
Abstract
Purpose of Review:
Tissue regeneration requires proper vascularization. In vivo studies identified that the endothelial colony-forming cells (ECFCs), a subtype of endothelial progenitor cells that can be isolated from umbilical cord or peripheral blood, represent a promising cell source for therapeutic neovascularization. ECFCs not only are able to initiate and facilitate neovascularization in diseased tissue but also can, by acting in a paracrine manner, contribute to the creation of favorable conditions for efficient and appropriate differentiation of tissue-resident stem or progenitor cells. This review outlines the progress in the field of in vivo regenerative and tissue engineering studies and surveys why, when, and how ECFCs can be used for tissue regeneration.
Recent Findings:
Reviewed literature that regard human-derived ECFCs in xenogeneic animal models implicates that ECFCs should be considered as an endothelial cell source of preference for induction of neovascularization. Their neovascularization and regenerative potential is augmented in combination with other types of stem or progenitor cells. Biocompatible scaffolds prevascularized with ECFCs interconnect faster and better with the host vasculature. The physical incorporation of ECFCs in newly formed blood vessels grants prolonged release of trophic factors of interest, which also makes ECFCs an interesting cell source candidate for gene therapy and delivery of bioactive compounds in targeted area.
Summary:
ECFCs possess all biological features to be considered as a cell source of preference for tissue engineering and repair of blood supply. Investigation of regenerative potential of ECFCs in autologous settings in large animal models before clinical application is the next step to clearly outline the most efficient strategy for using ECFCs as treatment.
Introduction
C
The efficiency of cell-based tissue regeneration and complete restoration of functionality relies on the cell type that (1) has to possess the ability to induce proper establishment of a new vascular bed; (2) exhibits a behavior that evokes a regeneration-related switch of the phenotype of tissue-resident cells or differentiation of resident stem/progenitor cells; or (3) possesses a feature that can be modulated by gene manipulation in such way that it will further enhance the regenerative potential of the cell or can be used as a gene delivery vehicle to modulate the behavior of other cells. Mesenchymal stem cells are fulfilling some of the aforementioned criteria and at present are the most often used cell type for tissue regeneration and application in disease treatment in preclinical animal models 3 and clinical trials. 4 However, the lack of an intrinsic propensity for formation of new blood vessels seriously hampers their applicability especially in tissue engineering. 5 The emergence of endothelial colony-forming cells (ECFCs), a subset of circulating endothelial progenitor cells with a high proliferative potential, has opened new modalities for cell-based therapies and tissue engineering.
This survey discusses the reported findings from in vivo studies that aimed to explore the potential use of ECFCs to induce neovascularization, to facilitate new tissue formation during tissue regeneration, or to target diseased tissues specifically.
What Are Endothelial Colony-Forming Cells?
ECFCs originate from progenitor cells present in the peripheral or umbilical cord blood and, once isolated in culture, they possess all phenotypical and functional characteristics specific for endothelial lineage. These cells also referred as late outgrowth endothelial progenitor cells (late EPCs) or blood-outgrowth endothelial cells differ from the early outgrowth CD14+/CD45+ EPCs (early EPCs), which originate from the myeloid-monocytic lineage and participate in the processes of vascular regeneration in a paracrine manner. 6 In contrast to early EPCs, ECFCs possess robust proliferative and clonal ability granting generation of sufficient number of cells for clinical application in a relatively short period. Most importantly, these cells incorporate into the endothelial lining of newly formed blood vessels and exhibit pronounced vascularization ability in vivo. 6 Especially, their neovascularization potency is augmented when ECFCs are used in combination with stem/progenitor cells, such as mesenchymal stem cells or adipose tissue-derived stem cells. 7 In the past decade, many in vivo studies outlined ECFCs as a promising cell type for new cell-based improvement of disease outcome. They regard different animal models of ischemic diseases as well as animal studies for validation of novel tissue-engineered concepts for tissue regeneration and reconstruction. However, little progress was offered with respect to the translation of those proof-of-concept findings into clinical settings.
Notwithstanding, ECFCs have been successfully isolated from healthy donors and patients8–11 paving the path for use of these cells as a cellular platform to study the pathophysiology of certain diseases as well as for autologous treatment of various pathologies.
In this review, we use the term ECFCs for the entire spectrum of names by which these cells are known. It specifies peripheral- 12 or cord blood- 13 derived or tissue-derived 14 cells belonging to the endothelial lineage, which possess robust proliferative and clonal ability that exhibit in vitro and in vivo angiogenic capacity and express endothelial cell markers such as CD31, CD146, CD309, CD144, vWF, eNOS and are negative for hematopoietic (CD45, CD14) or mesenchymal stem cell markers (CD70, SMA). 6 In addition, the clinical applicability of reported findings was a criterion for this survey, thus excluding in vivo studies that were performed with materials that pose safety concerns and have little application perspective for patients.
