
Abstract
Tissue engineering and regenerative medicine have significant potential to treat bone pathologies by exploiting the capacity for bone progenitors to grow and produce tissue constituents under specific biochemical and physical conditions. However, conventional tissue engineering approaches, which combine stem cells with biomaterial scaffolds, are limited as the constructs often degrade, due to a lack of vascularization, and lack the mechanical integrity to fulfill load bearing functions, and as such are not yet widely used for clinical treatment of large bone defects. Recent studies have proposed that in vitro tissue engineering approaches should strive to simulate in vivo bone developmental processes and, thereby, imitate natural factors governing cell differentiation and matrix production, following the paradigm recently defined as “developmental engineering.” Although developmental engineering strategies have been recently developed that mimic specific aspects of the endochondral ossification bone formation process, these findings are not widely understood. Moreover, a critical comparison of these approaches to standard biomaterial-based bone tissue engineering has not yet been undertaken. For that reason, this article presents noteworthy experimental findings from researchers focusing on developing an endochondral-based developmental engineering strategy for bone tissue regeneration. These studies have established that in vitro approaches, which mimic certain aspects of the endochondral ossification process, namely the formation of the cartilage template and the vascularization of the cartilage template, can promote mineralization and vascularization to a certain extent both in vitro and in vivo. Finally, this article outlines specific experimental challenges that must be overcome to further exploit the biology of endochondral ossification and provide a tissue engineering construct for clinical treatment of large bone/nonunion defects and obviate the need for bone tissue graft.
Introduction
B
Tissue engineering strategies have been developed that exploit the capacity for mesenchymal stem cells (MSCs) to produce bone tissue constituents when grown on biocompatible scaffold materials and in the presence of specific biochemical factors.2,3,20–33 However, biomaterial-based approaches are not yet successful and regenerated bone tissue is not yet widely used for clinical treatment of large bone defects. The first half of this review will summarize conventional bone tissue engineering strategies, focusing in particular on biomaterial-based tissue engineering approaches and their limitations.
Recent studies have proposed that in vitro tissue engineering approaches should strive to simulate in vivo developmental processes and, thereby, imitate natural factors governing cell differentiation and matrix production, following the paradigm recently defined as “developmental engineering.” 34 In particular in vitro approaches, which mimic aspects of bone formation during embryogenesis, can promote mineralization and vascularization to a certain extent both in vitro and in vivo. However, these findings are not widely understood. For that reason, the second half of this review article presents noteworthy experimental findings from researchers focusing on developing an endochondral-based developmental engineering strategy for bone tissue regeneration. The specific in vitro and in vivo methodologies for endochondrally priming MSC constructs are discussed. This review will provide an understanding of the priming conditions needed to produce a mineralized and vascularized construct that when implanted in vivo can induce bone tissue formation.
