hepatocyte growth factor [HGF]) were used as nonspecific chemoattractants. PDGF-AA had the most potent chemotactic effect. The number of the cells that migrated was 3.1 times higher than the negative control (0.1% bovine serum albumin [BSA]). When the dose–response effect of PDGF-AA was tested, no additional increase in MSC migration was observed above a concentration of 50 ng/mL. 11
Transforming growth factor (TGF)-β also has chemotactic capacity to adult stem cells. TGF-β3 alone recruits adipose stem cells (ASCs) and BM-MSCs, whereas codelivery of TGF-β3 and SDF-1 is chemotactic to synovial stem cells. Although SDF-1 attracts cells, it is less effective than TGF-β for all three cell types. 40 TGF-β3 induces the expression of VEGF and fetal liver kinase-1 (Flk-1), a major VEGF receptor, in mouse macrophages. 41 HGF also attracts MSCs and remains functional even after being released from collagen or fibrin gels. 42
Proliferation factors
A number of factors induce MSC proliferation. The fibroblast growth factor (FGF) family are typical growth factors for proliferation, although members of the TGF-β superfamily also have proliferative effects. 43 FGF-2 is commonly used during MSC expansion to maintain the chondrogenic potential of MSCs, while concomitantly reducing population doubling time. 44 FGF-2 markedly increases the growth rate and life span of rabbit, canine, and human bone BM-MSCs in monolayer cultures, which is more prominent in low-density cultures than in high-density cultures. 45 FGF-2 and FGF-9 differentially affect the amount of cartilage formed, depending on the stage of MSC chondrogenesis. 46 FGF-18 also has a potent proliferative effect on elastic chondrocytes and their progenitors in vivo and articular chondrocytes cultured in vitro. 47 The Wnt family of growth factors is known to interact during embryonic cartilage development to stimulate MSC proliferation. 48 Wnt3A in combination with FGF2 has been shown to support long-term expansion of human bone BM-MSCs, suggesting that Wnt3A might be a good factor to stimulate proliferation of endogenous MSCs. 49
Differentiation factors
Members of the TGF-β superfamily are most frequently used to induce chondrogenic differentiation from stem cells. 50 TGF-β1 is the crucial factor in classical chondrogenic differentiation medium, 51 whereas TGF-β3 is increasingly used in recent studies.50,52 However, both TGF-β1 and TGF-β3 similarly induced hypertrophy in MSC chondrogenesis. 53
On the other hand, TGF-β1 and TGF-β3 exhibited marked differences in their effects on chondrogenesis in posterofrontal suture-derived mesenchymal cells. TGF-β3 significantly increased cellular proliferation, whereas TGF-β1 promoted chondrogenic differentiation. 54
Bone morphogenetic proteins (BMPs), also part of the TGF-β superfamily, induce chondrogenesis from stem cells. 55 As chondrogenic induction from BMPs is an intermediate step to bone formation, BMPs should be applied with other stimuli. TGF-β and BMPs interact strongly during developmental chondrogenesis. 56 BMP-2 is the most commonly investigated factor among BMPs.57,58 The effect of BMP-2 is dependent on Sox-9 cell status; high Sox-9 expression leads to BMP-2-driven chondrogenesis, and low Sox-9 expression proceeds to BMP-2-driven osteogenesis. 59 BMP-6 or −7 are required with TGF-β, for chondrogenesis of ASCs.60,61 BMP-6 is known to induce TGF-β receptor1 expression in ASCs, thereby rendering them responsive to TGF-β.