Therapeutic Use of ECFCs in Neovascularization and Tissue Regeneration
Accumulated evidence indicates that ECFCs can exert their therapeutic potential (1) by physical incorporation into neovessels of the target tissue, thus improving oxygenation and delivery of nutrients into the affected area 15 ; (2) in a paracrine manner by creating a niche for differentiation of stem/progenitor cells toward needed somatic cell type16,17; or (3) if genetically manipulated, to skew the disease progression toward a more positive outcome. 18 A large pool of in vivo studies reported that the intrinsic neovascularization ability of ECFCs underlies the restoration of blood perfusion in ischemic tissue in the mouse hind limb model,19,5 sustained the survival of tissue-resident somatic cells and improved hemodynamic parameters after myocardial infraction,20,21 or induced symptom relief in case of stroke. 22 Embedded into biocompatible materials, ECFCs have been used to create preformed vascular networks within tissue-engineered scaffolds that efficiently interconnected with host vasculature and facilitated tissue regeneration after in vivo implantation. 23
Recent advances in the field of regenerative medicine introduced the concept to use ECFCs, often independently of their neovascularization features, for treatment of diseases such as diabetes mellitus, 24 cancer, 25 or hemophilia A. 26 These latter aspects of ECFC applications are discussed below (Other Approaches that Utilize ECFCs for Tissue Regeneration Section Genetically Modified ECFCS as a Vehicle for Treatment Section). The next sections focus on the therapeutic approaches that hold promise to use ECFCs for tissue repair and regeneration. Two major applications forms have been used in vivo, namely, cell bolus into the systemic circulation or targeted tissue, or embedded in biocompatible scaffolds. Since the identification of ECFCs, most research that aimed to improve regeneration using the neovascularization ability of ECFCs was performed by injecting a cell bolus alone or in combination with other cell types either into systemic circulation or directly into targeted tissue affected by ischemia. In vitro or in situ prevascularization of engineered biocompatible scaffold by ECFCs represents an alternative approach using the neovascularization ability of ECFCs for tissue engineering assisted regeneration of tissues and organs. Both approaches can be used to deliver modified ECFCs as a vehicle for gene therapy (Fig. 1).
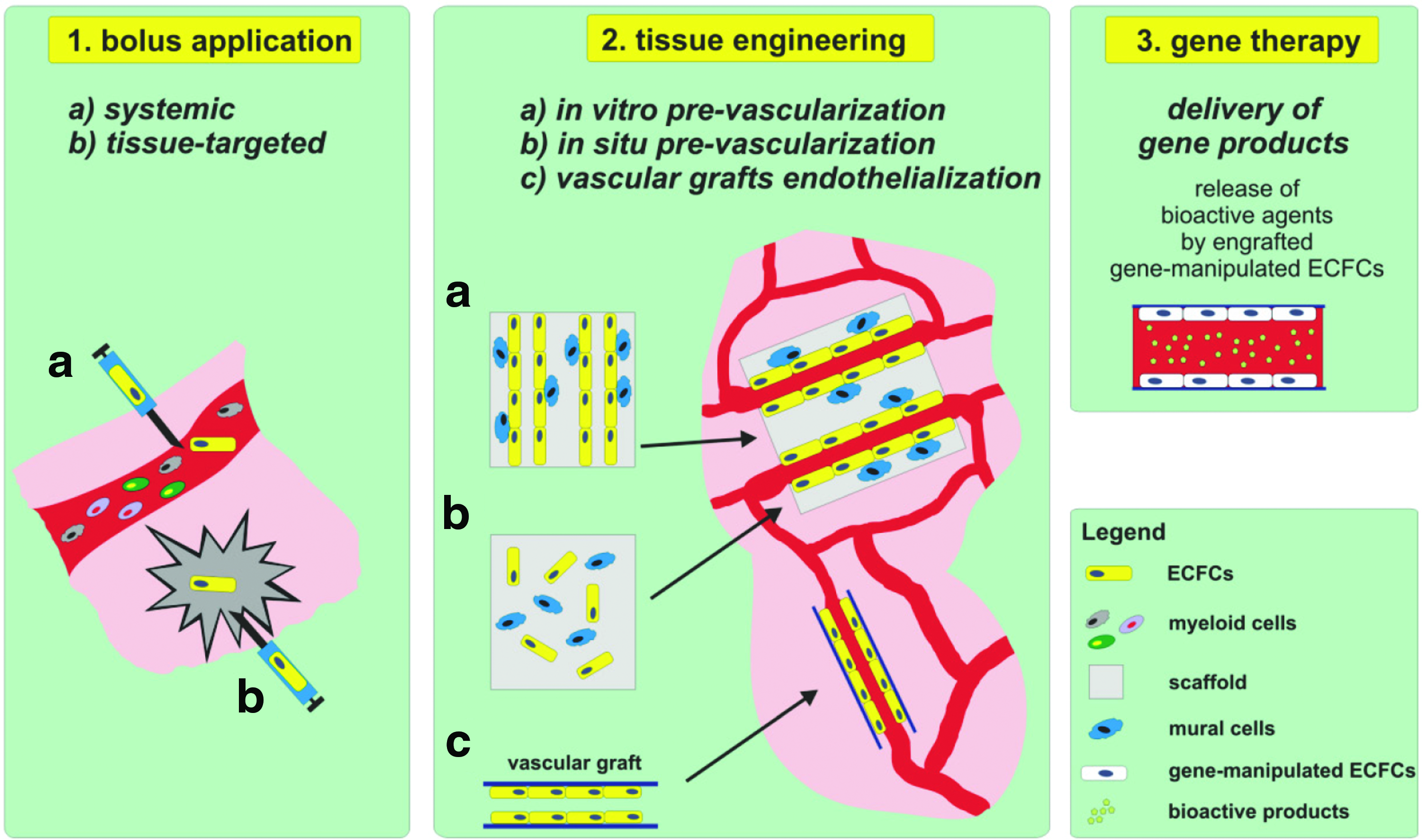
Application of ECFCs for tissue regeneration and repair. Data obtained from in vivo studies delineated three main approaches of use of ECFCs for regenerative purposes. Tissue regeneration and repair can be accomplished by injecting ECFCs either into systemic circulation (
Bolus Application of ECFCs
Systemic application of ECFCs induced neovascularization-dependent tissue regeneration in animal models of peripheral ischemia, 5 vessel damage,27,28 traumatic brain injury, 29 stroke, 22 or lung-related diseases. 30 However, the encouraging results from those studies were blunted by the observation that intravenous bolus application of ECFCs was accompanied by entrapping the ECFCs in tissues that were not primary targets 31 and low engraftment and short survival rate into the targeted area. 5 Furthermore, those studies did not provide further insight in the mechanism of how ECFCs participate in neovascularization and tissue regeneration. Thus, intravenous bolus injection of ECFCs has drawbacks that impair the therapeutic efficacy of these cells, suggesting that this application route is not the most efficient approach for administration of ECFCs.