Bone Tissue Engineering
Tissue engineering and regenerative medicine are promising scientific fields that have significant potential to develop alternative strategies to regenerate deficient bone tissue.2,3,21,35 These approaches strive to regenerate bone in the laboratory environment (in vitro) by exploiting the capacity for biological cells (progenitors) to grow and produce tissue constituents under conditions that emulate the body's biochemical and physical environment. Abundant research has been dedicated to identifying specific factors and the desired properties of biocompatible scaffold materials to encourage bone regeneration in vitro.2,3,20–33 In particular various biomaterial scaffolds have been explored as conduits for bone regeneration, with the aim of identifying the most appropriate chemical, biological, and physical properties to encourage in vitro bone production.24–30
Naturally derived polymers, such as collagen, glycosaminoglycans (GAGs), gelatin, chitosan, silk, fibrin, and elastin, have been widely used in a variety of tissue engineering applications.36,37 Collagen-based scaffolds in particular are widely used for bone tissue engineering applications,38,39 and when combined with GAGs enhance osteoblastic differentiation both in vivo and in vitro.40–45 Synthetic polymers, such as poly(lactic acid) (PLA) films and poly(lactic-co-glycolic acid) (PLGA), can induce osteoblastic activity that is indicative of bone formation,28–30 and their mechanical properties, shape, microstructure, and degradation rates can be tailored to suit various applications.46,47 Ceramic hydroxyapatite (HA) and β-tricalcium phosphate (β-TCP) scaffolds are also osteoconductive48–51 and can be combined with osteogenic growth factors and bioactive proteins.52–54 Vascular networks have been integrated into HA ceramic scaffolds to increase bone formation. 50 Bioactive glasses, such as calcium containing silica glasses, can produce a bioactive hydroxycarbonated apatite layer to enhance bonding to biological tissue 52 and their resorption rates can be tailored to deliver ions in a controllable manner to activate cell differentiation.52,55
There are many limitations to biomaterial scaffolds. Synthetic polymers can release acidic by-products and stimulate adverse tissue and inflammatory reactions. 47 Ceramic scaffolds and bioactive glasses are extremely brittle, and thus, unsuitable for treatment of fractures/defects in skeletal locations where physical loading is expected. Composite scaffolds of both ceramic and polymer, such as Collagen-HA (CHA) and polycaprolactone-calcium phosphate (mPCL-CaP) scaffolds, are being explored as alternatives to overcome the limitations of low mechanical stiffness seen in polymer-based scaffolds and the brittle nature of calcium/bioactive glasses.29,37,56,57 However, MSC-seeded scaffolds show variable success with regard to formation of a strong mineralized bone matrix, 32 and once implanted MSC-seeded scaffolds become encapsulated31,58,59 and host vasculature is inhibited, which can lead to cell death (necrosis), and ultimately, failure of the implant construct.31,58–61 Clinical trials are investigating the efficacy of bone tissue engineering constructs using autologous MSCs (Table 1) and in general, the results from these studies demonstrated that it is safe to use human MSCs for bone regeneration. However, the bone tissue regeneration potential of these studies thus far has been substantially less promising than those of animal studies.62,63 Therefore, biomaterial-based approaches are not yet successful and regenerated bone tissue is not yet widely used for clinical treatment of bone defects.
CT, computed tomography.
Developmental Engineering
Bone tissue engineering strategies largely strive to produce constructs that either mimic the functional and mechanical properties of the bone tissue32,64–78 or provide a conduit to deliver biochemical and biological factors to induce bone formation.2,24,31,40,41,71,79–103 In contrast the premise of developmental engineering approaches is not to replicate the properties of the fully functional tissue, but rather to simulate in vivo developmental processes and, thereby, take advantage of the natural mechanisms governing cell differentiation and matrix production by which ossification occurs in the body. Bone development during early fetal development arises by two distinct biological processes: intramembranous ossification and endochondral ossification. Both processes begin when an organic matrix (osteoid/cartilage template) is initially laid down by osteoblasts and then mineral crystals are produced and grow slowly over time to produce bone tissue. 104 Recent tissue engineering strategies have sought to replicate certain aspects of the endochondral ossification with promising results,33,105–114 and thus, this review will focus hereafter on the biological process of endochondral ossification and developmental engineering based on replicating this in vivo process.
