
Abstract
Collagen-based biomaterials are widely used in the field of tissue engineering; they can be loaded with biomolecules such as growth factors (GFs) to modulate the biological response of the host and thus improve its potential for regeneration. Recombinant chimeric GFs fused to a collagen-binding domain (CBD) have been reported to improve their bioavailability and the host response, especially when combined with an appropriate collagen-based biomaterial. This review first provides an extensive description of the various CBDs that have been fused to proteins, with a focus on the need for accurate characterization of their interaction with collagen. The second part of the review highlights the benefits of various CBD/GF fusion proteins that have been designed for wound healing and bone regeneration.
Introduction
C
Most of the collagen-derived biomaterials that have been developed for tissue engineering and regenerative medicine purposes simply trap the drugs in their network. 6 Drug release is thus governed by diffusion through the pores of the scaffold, which makes sustained delivery over a long period difficult to achieve. In particular, there is generally an initial burst release, which causes high drug loss and possible side effects.7,8 For example, a burst of bone morphogenetic protein-2 (BMP2) was shown to promote inflammation and osteoclastic activity. 7 Although extended release may be attained by reducing the size of the pores, this strategy hinders nutrient transport and severely affects cell viability. 9
GFs are frequently used in therapeutic strategies such as bone and cartilage engineering, 10 myocardial regeneration, 11 and wound healing 12 because they can promote cell adhesion, migration, differentiation, and proliferation. Their efficacy, however, depends highly on their spatiotemporal delivery, an essential trait that can affect cell fate. 13 Although no GF binding site has been reported in collagen yet, 14 some GFs, for example, transforming growth factor-β1 (TGF-β1), basic fibroblast growth factor (bFGF), and BMP2, naturally present a strong affinity for collagen and can bind to collagen-based scaffolds via ionic interactions.7,15,16 Their capture, however, depends on the isoelectric point of the GF 7,17 and the release profile usually features an initial burst, 7 which makes this approach unsuitable for controlled release.
A variety of techniques have been developed for the covalent chemical conjugation of GFs to collagen scaffolds so as to provide a highly localized and long-lasting signaling. The random covalent grafting of GFs has proved worthwhile,18,19 in particular by photoirradiation 20 or through the NHS/EDC coupling chemistry that targets free amine groups on the GF. 21 The attachment of the proteins to the substrate may however mask or alter their receptor binding moieties and, consequently, negatively impact their biological activity.22,23 The covalent binding of GFs in an oriented manner with the help of a cysteine tag has also been investigated. 24 The addition of a cysteine may, however, hamper protein folding via the formation of unwanted disulfide bridges, hence also resulting in a loss of activity. Importantly, covalent binding may prevent the endocytosis of the GF, which can be crucial for signaling. 25
To circumvent these limitations, numerous research groups have focused on the tethering of GFs in a stable but noncovalent manner using specific affinity pairs. Strategies that have been reported involve biotin and streptavidin, 26 two complementary coil peptides,27–29 DOPA-containing peptides, 30 or specific binding domains that target heparin/heparan sulfate, 31 laminin, 32 fibrin, 33 or fibronectin (FN). 34 In particular, fusion proteins with a collagen-binding domain (CBD) have been the subject of intense research to functionalize collagen-based biomaterials. Interestingly, type I collagen is the most abundant protein in the human body 35 and is found in the extracellular matrix (ECM) of bones, skin, tendons, cornea, artery walls, as well as internal organs. Hence, a recombinant collagen-binding GF tethered to a resorbable collagen biomaterial would also bear the potential to bind to the extracellular collagenous matrix after resorption of the implant, thus exhibiting a significantly prolonged bioavailability.
The purpose of this review is (1) to examine the various CBDs that have been characterized and discuss how their origin and size affect their affinity and specificity for collagen and (2) to assess to which extent collagen biomaterials functionalized with CBD fusion proteins have emerged as powerful tools in the fields of wound healing and bone regeneration.
Collagen-Binding Domains
The term CBD encompasses very distinct polypeptide domains, which are either engineered or derived from native collagen-binding proteins such as FN, the von Willebrand factor (vWF), and several collagenases. This diversity of origin results in a great heterogeneity of sizes, from a seven amino acid-long peptide to a 42-kDa protein domain (Table 1). Bearing in mind that these domains will be fused to GFs and that controlling their release is of paramount importance for tissue engineering and regenerative medicine applications, it is crucial to assess how their sequence and their length affect their interaction with collagen, especially in terms of stability.