Targeted application of a cell bolus into the affected tissue circumvents the low engraftment of ECFCs and imposes greater control over the systemic distribution of the cells and the cell number needed to establish the desired therapeutic effect. Studies performed with rodent animal models of myocardial infarction,20,32,33 traumatic brain injury, 34 or retinal ischemia 35 reported promising improvement of organ function and increased vascularization after application of ECFCs by intramyocardial, intracerebroventricular, or intravitreal injection, respectively.
Supporting the angiogenic efficacy of bolus injected ECFCs
The initial animal studies where ECFCs were applied as bolus cell injection for treatment of pathological conditions pointed to the necessity to develop other strategies that will increase the therapeutic potency of ECFCs by using stem or progenitor cells in addition to ECFCs or to modulate the ECFC function by pretreatment with soluble bioactive components, environmental clues, or gene transfer.
Coimplantation of ECFCs with cell types that can support neovascularization by acquiring mural cell phenotype and stabilize the newly formed vasculature resulted in even greater regeneration benefits. A combination of ECFCs with adipose tissue-derived stem cells (ADSCs), 36 MSCs, 7 or SMCs37,38 significantly improved the neovascularization score by forming functionally long-lasting new blood vessels that are interconnected with host vasculature, alleviated symptoms, and improved overall organ or tissue functionality in different models of ischemia. Interestingly, similar results were obtained when ECFCs were coimplanted with host myeloid cells. 39 While in cell therapy studies, ECFCs were indispensible for neovascularization, the accessory cells are not, as already reported for MSCs 5 and myeloid cells. 39
Support of ECFC-mediated vascularization by growth factors and hypoxia
Augmentation of the therapeutic ability of ECFCs can also be achieved by exposing the cells to prosurvival or proangiogenic trophic factors or exposing the cells to environmental conditions that usually are present in ischemic areas. Pretreatment of ECFCs with the chemokine SDF-1, 40 fucoidan,19,41 the formyl peptide receptor-2 agonist WKYMVm, 42 and erythropoietin 43 improved hind limb blood flow and capillary density, and showed less necrosis and better muscle preservation than the animals treated with control ECFCs in the mouse hind limb ischemia model. Intramyocardial application of ECFCs pretreated with a very low concentration of genistein (10−10 M) induced cellular proliferation and secretion of angiogenic cytokines at the ischemic sites of a murine acute myocardial infarction model. 20 It was accompanied by enhanced capillary formation, decreased myocardial fibrosis, and improved overall cardiac function over a period of 28 days. 20 Stimulation of ECFCs with adiponectin significantly improved hind limb perfusion in streptozotocin-induced hyperglycemic nu/nu mice. 44 Most importantly, pretreatment of ECFCs isolated from diabetes mellitus patients with adiponectin completely reversed their reduced neovascularization ability in vivo. Although the duration of this improvement needs further evaluation, this pretreatment offers a novel and promising approach to counteract the functional impairment of ECFCs in diabetic patients.