Endochondral Ossification
Long bones within the body are created through a process known as endochondral ossification during early fetal development. This process begins when MSCs condense into clusters and begin to differentiate to become chondroblast cells, which begin to secrete a matrix rich in collagen type II and the proteoglycan aggrecan, to form a rudimentary cartilage template and also form a membrane around this, known as the perichondrium. The chondroblasts differentiate to become chondrocytes, and the cartilage aggregate grows through chondrocyte proliferation and matrix production.104,115,116 Bone tissue formation then proceeds through a series of critical events: (1) Chondrocytes cease to proliferate within the cartilage template, enlarge, and synthesize type X collagen, (2) these chondrocytes become hypertrophic, (3) the hypertrophic chondrocytes signal for the influx of blood vessels by producing vascular endothelial growth factor (VEGF) and osteoblasts, 117 and finally (4) hypertrophic chondrocytes located directly adjacent to the perichondrium differentiate to become osteoblasts and form an extracellular matrix containing Alkaline Phosphatase (ALP) 115 at sites of initial mineralization118,119 and secrete collagen type I-rich matrix resulting in the formation of the bone collar (Figure 1). After this initial bone formation, further bone production and growth occur at the secondary ossification centers and are regulated by the epiphyseal or growth plate to facilitate lengthening of bone. After puberty, the lengthening of bones stops and the growth plate fuses and is replaced by bone, known as the epiphyseal line. 104

Endochondral ossification: (1) formation of the cartilage template, (2) chondrocytes become hypertrophic and endothelial and MSCs begin to invade and vessels are formed, (3) mineral begins to be deposited upon the cartilage template, (4) marrow, secondary ossification center and epiphyseal plate are formed. MSCs, mesenchymal stem cells.
Endochondral Ossification Based Developmental Engineering Strategies for Bone Tissue Regeneration
As outlined above, endochondral ossification is a tightly regulated process that is governed by the multicellular population, which produces various biochemical signals to direct chondrogenesis, vascularization, and ultimately, bone matrix production. Recently, researchers have sought to investigate whether the recreation of such cues in vitro could manipulate the cells to undergo an endochondral-like ossification process both in vitro and following implantation in vivo. These specific research studies are discussed in detail below.
Formation of the cartilage template
The cartilage template is a fundamental precursor for bone development of long bones during endochondral ossification. Thus, researchers have investigated whether the exposure of MSCs to the biomechanical cues of cartilage formation in vitro might ultimately manipulate the cells to undergo endochondral ossification. The premise of such studies is that, by enabling the cells to form the cartilage template before mineralization, these approaches mimic more closely the endochondral ossification process that occurs naturally during early fetal development. Chondrogenic differentiation can be induced when MSCs are exposed to chondrogenic differentiation supplements, including TGF-β3, dexamethasone, ascorbic acid,
In vivo implantation of chondrogenically primed MSCs, in both aggregate and scaffold form, has been conducted to investigate the potential for healing large segmental defects.111,129 Chondrogenic priming of rat MSCs seeded on a PLGA scaffold was conducted and these constructs were implanted into both critical sized (5 mm) and full thickness (15 mm) rat femoral defects. 111 Newly formed cortical bone was reported for all of the constructs examined radiographically by 8 weeks, and full bone union was reported 16 weeks postsurgery. Chondrogenically primed human MSC aggregates were also implanted into a 6 mm femoral defect. 129 Micro-computed tomography (CT) scanning revealed that there was significantly more bone formation in the defect treated with chondrogenically primed MSCs compared to undifferentiated MSCs after 4 and 8 weeks. These studies have established that chondrogenic priming of MSCs can entice bone formation in both ectopic and critical defect models. However, some of these studies observed core degradation 108 and an uneven distribution of bone mineral throughout the construct.33,107