Among other growth factors, FGF-18 promotes chondrogenic differentiation through FGFR3. 62 Growth and differentiation factor 5 also has a potent chondrogenic potential.63,64 Kartogenin, a newly found chondrogenic molecule, promotes chondrocyte differentiation in vitro and shows chondroprotective effects in vivo. Kartogenin binds filamin A, disrupts its interaction with the transcription factor core-binding factor β subunit (CBFβ), and induces chondrogenesis by regulating the CBFβ-RUNX1 transcriptional program. 65 Intraarticular injection of kartogenin also enhances the quality of full-thickness cartilage defects repair after microfracture in rabbits, with better defect filling and increased hyaline-like cartilage formation. 66
Influence of the extracellular matrix
The extracellular matrix (ECM) influences the migration potential of cells. Migration itself is a complex process, involving the polarization of the cell followed by formation of a protrusion in the direction of movement, and the cell attachment and contraction. 67 The integrins are the most important ECM receptors contributing to cell migration.68,69 When ligands bind to the extracellular portions of the integrin receptor, intracellular signaling cascades are activated that control the formation and potentiation of adhesion, cytoskeletal reorganization, and cell polarity throughout migration.68,69
ECM proteins exhibit abundant nanotopography. 70 For example, collagen fibrils present nanoscale structures which are∼300 nm long and 1.5 nm wide. 71 ECM provides directional cues to the cells and directs the motility response through anisotropy in the microenvironment, a phenomenon called contact guidance. 72 Topography at the nanoscale level has been found to alter focal adhesion size and orientation.73,74 This, in turn, changes the intracellular tension and cytoskeletal arrangement within the cell, altering cell shape and intracellular signaling. Self-renewal of MSCs requires an intermediate level of cellular tension, while adipogenic and osteogenic differentiation require a lower and higher level of cellular tension, respectively. 75
The physical properties of the ECM, such as its elasticity or stiffness, are equally important in the control of cellular processes.76,77 Engler et al. showed that a soft matrix of 0.1 kPa supported differentiation of MSCs into neuronal-like cells, a medium elasticity matrix of 11 kPa induced myogenic differentiation, and a stiff matrix of 34 kPa promoted osteogenic differentiation. 78
Endogenous Cell Source for Cartilage Regeneration
Endogenous progenitor cells from different sources respond to chemotactic signals and contribute to endogenous regeneration of cartilage.
Bone marrow
BM-MSCs are the most investigated MSC source in vitro.51,52,79–81 Microfracture is the most commonly applied surgical intervention for cartilage repair. The procedure has been used principally to repair localized chondral defects rather than diffuse lesions, such as osteoarthritis. Small holes are created on the injured articular surfaces through the subchondral bone to the marrow cavity, which allows blood and marrow components to move into the cartilage defect. 82
There is growing evidence that MSCs originate from pericytes, which are branched cells located on the outer side of small blood vessels and have close connection with the vessels' endothelial cells. 83 The proximity to vessels would allow pericytes to enter the bloodstream quickly in response to appropriate signals. 84 Pericytes exhibit in vitro phenotypes similar to that of osteoblasts, 85 and express MSC markers such as CD44, CD73, CD90, and CD105. 5
Synovium
Progenitor populations are present within the synovium86–89 and synovial fluid,90,91 and both populations possess chondrogenic potential. The exact origin of synovial fluid-derived cells is unclear, but they increase in number during injury.92,93 These synovial progenitor cells may contribute to normal joint maintenance and provide a source of cells for endogenous repair of chondral defects.94–96