Hypoxia, acidic environment, as well as abundance of fibrinous exudate are usual components of an inflammatory milieu. Preconditioning of ECFCs with these environmental clues before their use as a therapeutic means to induce regeneration is an interesting approach for increasing of their angiogenic efficacy. Exposure of ECFCs to 1% 45 or 2% O2 46 up to 24 h enhanced the proliferation and survival of transplanted ECFCs, and triggered a proangiogenic cytokine secretion at ischemic sites through HIF-1α/TWIST 45 and the JAK2/STAT3 axis. 46 These cellular effects improved the blood flow ratio and the capillary density in ischemic mouse hind limb. Similar results of improved neovascularization into ischemic tissue driven by activation of AKT and ERK1/2 have been reported after pre-exposure of ECFCs to the pH 6.6 acidic environment 47 or shortly cultured in the presence of human platelet lysate. 48
Increased survival and proangiogenic ability by activation of either AKT or the JAK2/STAT3 axis have also been used as an approach to genetically modulate these pathways in ECFCs to increase their therapeutic efficacy. Restoring the Akt1 activity in ECFCs from South Asian men rescued their inherited reduced vascular reparative potential in mouse hind limb ischemia, 49 while ex vivo transduced ECFCs with Akt and hemoxygenase-1 have improved cardiac performance and reduced negative remodeling after myocardial infarction in nude mice. 33 Furthermore, selective silencing of Lnk adaptor protein activated the JAK2/STAT3 pathway in ECFCs, which accelerated angiogenesis and promoted the tissue repair in ischemic hind limb. 50 Therefore, strategies that increase the therapeutic efficacy of ECFCs offer a way to circumvent drawbacks of bolus application of ECFCs in clinical settings and might provide better therapeutic outcome. Notwithstanding, bolus application of ECFCs remains a clinically relevant application route especially for treatment of pathological conditions when a targeted intratissue use is the only way to introduce cell-based therapy, such as in myocardial infarction, or by systemic application in intimal hyperplasia, 28 or lung repair. 30
Use of ECFCs in Prevascularized Tissue-Engineered Constructs
ECFCs can also be used as a means for generation of prevascularized tissue-engineered constructs. This approach offers greater control of the microenvironmental components that govern blood vessel assembly. It also provides the possibility to introduce other cell types that either facilitate angiogenesis or differentiate toward tissue-specific cell lineage. Furthermore, it allows to modulate the biological activity of ECFCs by changes of the structure of biocompatible scaffold or by addition of trophic mediators that support neovascularization and regeneration. Earlier studies with ECFCs embedded in scaffold made of Matrigel 51 showed that the cells formed functionally long-lasting and host-interconnected vascular networks in vivo especially in combination with MSCs 7 or ADSCs. 36 These findings provided a platform for researchers in the tissue engineering field to start evaluating the potential of ECFCs for tissue regeneration and reconstruction in conditions that can be applied in clinical practice for treatment of different pathological conditions.
Effect of scaffold materials on neovascularization properties of ECFCs
A tissue-engineered graft essentially consists of tissue cells supported by a (bio)matrix. Both natural biomatrices and biocompatible synthetic matrices can be used, some of which even display self-organizing properties toward the embedded or surrounding tissue cells. Both type of materials are relatively safe for clinical use, mimic structural and functional properties of the natural extracellular matrix, and can be modulated by physicochemical processes to improve the biomechanical properties of the scaffold to maintain cell function, the development of the vascular network, and tissue regeneration. 52 The in vivo research studies that evaluated the potential of ECFCs for tissue engineering are discussed in the next sections according to the use of natural or synthetic biocompatible materials.
Natural materials, either chemically modified or combined with peptides
When embedded in hyaluronan acid (HA),53–55 gelatin, 56 alginate, 57 collagen or fibrin hydrogels,58–61 ECFCs exhibited potent neovascularization effects. Nevertheless, not all natural biopolymers are suitable for tissue engineering. As reported previously, gelatin lacks mechanical stability, 62 alginate by itself has limited intrinsic propensity to facilitate angiogenesis because of uncontrolled degradation, 63 while ECFCs embedded in HA hydrogels exhibited similar neovascularization effects as the empty control scaffolds. 55 However, addition of integrin-binding RGD peptides and VEGF121 or HGF57,64 to alginate gels created a permissive proangiogenic environment, which resulted in increased neovascularization driven by ECFCs after intramuscular application of the scaffold in the mouse ischemic hind limb.
Similarly, addition of RGD- and MMP-sensitive peptides to HA scaffolds laden with human ECFCs resulted in vascular network formation in tumor-bearing mice composed of 60% of the blood vessels containing both human ECFCs and host endothelial cells, while the remaining vessels contained only human ECFCs. 55 The newly formed vascular bed in ECFC-laden scaffolds anastomosed with the host's circulation and supported blood flow in the hydrogel after 14 days of transplantation, while human microvasculature could not be detected in implants of HA hydrogels without ECFCs.