See Quantitative Characterization of CBD-Collagen Interactions section for methodological approach.
The plateau (saturation) has not been reached during the experiment.
bFGF, basic fibroblast growth factor; BMP, bone morphogenetic protein; CBD, collagen-binding domain; ELISA, enzyme-linked immunosorbent assay; FN, fibronectin; LRR, leucine-rich repeat; PDGF, platelet-derived growth factor; PlGF, placental growth factor; SPR, surface plasmon resonance; VEGF, vascular endothelial growth factor; vWF, von Willebrand factor.
Quantitative characterization of CBD-collagen interactions
For most of the CBDs under study, either an apparent affinity (thermodynamic association constant, KA, expressed in M−1), or its inverse, the thermodynamic dissociation constant (KD, expressed in M), has been reported to characterize the strength of their interaction with collagen. Given the diversity of their origin and of the types of collagen, it is unsurprising that the dissociation constant values broadly span from the low nanomolar to the micromolar (Table 1). Such a range of affinities for CBD-collagen interactions could be exploited to target certain types of collagen with high specificity and/or to finely tune the release rate of a protein of interest, by fusing it to an appropriate CBD. However, to avoid any misinterpretation of the data presented in Table 1, one may analyze them with caution.
On the one hand, many techniques have indeed been used by the research community to determine the affinities of the various CBDs for the different types of collagen. Those include surface plasmon resonance (SPR)-based biosensor assays, 36 enzyme-linked immunosorbent assays (ELISA), 14 radioactivity assays by using iodinated proteins, 37 as well as fluorescence titration assays by measuring changes in anisotropy of fluorescein-labeled collagen chains. 38 All of these methods have their own merits and limitations to assess an apparent dissociation constant. 39 Interestingly, large differences can be pinpointed between the values derived from distinct techniques within the same report. For example, when characterizing the interactions between the von Willebrand A1 domain for type III collagen, Morales et al. determined an apparent KD of 8 nM by SPR and of 400 nM by ELISA. 40 Such a discrepancy can result from several biases inherent to the techniques at hand. For instance, in most SPR assays, the collagen is covalently attached to the biosensor surface, whereas in ELISA, the substrate is nonspecifically adsorbed at the bottom of the well. Both approaches may thus lead to distinct alterations of the three-dimensional conformation of the triple helix of collagen and, as a result, affect its interaction with CBDs. 40 Other artifacts related to inappropriate assay conditions, as those documented in specialized reports,41,42 may also be invoked to explain these discrepancies.
On the other hand, it is not uncommon that distinct research groups report different apparent KD for the same interaction, even when assessed with the same assay (Table 1). For example, Sun et al. reported an apparent KD of 5.5 nM for the capture of the fusion protein TKKTLRT-PDGF (platelet-derived growth factor) by a type I collagen membrane, whereas Lin et al. reported an apparent KD of 97 nM for the same protein incubated on type I collagen-coated wells.43,44 As much as the techniques, the procedure for data processing and KD determination may therefore be worth questioning. In most of the articles here reviewed, the apparent KD values were indeed derived by linearizing the experimental data via a Scatchard plot for instance. Such an approach is prone to introduce errors and biases, 45 especially when the plot deviates from linearity and when data are omitted.46,47
To overcome these limitations, we here propose a rationalized method for data processing. By curve fitting the complete data set with the GraphPad Prism 6 Software (GraphPad Prism Software, Inc., San Diego, CA), using a Hill-type equation that assumes a 1:1 interaction, the half-maximal effective concentration (EC50) of a CBD binding to collagen can be calculated:
where [CBD] and Y correspond to the incubated CBD concentration and the corresponding response (signal minus blank), respectively. Ymax, the theoretical maximal response, and EC50, the half-maximal effective concentration, are outputs. Note that the Hill slope was fixed at 1.0 for the sake of comparison.