It is likely that the duration of cartilage template formation is important for bone formation by endochondral ossification. Cartilaginous limb buds have been estimated to form in humans by the 26th day of gestation,120–122 whereas in vivo studies have shown that bone formation begins in chick bones when the embryo is 9–18 days old.125–129 A recent study sought to determine the optimum time for chondrogenic priming of MSCs to enhance osteogenic differentiation by MSCs in vitro. 130 It was shown that chondrogenic priming for specific durations (14, 21 days), before being exposed to osteogenic supplements, can enhance both the osteogenic and chondrogenic differentiation of both murine MSCs and human MSCs in vitro. 130 Chondrogenic priming also induced more in vitro mineralization by MSCs compared to culture with osteogenic supplements alone. Furthermore, it was demonstrated that chondrogenic priming for 21 days enhanced the distribution of minerals and resulted in a construct with mineralization throughout, rather than around the periphery. 130 Other studies have shown that the duration of media supplementation with chondrogenic factors is also important for the mineralization of bone tissue constructs in vivo.107,128 Human MSCs were chondrogenically primed on transwell inserts for either 1 or 2 weeks before subcutaneous implantation in a mouse model. The constructs that were chondrogenically primed for 1 week were absent 4 weeks post implantation, whereas constructs primed for 2 weeks developed an extracellular matrix rich in GAG and contained cells embedded in large lacunae. By 8 weeks the center region of these constructs was rich in collagen type X, indicating hypertrophy. 107 The in vivo potential of chondrogenically primed rat MSCs grown on polycaprolactone (PCL) or HA/TCP composite scaffolds has been investigated for a variety of priming periods (2, 3, and 4 weeks). 128 The results from this study showed that constructs primed for 3–4 weeks had a homogeneous deposition of cartilage matrix on both PCL and HA/TCP composite scaffolds in vitro, whereas in those primed for 2 weeks the cartilage matrix was mainly located on the periphery of the constructs. However, after 8 weeks of subcutaneous implantation, all implanted constructs gave rise to endochondral bone formation, and the bone volume was not affected by the length of priming time (2, 3, and 4 weeks). 128
Collectively these studies demonstrate that allowing cells to form a cartilage template, through chondrogenic priming, has a beneficial influence on both the in vitro and in vivo mineralization potential of MSCs and bone tissue engineering constructs. Moreover, the amount of time the cells are chondrogenically primed for can influence both the osteogenic and chondrogenic differentiation potential of MSCs both in vitro and in vivo. However, full bone bridging was only reported after 16 weeks, whereas other bone tissue engineering strategies have reported full bridging after 12 weeks.131–134 Moreover, none of these constructs had the mechanical integrity to withstand the necessary loads exerted due to everyday loading. Therefore, it is evident from these studies that mimicking the cartilage template alone may be insufficient to heal a large bone defect.
Hypertrophic cartilage template formation
After the cartilage template is established, chondrocytes within the cartilage template begin to undergo hypertrophy and recruit a mixture of cells (MSCs and endothelial cells) that are responsible for ossification and vascularization of the cartilage template. Hypertrophy can be induced in vitro under specific conditions, namely by culturing in thyroxine, β-glycerophosphate, dexamethasone, ascorbic acid, and IL-1β or by allowing chondrogenic differentiation to occur for a significant period of time (over 28 days). Hypertrophic priming is the process in which cells are either allowed to become hypertrophic, through long term culture, or are exposed to hypertrophic growth factors, as mentioned above. In vitro studies have investigated whether inducing hypertrophy in vitro would enhance the mineralization and vascularization potential of bone tissue engineered constructs. Porcine MSCs seeded on agarose hydrogels were cultured in hypertrophic medium (chondrogenic medium supplemented with β-glycerophosphate), and these conditions were shown to induce mineralization in vitro as seen through Alizarin Red Staining. 135 However, human MSCs cocultured with endothelial cells in normoxic conditions (oxygen levels of 20%), allowed for the formation and maintenance of prevascular structures, whereas the same constructs cultured in hypoxic conditions (oxygen levels of 5%) led to hypertrophy of the aggregates and did not allow for the formation of preliminary vascular structures. 136
Other studies have investigated whether hypertrophic cartilage grafts would induce bone formation in ectopic locations in rodents.109,112,137–139 Human MSC aggregates,
137
human MSC-seeded decellularized cartilage matrices,
112
and porcine MSC-seeded agarose hydrogels
138
were cultured in either chondrogenic medium112,137 or hypertrophic medium
138
for long durations (6–7 weeks) so that hypertrophy ensued. These hypertrophic cartilage grafts were found to undergo alterations similar to the endochondral ossification process, namely calcification, vascular invasion, and ultimately the deposition of minute ossicles, rather than differentiating further down the chondrogenic pathway.112,137,138 Human MSCs seeded on a collagen scaffold were exposed in vitro to hypertrophic factors before subcutaneous implantation and were found to induce bone tissue formation complete with an outer cortical-like perichondrial bone and an inner trabecular-like endochondral bone after 12 weeks.