Chemical modification of biomaterials can also influence the behavior of ECFCs in the matrix. Addition of vinyl methacrylate groups to gelatin gels (GelMA) not only induced neovascularization in implants laden with ECFCs and MSCs in immunodeficient mice but also revealed that the extent of vascular network formation was different between the GelMA constructs with final concentrations of 1% and 10% methacrylate. 65 In the 1% GelMA construct, numerous blood vessels containing erythrocytes were uniformly distributed throughout the implants, while there were fewer perfused blood vessels in the 10% GelMA constructs with microvessels mainly located at the periphery of the constructs. Gelatin-phenolic hydroxyl gels loaded with ECFCs and ADSCs induced a different extent of in situ vascularization, which was dependent on the concentration of H2O2 that was used for enzymatic crosslinking. 56 The number of lumens inside the construct decreased with increasing the H2O2 concentration that was in direct correlation with the reduced proteolytic degradability of the scaffold in nude mice. 56
Taken together, by changing the biochemical and physical properties of the local microenvironment within the scaffolds composed of natural polymers, it is possible to influence the vasculogenic ability of ECFCs and to guide neovascularization in vivo as was also reported for collagen scaffolds.66,67
Synthetic materials, combination of them or modified by physicochemical procedures
Synthetic polymers such as poly-
ECFCs embedded within scaffolds composed of synthetic polymers were used successfully to improve neovascularization and tissue regeneration. ECFCs embedded in heparin-immobilized PCL scaffolds loaded with VEGF induced significantly high density of blood vessel formation after 7 days of subcutaneous implantation in immunodeficient mice compared to empty scaffolds. 74 Subcutaneously implanted polycaprolactone–tricalcium phosphate (PCL-TCP) scaffolds loaded with ECFCs and human fetal MSCs induced robust neovasculogenesis and ectopic bone formation in vivo after 12 weeks. 16 Besides neovascularization, ECFCs demonstrated an osteoinductive effect by increasing the bone formation ability of human fetal MSCs through BMP/TGF-β signaling. Robust neovascularization accompanied by increased bone induction was also reported when ECFCs were embedded with bone marrow-derived MSCs in hydroxyapatite (HA)-polylactide-co-glycolide (PLG) scaffolds and applied for treatment of orthopic calvarian defect in albino nude rats. 75 Subcutaneous application of ECFCs combined with primary osteoblasts into starch polycaprolactone fiber meshes scaffold also induced bone formation. 76 An interdependent relationship between increased neovascularization and efficient osteogenesis was also reported when ECFCs alone were loaded into βTCP coated with fibronectin in a nude rat calvarial model. 70 Similar results were reported when ECFCs were embedded into BCP constructs enriched with BMP-2-induced bone formation in immunocompetent mice. 74 These studies indicate that ECFCs are suitable for bone repair.
In a mouse model of full-thickness excision wound, transplantation of ECFC-seeded RGD-g-PLLA scaffold resulted in better neovascularization compared to empty control scaffolds. 71 In addition, the ECFC-seeded RGD-g-PLLA scaffolds promoted superior dermal wound healing when compared with the intradermal ECFC injection indicating that the scaffold application route of ECFCs significantly improved cell localization, survival, and retention in the injured tissue (Table 1).
Early studies that used ECFCs for regenerative purposes emphasized the need to investigate and adopt different strategies to increase the therapeutical potential of ECFCs as well as to expand the list of possible indications for treatment of pathological conditions. (1) Pretreatment of ECFCs with bioactive compounds or factors increased the cell survival, tissue engrafting, as well as angiogenic capacity of ECFCs. (2) Gene manipulated ECFCs exhibited better therapeutical effect than source cells. (3) ECFCs combined with other cell types have shown facilitated angiogenesis as well as tissue regeneration and repair. (4) Embedding of ECFCs in natural and synthetic biocompatible scaffolds enables efficient neovascularization and induction of tissue regeneration and repair.
βTCP, β-tricalciumphosphate; BCP, biphasic calcium phosphate; ECFC, endothelial colony-forming cell; EPO, erythropoietin; PCL, polycaprolactone; PLGA, poly-
Other approaches that utilize ECFCs for tissue regeneration
In addition to their vasculogenic potential, ECFCs can also have a direct regenerative effect by secreting soluble regulators that govern tissue repair. ECFCs combined with tissue or organ derivatives such as pancreatic islands, skin sheets, or decalcified bone were also used for tissue-engineered regeneration.
Intraportal transplantation of ECFC-covered porcine pancreatic islets exhibited better graft function and survival, reduced inflammatory response, and better regulation of increased glucose levels in diabetic nude mice. 77 Reduced graft loss and dysfunction were reported when ECFCs were cotransplanted with a rat marginal islet graft under the kidney capsule of hyperglycemic NOD/SCID mice. 24 In vivo cell tracing demonstrated that ECFCs remained at the site of transplantation inducing beta cell proliferation, which was accompanied with improvement of glycemia and increased plasma C-peptide compared to control animals. ASCs and ECFCs combined with either freshly isolated mature human adipocytes or pig pancreatic islets in collagen/fibronectin gels established a functional vascular network after 2 weeks of subcutaneous application at the flank region of NOD/SCID mice. 36 The implants loaded with adipocytes exhibited typical morphology of vascularized adipose tissue, while the implants carrying islets contained vascular networks with clusters of cells staining positive for insulin. These studies support the use of autologous ECFCs in translational studies that aim to improve current islet transplantation protocols for the treatment of type 1 diabetes.