This curve-fitting method was applied to all the CBD-collagen interaction studies when raw results were available, by extracting the data from the research article figures using the WebPlotDigitizer 3.9 software. 48 When applicable, the EC50 values derived from the fits, the coefficient of determination (R2), as well as the number of experimental data points extracted from the figures (n) are given in Table 1, in addition to the apparent dissociation constant value (KD) that was reported by the authors. Although all R2 values were close to 1, which is indicative of a good fit between the model and the data, some research groups have used a low number of CBD concentrations in their investigation. Given that the reliability of any curve fitting is highly dependent on the number of data points, both the KD values they reported and the EC50 values we calculated need to be treated with caution. Furthermore, several cases of biphasic interactions were encountered, for which we only provided the lowest EC50 (corresponding to the highest apparent affinity).
With this analytical approach, we found that the interaction of TKKTLRT-PDGF for type I collagen, with reported KD values of 5.5 and 97 nM by Sun et al. and Lin et al., respectively, was best depicted by an EC50 of 1100 nM in both cases.43,44 This calculated value is plausible, given that the majority of the TKKTLRT fusion proteins feature a dissociation constant in the low micromolar range (Table 1).
Fibronectin
FN is a 440-kDa dimeric protein of the ECM. It binds to many biological partners such as collagen, fibrin, heparin, and integrin (Fig. 1A). FN has a very high apparent affinity for type I and type II collagens (KD values of 13 and 58 nM, respectively).49,50 FN binds even more strongly to denatured collagen, that is, gelatin (KD = 2 nM),50,51 which suggests that its binding sites within collagen are, at least partially, masked in the native triple helix.49,52

Structure of several proteins containing a CBD: human FN
The CBD of FN is a 42-kDa domain composed of four type I and two type II modules (I6-II1-II2-I7-I8-I9, see Fig. 1A). All six modules are required for full affinity, 53 supporting a cooperative mode of interaction where they simultaneously participate.54,55 The FN type II modules are found in many other collagen-binding proteins, including the mannose receptor, 56 the factor XII, 57 and the matrix metalloproteinase 2 (MMP2). 55 Similarly to the full-length protein, the CBD of FN displays a higher affinity for gelatin than for properly folded collagens. It is, however, monomeric and has a moderate affinity for type I collagen (KD = 420–800 nM),49,58 hence highlighting the importance of FN dimerization (and of the resulting avidity) for optimal binding to collagen. Numerous subdomains containing three or more modules have been studied, including a short peptide sequence extracted from the module I9, CQDSETRTFY, which binding to type I collagen proved to be very weak (EC50 = 900,000 nM, Table 1).59,60
von Willebrand factor
The vWF is a multimeric blood glycoprotein involved in hemostasis and formed from 275-kDa subunits crosslinked via disulfide bonds. 61 The mature vWF has a very high affinity for type I, type III, and type VI collagens (KD values of 1.8, 3.4, and 8 nM, respectively)62–64 but poorly binds to gelatin, in stark contrast to FN.65,66 Several moieties of the vWF came under scrutiny for their ability to bind to collagen (Fig. 1B), in particular, the 20.3-kDa A1 domain and the 19.3-kDa A3 domain. 67 Although conflicting results exist in the literature, the A1 domain of vWF does not seem to play a significant role in the binding of vWF to type I and type III collagens68,69 but is crucial for its interaction with the type IV and type VI collagens.64,70,71 In contrast, the A3 domain is considered to be the main collagen-binding site of vWF72,73 and has but a moderate affinity for the type I and type III collagens (KD values of 1800 and 1600 nM, respectively).40,67 The A3 domain is thought to interact with the RGQAGVMGF/RGEOGNIGF sequences of the α1/α2 chains of type I collagen and with the RGQOGVMGF sequence in type II and III collagens. 69