During normal synovial joint development, the synovium and cartilage originate from a common pool of progenitor cells, suggesting that synovial mesenchymal progenitor cells from the synovium share a common history with the cartilage surface of the joint. 97 Significant evidence suggests endogenous chondrogenic potential of synovial cells. Chondromatosis is a condition in which cartilage forms within the synovial tissue. 98
Cartilage
Chondrocytes are the most abundant cells in AC. However, chondrocytes are encased within a rigid matrix tissue. Endogenous chondrocytes must migrate through tissue to reach the defect site to directly contribute to cartilage defect repair. The migratory capacity of chondrocytes has been demonstrated in in vitro isolated cell systems and ex vivo cartilage explant cultures, although in vivo migration remains to be determined. 99
A resident population of progenitor cells, called AC progenitor cells (ACPCs), has been isolated from the surface zone of AC. This cell population exhibits high affinity for fibronectin, possesses high colony-forming efficiency, and expresses Notch 1, which is the cell fate selector gene. Inhibiting Notch signaling abolishes the colony-forming ability of ACPCs, whereas activating Notch rescues this inhibition. 100 ACPCs have been identified in bovine, 100 equine, 101 and human tissues.102,103 High telomerase activity and maintenance of telomere length, which are characteristics of an MSC population, are observed in ACPCs. 103 ACPCs have also been demonstrated to migrate actively toward diseased cartilage. 102
A small population of progenitor cells is also present in the perichondrial groove of Ranvier in rabbits. Cells in the perichondrial groove of Ranvier are positive for several markers associated with progenitor cells and stem cell niches, including Stro-1, Jagged1, and BMPr1a. These progenitor cells may be a potential source for cartilage repair under appropriate migration, mitosis, differentiation, and matrix production signals. 104
Scaffolds and Delivery of Bioactive Factors
Scaffolds are used to contain and deliver bioactive factors, including chemokines and growth factors to local defect areas. 105 Scaffolding materials must withstand various physical forces acting in the joint before durable matrix materials are produced at the local cartilage defect site. Scaffolds must allow cells to migrate into the material. 106
Recent studies have focused on hydrogels as the scaffolding material for cartilage repair. Hydrogels are biphasic materials with properties similar to AC and support the growth and differentiation of cells into new cartilage. Chondrogenic differentiation of migrated stem cells is also influenced by the material properties of the hydrogel. A vast number of hydrogels have been developed, including natural polymers (collagen, fibrinogen, agarose, alginate, and hyaluronic acid), synthetic polymers [e.g., poly(ethylene glycol) (PEG) and its derivatives], and mixtures of natural and synthetic polymers. 107
Hyaluronan (HA)-based scaffolds are frequently used due to their biocompatibility108–110 and adaptability.111,112 HA scaffolds modify the expression profile of human MSCs by upregulating the expression of chemotactic factors CXCR4, CXCL13, and stromelysin-1 and downregulating the expression of inflammatory and degradatory factors, including CXCL12, CXCR5, matrix metalloproteinase-13, and tissue inhibitor of metalloproteinase-1. 113 Applying hyaluronic acid gel following microfractures in cartilage defects of rabbit knees results in regeneration of thicker, more hyaline-like cartilage. 114 Treatment of full-thickness ovine cartilage defects with microfracture and covering with a cell-free cartilage implant made of a poly-glycolic acid scaffold, HA and autologous serum improved cartilage repair tissue formation compared to microfracture treatment alone in the midterm outcome after 6 months. Migration of cells into or enrichment of progenitors within the cell-free implant may be induced by autologous serum that contains a variety of chemokines and growth factors. Hyaluronic acid may induce or, at least, support the chondrogenic development of mesenchymal progenitors in microfracture. 115