In another study, ECFCs were embedded together with MSC-derived smooth muscle cells in collagen/fibronectin spheroids and loaded into porous decalcified bovine cancellous bone (PBCB, Tutobone) for treatment of calvarial critical size defects in the SCID mouse model. 78 Spheroids composed of ECFCs alone or in combination with smooth muscle cells were unable to form blood vessels, but once loaded into PBCB triggered in growth of host blood vessels into the implants, which correlated positively with bone formation within the implants. Furthermore, incorporation of ECFCs into tissue-engineered human skin substitutes resulted in formation vascular conduits enabling perfusion and survival of human bioengineered tissue, 79 while the combination of dermal fibroblast sheets with ECFCs induced robust vasculogenesis, adequate epithelial coverage, and normal matrix organization, which prevented excessive wound contraction. 80
Endothelialization of synthetic small-diameter vascular grafts using ECFCs opens another new venue for application. The pitfalls of previous studies performed with other endothelial cell types such as umbilical vein endothelial cells were mainly related to graft failure due to intimal hyperplasia or thrombus formation followed by vessel occlusion. 81 ECFCs attenuate intimal hyperplasia contributed by mesenchymal stem cells after vascular injury in a mouse femoral wire injury model, 28 but a modulating effect has still to be evaluated in appropriate vascular grafts. In animal studies, this type of application has been combined with gene transfer to improve anticoagulant properties. To that end, control ECFCs or ECFCs transfected with an adenoviral vector containing the gene for human thrombomodulin (ECFCs + AdTM) were sodden into the lumens of small-diameter ePTFE vascular grafts, which were implanted into femoral arteries of athymic rats. 82 After 28 days, the patency rate of control ECFCs and ECFCs + AdTM was 75–88%, while intimal hyperplasia was observed near both the proximal and distal anastomoses. However, these regions of anastomoses were negative for human vWF staining indicating that the host cells, but not the ECFCs, contribute to this negative side effect of graft implantation. 82
Published literature indicates that ECFCs affect the differentiation of stem/progenitor cells toward a somatic cell type in a paracrine manner by releasing soluble trophic factors such as BMPs, TGF-β, PDGF-BB, VEGF, or angiopoietins.16,17 Recent findings suggest that exosomes represent a paracrine mechanism through which ECFCs also exert a therapeutic effect. 83 These microvesicles, with a diameter ranging from 30 up to 100 nm, are involved in cell–cell communication as vehicles that transfer information (mRNA, miRNA, DNA)84,85 as well as proteins 86 between the cells. Intravenous application of exosomes derived from ECFCs attenuated renal injury in a mouse model of experimental acute kidney injury (AKI) by reducing plasma creatinine, tubular necrosis, macrophage infiltration, oxidative stress, and apoptosis. 83 Therefore, the use of exosomes released from ECFCs under stringent in vitro conditions represents a promising therapeutic modality for tissue regeneration that warrants further investigation in different animal models of diseases.
These studies emphasized that ECFCs possess desirable tissue regenerative attributes that are not only strictly dependent on the cells' neovascularization ability but also on the trophic capacity to create a microenvironment that induces a proregenerative phenotypical switch in tissue-resident somatic cells.
Genetically Modified ECFCs as a Vehicle for Treatment
While ECFCs possess appealing therapeutic potential by virtue of their ability to induce neovascularization and release of trophic factors that induce regeneration into affected tissue, these cells also represent an attractive cell source for vehicles for gene therapy.
ECFC-based gene delivery strategy was used as a feasible technology for administration of recombinant proteins in case of anemia or hemophilia A. It was shown that ECFCs transfected with a nonviral plasmid carrying complementary DNA for modified human coagulation factor VIII predominantly accumulated in bone marrow and spleen of NOD/SCID mice, from where efficiently secreted FVIII into circulation achieving therapeutic levels relevant for clinical treatment of hemophilia even after 5 months. 26
Similarly, genetically engineered ECFCs able to express controlled delivery of erythropoietin (EPO) were embedded in collagen/fibrin gels together with BM-MSCs and subcutaneously implanted in a nephrectomy-induced kidney failure mouse model or in a murine model of radiation-induced anemia. 18 ECFC-EPO combined with MSCs formed extensive vascular networks interconnected with the host vasculature and once engrafted, the cells expressed and secreted functional EPO in vivo. After 5 weeks, the mice with implanted ECFC-EPO have shown enhanced hematocrit, RBC count, hemoglobin concentration, reactive splenomegaly, and downregulation of endogenous EPO expression in the kidneys.