Shorter domains of vWF have also been studied for their ability to bind to collagens, in particular, WREPSFCALS and WREPGRCELN that were derived from the bovine and human vWF, respectively (Fig. 1B). 37 To avoid unwanted homodimerization or misfolding,74,75 two variants bearing a cysteine-to-methionine substitution in position 7, that is, WREPSFMALS and WREPGRMELN,76,77 have also been studied. The WREPSFCALS peptide is derived from a 21-kDa sequence (F570-K682) of the bovine vWF precursor (NP_001192237.1); this sequence, first described by Takagi et al., is located at the end of the propeptide 78 that is cleaved during posttranslational modifications. 79 As opposed to the A3 domain, the affinity of the WREPSFCALS peptide for type I collagen is very weak (EC50 = 29,000 nM, Table 1). 37 Nonetheless, it binds to all collagen types from I to V, in addition to gelatin. 37 Its 21-kDa parent fragment possesses the same characteristics but displays a higher affinity for type I collagen (EC50 = 780 nM, Table 1). 37
Placental growth factor
The placental growth factor (PlGF), a member of the vascular endothelial growth factor (VEGF) family, is a glycoprotein essential for angiogenesis, in particular, over the bone marrow-derived cells. 80 The PlGF gene undergoes alternative splicing and gives rise to several homodimeric variants between 33.4 and 49.4 kDa. The sequence analysis of two splice variants that interact differently with the ECM, namely PlGF1 and PlGF2, enabled the identification of a 2.8-kDa amino acid sequence that binds to collagen (RRRPKGRGKRRREKQRPTDCHL, Fig. 1C). 14 While initially considered as a heparin-binding domain, 81 this sequence binds very strongly to type I collagen (KD = 126 nM) and even more to FN, vitronectin, heparan sulfate, and other ECM proteins. 14 Although it has a very high isoelectric point (pI = 12.0), a scrambled version of the peptide did not bind to type I collagen, thus indicating that the interaction of this CBD with collagen is sequence dependent and not merely electrostatic. 14 Conversely, a C-terminal cysteine-to-serine substitution had little to no influence on the interaction. 14 To our knowledge, no reports have yet been made regarding the specificity of this CBD toward other collagen types.
Clostridium histolyticum collagenases
ColH (116 kDa) and ColG (126 kDa) are two collagenases from the pathogenic C. histolyticum (Fig. 1D). 82 They feature a broad substrate specificity and target various types of collagen as well as gelatin.83,84 For instance, the full-length ColH enzyme displays a strong affinity for type I collagen (KD = 99.5 nM). 85 Their CBDs, corresponding to the S3 domain (13 kDa) for ColH and the S3a+S3b domains (26 kDa) for ColG, are often associated with the polycystic kidney disease-like domain that precedes them, that is, S2a+S2b (20 kDa) for ColH and S2 (10 kDa) for ColG (Fig. 1D).
Interestingly, the S2b+S3 fragment of ColH binds to type I collagen in a biphasic manner, with a strong affinity interaction (KD = 339 nM) followed by a moderate affinity interaction at higher concentrations (KD = 2110 nM), 85 whereas the S3 domain alone poorly binds to it (KD = 15,900 nM). 85
Another interesting CBD is the short fragment of ColG corresponding to the S3a+S3b domains (Fig. 1D), which binds to all collagen types from I to IV. This domain even binds to the collagen-like peptide (POG) n , provided that the number of triad repeats, n, is large enough for the peptide to adopt a triple-helical conformation, 86 although weakly (KD = 63,000 nM). 87 Likewise, it does not bind to gelatin, further suggesting that this CBD recognizes the triple-helical structure of collagen.84,87
Decorin
Decorin is a small proteoglycan that interacts with collagen fibrils in all connective tissues. 88 Similarly to biglycan and fibromodulin, its core protein is mainly constituted of leucine-rich repeats (LRR). Although the glycosaminoglycan chains play a role in the binding of decorin to collagen, the core protein alone also binds to all types of collagen (I–VI) with strong affinity (e.g., KD = 6 nM for type I collagen).89–91 Major collagen binding sites were identified in a CBD composed of LRR 5 and 6 of the protein (NP_001911.1, Fig. 1E) that binds to type I collagen with moderate affinity (EC50 = 7400 nM, Table 1).88,89 Moreover, an 11-residue subdomain of this CBD, SYIRIADTNIT, showed promise as a short CBD, given its capacity to inhibit the interaction between decorin and type I collagen (Ki = 4000 nM). 89
Staphylococcus aureus adhesin
S. aureus is a very common opportunistic pathogen that persistently colonizes about 20% of the human population. 92 As a gram-positive bacterium, S. aureus is covered with adhesive proteins, among which are adhesins. 93 CNA35 is a 35-kDa fragment of the S. aureus adhesin (CNA) that hooks the bacteria to collagen. This CBD is composed of two domains: N1 and N2, which can entwine the collagen triple helix (Fig. 1F). 94 The interaction of CNA35 with the type I collagen shows a biphasic behavior characterized by a high-affinity binding mode (KD = 500 nM) and a low-affinity mode (our analysis indicated an EC50 > 300,000 nM). 95 To a lower extent, this CBD also binds to collagen II, III, and IV, but neither to collagen V nor to collagen VI. 95