Collagen I has also been used as a hydrogel to release growth factors during AC repair. A collagen I scaffold releasing SDF-1 led to enhanced repair of partial-thickness defects in rabbits.116,117 Agarose can be used in a microparticle form for the delivery of growth factor 118 or as a scaffolding matrix for other drug carrier materials such as poly(ethylene oxide) (PEO)-based diblock copolymers, micelle. 119 Agarose can also be complexed with other natural polymers such as chitosan and gelatin120,121 or mixed with platelet-rich plasma (PRP) 122 to be used as a scaffolding material for cartilage repair. Besides, chitosan–glycerol phosphate–blood implants improved hyaline cartilage repair in ovine microfracture defects by the incorporation of a thrombogenic and adhesive polymer, specifically, chitosan. 123
PRP is a supraphysiological concentration of platelets and takes advantage of growth factors harbored in α-granules.124,125 In vitro, PRP is found to stimulate cell proliferation and cartilaginous matrix production by chondrocytes and MSCs, enhance matrix secretion by synoviocytes, mitigate IL-1β-induced inflammation, and provide a favorable substrate for MSCs. In preclinical studies, PRP has been used as a gel to fill cartilage defects with variable results, or to slow the progression of arthritis in animal models with positive outcomes.124,126,127 Findings from current clinical trials suggest that PRP may have the potential to fill cartilage defects and enhance cartilage repair, attenuate symptoms of osteoarthritis and improve joint function with an acceptable safety profile. 124
Bioactive factors can be delivered by being simply dispersed within the scaffold matrix. The release rate of the bioactive factor depends on the interaction between the factor and the matrix, either mediated by encapsulation, electrostatic interactions, immobilization/tethering, ECM affinity, hydrogen bonding, and/or van der Waals forces. Dense scaffold crosslinking and a small pore size can suppress the large burst release. Some natural materials have innate physical properties that control the release of bioactive factors. 128 Heparin provides a typical example. Electrostatic interactions with heparin result in the sustained release of BMP-2 in heparin-conjugated fibrin (HCF) compared to burst release of BMP-2 from normal fibrin gels. The long-term delivery of BMP-2 in HCF resulted in the regeneration of hyaline-like cartilage in the defects subjected to microfracture as opposed to short-term delivery or no BMP-2 delivery. 129 An alginate–sulfate scaffold has heparin-like affinity interactions to TGF-β1, resulting in a more sustained release of TGF-β1. 130 Our group reported that PDGF-AA is released for 21 days when loaded in HCF. We also created osteochondral defects in the distal femurs of nude rats. The defects were sealed with HCF-only or PDGF-AA-loaded HCF. The nanoparticle (NP)-labeled MSCs dispersed outside the marrow cavity within 3 days after injection in the HCF-only group, and the labeled cells moved time dependently for 14 days toward the osteochondral defect. 11
Covalent bonding to the scaffold allows longer sustained delivery. Stable localization of growth factors is achieved by crosslinking TGF-β3 onto PLGA/gelatin/chondroitin sulfate/HA scaffolds through a condensation reaction between the carboxyl groups of the hybrid scaffold and the amine groups of TGF-β3. 131 However, tethering factors to scaffolds using very strong covalent bonding, as shown by biotin–streptavidin, requires caution because the tethered factors are not released sufficiently. 132