Systemic application of ECFCs encoding cytotoxic or inhibitory gene products has been used to induce antitumor responses in animal models of glioma, lung, or pancreatic tumors. ECFCs expressing antiangiogenic factors such as soluble VEGFR-1 (sVEGFR-1) and angiostatin–endostatin (AE) fusion proteins induced favorable disease outcome by reducing the tumor size and growth in C3TAG mice with spontaneous mammary tumors and in mice bearing established lung or pancreatic tumors, while in an orthotopic model of glioma, authors reported doubling of the median survival of mice treated with sVEGFR-1. 87 In Lewis lung carcinoma, tumor growth was suppressed only after the application of combination of cells producing sVEGFR-1 or AE proteins. Replicating oncolytic viruses such as measles viruses of the Edmonston B strain (MV-Edm) have shown promise for treatment of cancer. Interestingly, ECFCs express CD46 and can be easily infected by MV-Edm while being resistant to MV-Edm-mediated cell death and protected virus replication from host immune response. Infected ECFCs releasing attenuated measles viruses of the Edmonston B strain (MV-Edm) efficiently contaminated the U87 glioma cells leading to focal infection and cytopathic effects that reduced tumor size and prolonged survival of mice. 25 The above given studies showing that ECFCs can be attractive cellular vehicles for gene delivery.
Autologous Implementation of ECFCs in Nonmouse Models
Tissue regeneration is a complex yet finely tuned spatiotemporal process of continuous interaction between immune cells, vascular and tissue-resident stem or progenitor cells that actively occurs during lifetime. It is markedly accelerated after tissue injury. Exclusion of one component of this system in experimental animal models could lead to misleading interpretation of collected data compromising the clinical translation of reported proof-of-concept finding. Most of our knowledge about ECFCs is gathered from studies performed in immunocompromized murine animal models. However, interaction of implanted human ECFCs and immune cells showed that this interaction is crucial for regeneration. 39 Lack of immune response in immunocompromized animals interferes with evaluating the immunogenicity of ECFCs in vivo as well as the interplay between ECFCs and immune cells. Only one study reported low immunogenicity of human PB-derived ECFCs in a rat skin transplantation model with immunocompetent WKY rats, 88 which warrants further investigation of ECFC behavior in autologous or other intact animal models.
Other factors also influence the evaluation of therapeutical efficacy of ECFCs in murine animal models. Concomitant pathologies often present in mice models, such as diabetes mellitus in NOD mice, hinder the clear extrapolation of collected knowledge for development of new treatments based on the use of ECFCs. Many examples showed that ECFCs can be used to treat peripheral ischemia in a hind limb ischemic mouse model. However, the lack of an explanation whether the observed beneficial effect of ECFCs is due to direct involvement in angiogenesis or paracrine stimulation of arteriogenesis hampers the objective evaluation whether the obtained data are clinically relevant or not. In addition, despite its large flexibility, the mouse is less suitable for isolation and evaluation of the use of autologous ECFCs for disease treatment as its blood volume is relatively small. Therefore, these simplified murine animal models are unlikely to accurately represent the in vivo performance of ECFCs in humans.
Successful isolation of ECFCs from larger than murine animals offers an opportunity to test ECFCs in the autologous environment of intact organism. This approach offers a more reliable and accurate insight of how ECFCs behave during neovascularization and tissue regeneration. It may be preferable to start with a medium-sized animal, such as rat or rabbit, to test the feasibility of the treatment and to work out the eventual technical problems, and then to investigate the same concept in large animal models.
ECFCs have been successfully isolated and characterized from peripheral blood or organs of rats, 89 rabbits, 90 pig, 91 dog, 92 sheep, 93 goat, 94 as well as nonhuman primate 95 opening new ventures to investigate the possibility of using ECFCs for treatment of different pathological conditions in autologous settings. Isolated cells from different animals were compliant with the present criteria for characterization of ECFCs such as belonging to endothelial cell lineage, absence of markers characteristic for HSC or MSC, robust proliferation, and clonal ability, as well as in vitro and in vivo neovascularization capability. 15 Application of autologous ECFCs promoted bone fracture repair and bone regeneration,96,97 attenuated the effect of acute lung injury,30,98 and rescued infracted myocardium99–101 in different disease models in rat. ECFCs might represent a preferential cell source for endothelialization of vascular grafts 102 and for treatment of cardiovascular-related 90,102–104 or lung-related 105 pathologies as reported in various disease models in rabbit. Subsequent in vivo testing in large and relevant animal models offers a better opportunity to evaluate the performance of ECFCs in clinically relevant conditions, that is, closely resembling human pathophysiology. Autologous transplantation of ECFCs has shown promising results in the treatment of different pathological conditions in porcine,91,106–111 canine,92,112–116 ovine,93,117,118 and goat 94 animal models. Published reports of the beneficial role of autologous implanted ECFCs in large animal models also offer an opportunity for developing in vitro assays and markers that might have a scientifically sound correlation with clinically relevant parameters, while at the same time can monitor and predict the performance of ECFCs in vivo.