109
Moreover, there was an ingrowth of blood vessels, and marrow cavities were beginning to form within the ossicles.
109
Other studies have shown that subcutaneous implantation of a tissue engineered decellularized hypertrophic cartilage could induce vascularization and de novo mineral accumulation.
139
Moreover, when these were implanted in a critically sized segmental defect, full bridging was observed in over half of the defects, whereas no bridging was found in the empty control. One study compared the bone regeneration potential of an endochondrally primed CHA construct to a CHA cultured in osteogenic medium alone.
140
Similar to previous studies, to induce hypertrophy collagen-hyaluronic acid (CHyA) and CHA, constructs seeded with rat MSCs were chondrogenically primed for 21 days with a further 14 days in hypertrophy media (containing
Prevascularization of bone tissue engineered constructs
Vessel formation is a critical and integral part of the bone formation process that occurs during endochondral ossification in vivo. Vessel invasion typically occurs between 14 and 18 days of embryogenesis when the cartilage template is established141,142 and involves endothelial cells invading through the cartilage canals already present in the developing bone tissue115,116,141,143 by a process known as quiescent angiogenesis. It is believed that bone vasculature provides the necessary growth factors, hormones, chemokines, and nutrients to induce bone mineralization. Therefore, it has been proposed that cells within tissue engineered constructs that have no vascular supply do not get the necessary factors to regenerate the damaged bone tissue and, ultimately, die when implanted in vivo.144,145
Certain bone tissue engineering strategies have strived to prevascularize bone tissue engineered constructs by different mechanisms as follows: (1) growth factor delivery, (2) microsurgery, (3) scaffold fabrication, and (4) coculture methodologies, whereby vascular cells are cultured with the MSCs. Due to the tight link between osteogenesis and angiogenesis, delivery of angiogenic growth factors to the site of injury has shown great promise for bone healing by significantly increasing the vascularization of the constructs once implanted in vivo.146,147 However, the uncertainty with dosage, the instability of the growth factors in vivo, and the shear cost of therapy have meant that further investigation is required. 148 Microsurgery involves taking advantage of the hosts own vasculature network and wrapping it around the engineered constructs using techniques such as flap fabrication or arteriovenous loop. Despite the increase in bone formation,149–151 this strategy is highly invasive and involves two different surgical interventions, and there is a much greater chance of donor site morbidity. The development of scaffolds for bone tissue engineering, which support not only the bone forming cells but also house a vascular network, is a new promising technique.152–154 The results from such studies have been contradictory, owing to the fact that scaffolds with smaller pores to support angiogenesis induced chondrogenesis rather than osteogenesis through the endochondral pathway.152,154 These small pores favored hypoxic conditions and induced osteochondral formation before osteogenesis, whereas the larger pores allowed for vascularization and led to direct osteogenesis.152,154 Thus there is a clear need for further studies to fully develop a scaffold that will induce both bone formation and vasculogenesis.