Nature-Inspired Bioadhesive Materials to Retain Cells
Tissue adhesives are liquid or semi-liquid compounds applied for wound closure, soft tissue adhesion, and hemostasis. They are comprised of natural substances or synthetic chemicals. 133 They exist in the form of monomers, prepolymers, or noncrosslinked polymers and undergo polymerization or crosslinking to form an insoluble adhesive complex under appropriate conditions. If one of the substrates involved in adhesion is a biological body, the phenomenon is termed bioadhesion. 134
Mussels secrete adhesive materials called mussel adhesive proteins that allow them to firmly adhere to various underwater surfaces, primarily due to the presence of
Endogenous Cartilage Repair Using Chemotactic Factor and Bioadhesive Materials
In view of the fact that implantation of exogenous cell is associated with complicated quality control and regulation issues as well as increased cost, several studies have been performed to test the idea of mobilizing endogenous cells from bone marrow, synovium, or AC. Giannoni et al. devised a delivery system for TGF-β1 to repair partial-thickness cartilage defect, which consisted of encapsulation with liposome and use of collagen matrix for the controlled release of TGF-β1. The use of the system reduced the burst release of TGF-β1 by 28%. 142 The current author had fabricated a dual growth factor-releasing scaffold that contained TGF-β2 and BMP-7 immobilized to porous polycaprolactone (PCL)/F127 to enhance the healing of cartilage defect. When the scaffolds were implanted in the rabbits, the presence of growth factor significantly improved the gross appearance of the repair while the addition of ASCs to the scaffold did not affect the result. 143 A two-layer hydrogel system was developed, spatially presenting the chondroinductive TGF-β1 in one layer and the osteoinductive BMP-4 in a second layer through affinity binding to the matrix. Administration of the bilayer system into a subchondral defect in rabbits induced endogenous regeneration of AC and the subchondral bone underneath within 4 weeks, indicating that stem cells migrating into the defect were able to sense the biological cues spatially presented in the hydrogel and respond by differentiation into the appropriate cell lineage. 144 Exogenous basic FGF (bFGF) may promote AC repair by upregulating the levels of multiple growth factors. A double-layered collagen membrane sandwiched with bFGF-loaded nanoparticles between a dense layer and a loose layer was implanted into full-thickness AC defects in rabbits. In the group with the loose layer facing the surface of the subchondral bone, fast release of bFGF was observed, and early high levels of endogenous TGF-β2, VEGF, bFGF, BMP-2, −3, BMP-4 in synovial fluid were detected on day 3. 145 In a recent study, TGF-β3 and mechano-growth factor (MGF)-functionalized silk fibroin scaffolds enhanced endogenous stem cell recruitment and facilitated in situ AC regeneration in rabbits. 146 Nel-like molecule-1(NELL-1), a protein first characterized in the premature cranial suture fusion and believed to accelerate differentiation along the osteochondral lineage, was incorporated into chitosan nanoparticles and embedded into alginate hydrogels. When implanted in the rabbits, histology of NELL-1-treated defects closely resembled that of native cartilage. 147
We have devised a new design concept for endogenous cartilage regeneration without cell implantation. The best chemotactic factor, PDGF, was chosen through in vitro cell migration assay from several known factors. PDGF and TGF-β3, which not only are the best known chondrogenic factors, but also possess chemotactic activity, were loaded on HCF. HCF loaded with PDGF-AA and TGF-β3 induced migration of BM-MSCs and enhanced cartilage repair of osteochondral defects in nude rats. 11