Conclusion and Further Perspectives
The research performed after the identification of ECFCs 12 suggests that harnessing the robust angiogenic potential of ECFCs holds promise to use these cells for regenerative purposes. ECFCs are expressing cell surface markers characteristic for endothelial lineage and are devoid of markers such as CD45 or CD14, which suggest that ECFCs are not derived from the hematopoietic stem cell pool. At present, little is known about the origin of these cells as well as the pathways that govern endothelial differentiation of stem or progenitor cells that give rise of ECFCs.
These cells can be isolated from cord or peripheral blood as well as adipose tissue using clonal assays. Previous research reported that ECFCs isolated from cord blood possess different angiogenic and endothelial cell characteristics compared to the cells isolated form peripheral blood. CB-ECFCs proliferate more and are able to form new vasculature that last longer in vivo than PB-ECFCs. 119 Thus, selection of CB-ECFCs seemed to be a more appropriate cell type for regenerative treatments than PB-ECFCs. However, the latter but not the former is more resistant to karyotype abnormalities that can occur during in vitro manipulation. This indicates that ECFCs isolated from peripheral blood are safer and therefore overall a better cell type for regeneration than CB-ECFCs. An alternative was suggested by Prasain et al. 120 who reported successful generation of ECFCs from iPSCs using an induction protocol based on the presence of trophic factors that govern endothelial differentiation. These cells were able to form human vessels in mice and repair the ischemic mouse retina and limb, and most importantly, they lack teratoma formation potential. This approach offers a platform to generate a clinically relevant number of ECFCs in a controlled manner as well as cells from patients with pathologies, which are reported to impair the angiogenic ability of ECFCs,9,121–123 but demands further investigation especially in large animal models.
Animal studies with ECFCs undoubtedly pointed to improved neovascularization and regeneration in different disease models but little is known about the ability of these cells to home to different organs, how the microenvironment clues affect their phenotype and functionality, how ECFCs contribute to tissue-specific cell populations, and to what extent they contribute to tissue repair. After systemic application of ECFCs in rodent models, it was reported that most of the cells were mainly entrapped in the lungs, while only a modest accumulation occurred in the kidneys, liver, and spleen. 124 Intraperitoneal or intrahepatic administration of ECFCs in a fetal sheep model showed that the cells were either located in the perivascular regions of the liver still expressing endothelial makers or migrated to the intestinal crypt region contributing there significantly to the myofibroblast population. 125
Labeling of ECFCs with magnetic particles and guiding the circulating cells with magnetic field into targeted tissue 126 might represent an example of the strategy of how ECFCs can better be accumulated and engrafted into the desired region. This will allow to follow up the phenotype and the functionality of implanted cells over a longer period of time, which will help to better understand the processes that regulate the fate of ECFCs during regeneration in a specific environment and disease.
Conversely to better neovascularization and regeneration, the experience with application of ECFCs in immunocompromized mice bearing small human lung tumors or fibrosarcoma calls for caution since the implanted cells facilitated the disease progression 127 possibly through a paracrine mechanism. 124
Extensive research confirmed that biocompatible scaffolds laden with ECFCs contribute to tissue regeneration by neovascularization as well as by inducing differentiation of implanted or tissue-resident stem or progenitor cells toward desired somatic phenotype. Promising results were achieved in treatment of bone-related 74 or skin pathologies, 79 which warrants translation of this approach into a clinical setting in the near future. However, the plethora of biocompatible materials used for TE studies as well as not yet well understood how additional trophic factors regulate the fate of ECFCs in scaffolds blurs the vision how and when we can use ECFCs for regeneration. For example, treatment of skin defects caused by burns by TE skin equivalents that incorporate ECFCs will demand a different approach than the treatment of skin ulcers caused by diabetic vasculopathy. Therefore, before the use of ECFCs for TE purposes, it is important to clearly underline the scaffold design parameters (cell retention, control cell fate and differentiation, stimulation of tissue formation and remodeling, biodegradability, hemocompatibility, bioactivity in case of natural biomaterials), which should be tailored by the requirements of specific tissue regenerative or pathological milieu.
Accumulated evidence strongly suggests that ECFCs represent a promising and suitable source for prevascularization of tissue-engineered constructs for bone and skin regeneration, endothelialization of tissue-engineered vascular grafts, vascular repair, treatment of myocardial infarction, and other ischemic-induced diseases, as well as a vehicle for gene therapy. However, irrespective of the animal model used to investigate the therapeutic efficacy of ECFCs, it is necessary to clearly define experimental outcomes that can significantly contribute to translation of collected data in clinically relevant situations. In addition, by choosing an appropriate application route that can easily be translated in clinical settings, the regenerative efficacy of ECFCs can be tuned to the demands of the local tissue microenvironment.
Footnotes
Acknowledgments
The project was financially supported by a grant of the Netherlands Initiative for Regenerative Medicine (NIRM). The authors can exclude any conflict of interest and financial relationship related to the content of the presented project.
Disclosure Statement
No competing financial interests exist.