During endochondral ossification complex biochemical signaling is governed by multiple cell populations to induce vascularization. When hypertrophy of the cartilage template begins, hypertrophic chondrocytes directly adjacent to perichondrial cells differentiate to become osteoblasts and secrete growth factors to promote invasion of MSCs and endothelial cells to induce vessel formation and infiltration.155,156 In vitro, coculture studies have investigated whether coculture approaches, involving MSCs and endothelial cells, might mimic this in vivo multicellular niche. In particular, direct cell–cell contact of MSCs or primary osteoblasts and endothelial cells has been induced through the formation of cellular aggregates,136,157–161 and such studies have reported that prevascular networks can be formed in cellular aggregates cocultured in vitro.136,157,158,160,161 Other studies have investigated whether prevascularizing three-dimensional (3D) tissue engineered scaffolds, such as trabecular bone, 162 PCL, 163 poly(LLA-co-DXO), 164 collagen GAG,165,166 and HA 167 scaffolds in vitro, through the coculture of MSCs and Human Umbilical Vein Endothelial Cells (HUVECs), would allow faster host integration postimplantation.162–167 It has been shown that prevascular networks can be formed in a subcutaneous animal model in vivo when human MSCs162,164,165,167 are first cocultured with HUVECs in vitro. In vitro studies have shown that direct coculture of MSCs or osteoblasts with endothelial cells can upregulate production of the early osteogenic marker ALP,168–171 without the presence of osteogenic supplements. Other studies have investigated whether coculture of MSCs and endothelial cells can increase ALP production in 3D polymer scaffolds164,172 or 3D and cellular aggregates.157–159,173 The coculture of MSCs or osteoblasts with chondrocytes has been shown to have beneficial effects on ALP production in both two-dimensional (2D) and 3D in vitro culture.174–176 One study investigated the effect of coculture of human MSCs and chondrocytes without the use of osteogenic supplements and found there was no ALP production/expression in 3D aggregate culture. 175 However, a study involving direct 2D coculture of rat osteoblasts and bovine chondrocytes reported that there was significantly higher ALP activity in the coculture group compared to the osteoblast group alone. 176 Coculture of MSCs and endothelial cells through transwell inserts has been shown to induce MSCs to undergo both osteogenesis and chondrogenesis through the endothelin-1 phosphatidylinositol 3-kinase/AKT (AKT) signaling pathway. 177 It is clear from such studies that the signaling that occurs between the different cell types present within the cartilage template is imperative for the formation of bone in vivo.
Combined formation of the cartilage template and prevascularization of the cartilage template
A recent study sought to incorporate the production of the cartilage template and the vascularization of the construct simultaneously motivated by the fact that both chondrogenesis and vascularization are crucial precursors for bone formation during endochondral ossification in vivo. It was shown that chondrogenic priming of human MSC aggregates (for 21 days), followed by coculture of human MSCs and HUVECs to entice vascularization, significantly increased the osteogenic potential of the culture compared to chondrogenic priming alone. 161 This study also reported that both MSCs and HUVECs must be added to the formed cartilage template for the formation of rudimentary vessels to occur in vitro. 161 Moreover, it was shown that both chondrogenic priming (for 21 days) and subsequent vascular priming (coculture with endothelial stem cells and MSCs) can induce osteogenesis of a 3D scaffold-less construct without the use of any osteogenic supplements. In fact, the results indicated that the addition of osteogenic supplements inhibits early osteogenic markers (ALP expression), promotes late osteogenic markers (calcium content), and produces mineral deposition throughout the cellular aggregate, rather than in the form of discrete mineralization nodules. 178 An in vivo study, with a subcutaneous immunocompromised rodent model, demonstrated that aggregates that had been both chondrogenically primed and prevascularized before implantation were found to have mature endogenous vessels (indicated by α-smooth muscle actin walls and erythrocytes) after 4 weeks of subcutaneous implantation and also viable human MSCs (detected by bioluminescent imaging) 21 days after subcutaneous implantation. In contrast, aggregates that were not endochondrally primed had no vessels present within the aggregate interior and human MSCs did not remain viable beyond 14 days. Interestingly, the endochondrally primed aggregates were also the only group to have mineralized nodules present within the cellular aggregates. 179 Taken together, these studies suggest the beneficial role that mimicking two or more critical aspects of the endochondral ossification process can have on bone tissue formation.