We added water-resistant catechol-conjugated chitosan catecholamine (CHI-C) adhesive gel patch to maximize the therapeutic effect. This hydrogel blocks the diffusion of protein and cells. This system has the following characteristics. First, anisotropic is the direction of release of the therapeutic proteins encapsulated within a hydrogel depot. The adhesive CHI-C gel patch on top of the fibrin gel allows encapsulated chemotactic factors (PDGF-AA and TGF-β3) to be released only in the direction of bone marrow, which maximizes MSC migration in a given concentration of growth factors. Second, loss of the migrated cells from the disease site is prevented by providing a physical barrier with tissue adhesive properties. Migrated cells are retained on the defect site by the adhesive chitosan–catechol barrier. The results showed that this adhesive gel patch greatly promotes effective recruitment of hMSCs. Moreover, the adhesive barrier prevents further migration and dispersion of hMSCs that otherwise would not be properly located at the disease site. In vivo imaging and macroscopic histological assessments demonstrated significant improvements in cartilage tissue (Fig. 2). 12
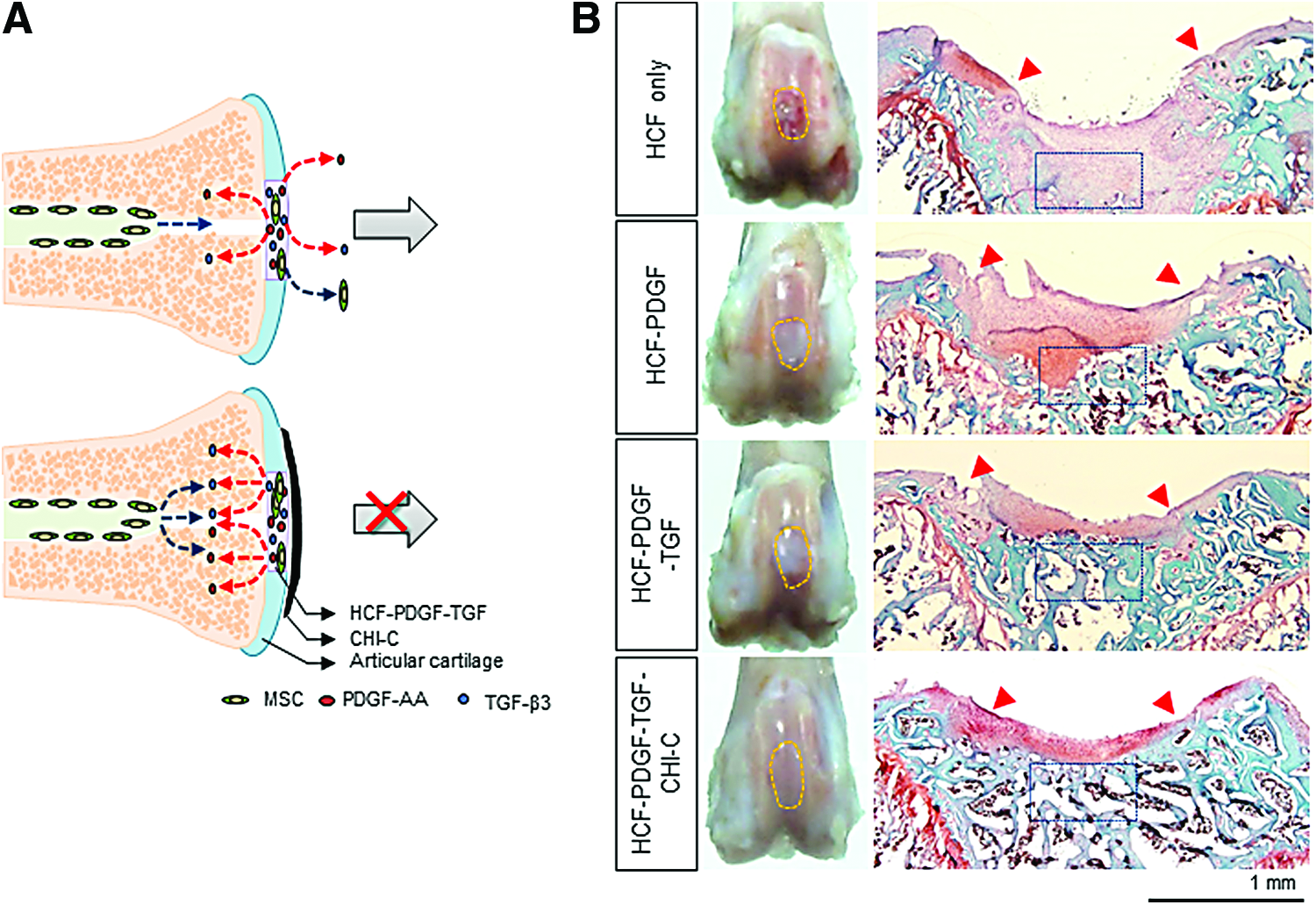
The author's method for endogenous cartilage regeneration using chemotactic factor, differentiation factor, and bioadhesive materials.
Summary and Future Perspectives
Cartilage repair and regeneration techniques do not represent a standard medical care widely adopted by average orthopedic surgeons at this time. However, these procedures will eventually evolve to better address traumatic cartilage defects with expanding market for the techniques. They will be developed to treat early osteoarthritis in the long term.
Cell therapy for cartilage defect has been studied and practiced for more than two decades. Autologous cell implantation is burdened with the high cost of the procedure and the delay associated with cell culture. Allogeneic cells make more sense commercially than autologous cells, but approval from regulation agencies for clinical applications is challenging due to safety and quality control issues. Achieving endogenous cartilage repair with this “cell-less” approach may possibly offer a much less expensive and more reproducible method for clinical application. Attracting a greater number of cells by chemotaxis, retaining recruited cells with bioadhesive material, and chondrogenic differentiation of the retained cells using growth factor can be achieved in one system. Arthroscopic application of the system will facilitate the surgical procedure and minimize the morbidity to the patients.
However, several issues must be addressed in this system before considering clinical application. The long-term toxicity of bioadhesive materials, such as the catecholamine polymer, needs to be investigated in large animal models. Ideally, the factor must be selective for MSCs, but not attract endothelial cells. Inflammatory and endothelial cells are also recruited by chemotactic factors. Prolonged presence of inflammatory cells, such as neutrophils or lymphocytes, can delay regeneration of cartilaginous tissue. Recruiting endothelial progenitor cells is potentially a critical issue during AC regeneration because angiogenesis leads to endochondral ossification. While PDGF we used is more chemotactic for MSCs than chemokines, this factor also recruits unwanted types of cells. Screening and searching for small molecules that are selective for MSCs may provide a direct answer to this problem.
Footnotes
Acknowledgment
This work was supported by a grant from the National Research Foundation (NRF) funded by the Korean government (2015R1A2A1A09002793).
Disclosure Statement
No competing financial interests exist.