Conclusions and Future Perspective
To date, bone tissue engineering strategies have either designed scaffolds that mimic the mechanical properties of natural bone tissue32,64–78 or have designed scaffolds that incorporate the cells or necessary growth factors to induce osteogenesis and then degrade.2,24,31,40,41,71,79–103 However, recently, researchers have sought to design constructs that replicate separately aspects of the bone formation process that arise during bone development, specifically (1) the formation of the cartilage template during endochondral ossification,33,107–109,111–113,129 (2) the formation of a hypertrophic cartilage template,109,112,135–137,180 or (3) the prevascularization of the construct.157,158,162–167 Each of these studies has advanced the understanding of in vitro and in vivo factors that can govern bone regeneration (Figure 2).
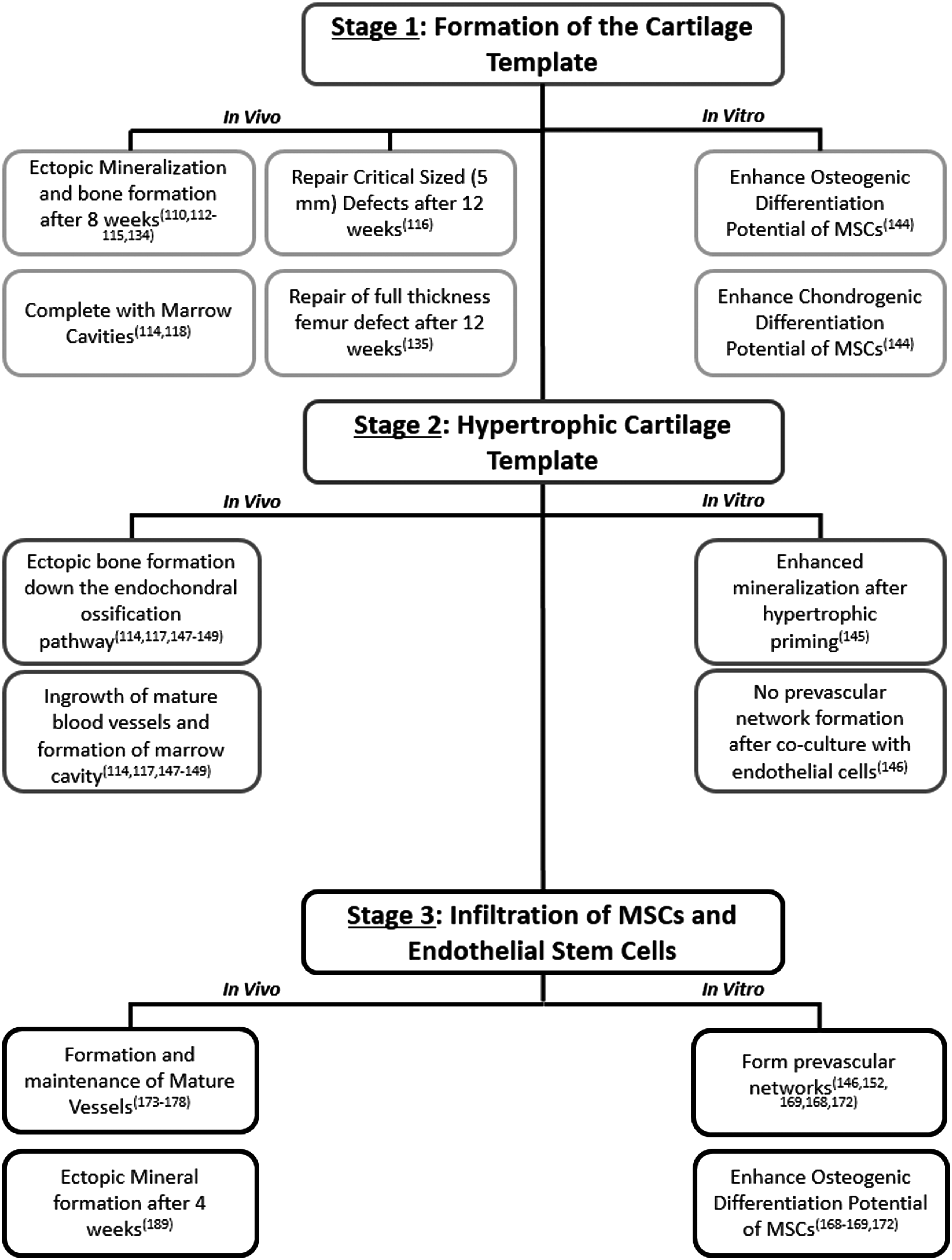
Flowchart of the in vitro and in vivo results of constructs, which have been designed to replicate specific aspects of the endochondral ossification process.
The provision of bone tissue constructs appropriate for clinical use is limited, due to issues with core degradation and an uneven distribution of bone mineral throughout the construct.33,107,108 In particular no study has managed to design an in vitro bone tissue engineering construct that can ensure osteogenesis, angiogenesis, and survivability once implanted in vivo. The individual studies reported in this review provided a novel understanding of potential endochondral priming conditions that might serve to eradicate previous limitations. When analyzed together, the findings of this review provide a novel perspective on the optimum conditions needed to regenerate bone tissue for treatment of large bone defects.
However, several challenges still remain to be addressed before such bone tissue engineered constructs can be used for treatment of a large bone defect in a load bearing location and these are outlined as follows.
(1) While coculture of MSCs, endothelial cells, and chondrocytes can regulate both the mineralization and vascularization potential of MSCs in vitro, the exact signaling pathways that are activated need to be investigated to fully understand the role of these cells during the endochondral ossification process in vivo and, thereby, strive to determine the optimum in vitro/ex vivo culture methodologies.
(2) It has been shown previously that mimicking the combination of the formation of the cartilage template and the prevascularization of this template improved survivability, enhanced vessel formation, and increased mineralization within the cellular aggregate when implanted subcutaneously in vivo. 179 However, currently no approach has sought to incorporate all three critical aspects of the endochondral ossification process. Therefore, future bone tissue engineering strategies should investigate a three-step endochondral priming process combining (a) chondrogenic priming, (b) hypertrophic priming, and (c) vascular priming.
(3) Experimental studies have found that the application of mechanical stimulation can influence the rate of endochondral ossification.181–183 Specifically, the application of hydrostatic pressure can significantly enhance the chondrogenic differentiation process of stem cells and, thus, influence the rate of endochondral ossification.184–192 However, in vivo, all of these three aspects, chondrogenesis, vascularization, and mechanical stimulation of the cartilage template, are essential for bone formation. To date no approach has sought to recreate both the biochemical and mechanical extracellular environment of the endochondral ossification process to enhance the mineralization potential of bone tissue engineering constructs. Future studies should investigate an in vitro tissue regeneration strategy that not only mimics the cellular and biochemical environment but also the mechanical environment applied during endochondral ossification in vivo. Mathematical and computational models have predicted that a variety of stimuli can both promote and inhibit cells to undergo chondrogenic and osteogenic differentiation during bone development193–196 or fracture healing.197–200 However, the appropriate level of hydrostatic pressure and other physical parameters that will drive bone regeneration by endochondral ossification are still unknown.
(4) Finally, the exact degree to which endochondral priming should be induced in vitro, before implantation in vivo, is largely unknown. In particular, it is debated whether a construct could be allowed to become fully mineralized in vitro such that it could withstand the implantation in a load bearing location. Moreover if this fully mineralized construct would allow for faster host integration and bone formation. A concern with such an approach is that the mineralized construct would be resorbed when implanted in vivo and hinder integration within the defect, however, this requires further investigation. However by striving to overcome these hurdles, a bone tissue engineering construct might be created in vitro that when implanted can autonomously engraft, remodel, and, ultimately, lead to enhanced bone healing for clinical treatment of large bone/nonunion defects and obviate the need for bone tissue grafts.
Footnotes
Disclosure Statement
No competing financial interests exist.