Vibrio mimicus metalloprotease
V. mimicus is a pathogenic bacterium that is responsible for certain cases of gastroenteritis. It expresses a metalloprotease named VMC and that binds to type I, II, and III collagens, in addition to gelatin. 96 A 33-residue-long CBD was identified, LVLSRPGQFAQWAQTVKNLGEQYNAEFAVWLDT (3.8 kDa), which has a moderate affinity for type I collagen (KD = 4000 nM). 97 Interestingly, this CBD contains two FAXWXXT repeats, which were shown to be very important for its collagen-binding capacity. The second repeat in particular is well conserved across several species of Vibrio. 97
Engineered collagen-binding peptides
Several collagen-binding peptides were identified by phage display 98 or by ribosome display, an in vitro method for selection and evolution of peptides (unpublished results). 99 Another strategy to engineer a CBD based on the concept of sense–antisense peptide recognition, from which TKKTLRT ensued. This peptide was engineered as the antisense peptide of the collagenase cleavage site within the α2 chain of type I collagen. Interestingly, TKKLRT has almost the same hydrophilicity plot as the SQNPVQP and SSNPIQP sequences, both of which are found in a collagenase (the matrix metalloproteinase 1 and the neutrophil collagenase, respectively). 100 TKKTLRT can bind to gelatin, 101 but has a moderate affinity for type I collagen (our analysis indicated an EC50 of 2300 nM, Table 1). 100 In addition, Fukata et al. demonstrated that its maximal binding capacity to type I collagen is significantly weaker than that of the ColH-derived CBD that comprises the S2b+S3 domains. 102 Similarly to the PlGF-derived CBD, TKKTLRT features a very high isoelectric point (pI = 11.2), and a scrambled counterpart that did not bind to collagen, namely LTTKKR, was identified. 100 The data thus indicated that the TKKTLRT-collagen interaction is sequence dependent and not solely electrostatic.
Collagen-mimetic peptides
The collagen-mimetic peptides (CMPs), also known as collagen hybridizing peptides, are composed of 6–10 repeats of the GPO or GPP triads, where O is the hydroxyproline. 103 These peptides mimic the triple helix conformation of collagen and have a high propensity to hybridize with collagen, both in vitro and in vivo. 103 More specifically, CMPs can interact with the unfolded strands of denatured collagen to reform a triple-helical structure. 104 On account of their capacity to target unfolded collagen, CMPs are of prime interest for the treatment of pathological conditions that cause collagen degradation, such as cancer, atherosclerosis, arthritis, and fibrosis. 98 CMPs have demonstrated high specificity and high affinity for type I–V collagens, with dissociation constants as low as 10 nM, depending of the number of triad repeats. 105 However, since they spontaneously self-assemble into helical homotrimers, CMPs require heating prior use so as to favor a monomeric state and thus to enable their interaction with denatured collagen. To circumvent this issue, Wahyudi et al. have modified a (GPO) n sequence with a photocleavable group that prevents multimer formation, until the group is removed using UV-irradiation. 98
Applications of GFs Fused to CBDs
Numerous CBDs have been either discovered or engineered over the last decades, which provides the research community with a remarkable toolbox to target collagen for therapeutic applications. This collection of peptide sequences has been efficiently exploited for the stable attachment of proteins and cells to collagen, be it in scaffolds or in vivo. In fact, many recombinant collagen-binding proteins have been reported in the literature, most of which are GFs or cell-binding domains.
Regarding their production, bacterial expression has been the most common platform for these recombinant proteins because of the high yields that can be reached, even though bacteria produce nonglycosylated proteins. Indeed, all of the previously described CBDs, except those derived from FN and decorin, are naturally unglycosylated and can therefore be expressed in Escherichia coli without any loss of collagen-binding activity.55,67,106 In the specific case of FN, it is not clearly established whether the glycosylation of the Asn511 residue within the module I8 significantly impacts its affinity for collagen.107–109 Altogether, bacterial expression is generally suitable as far as the collagen-binding moiety is concerned. It is, however, crucial to consider whether the glycosylation state of the fused protein of interest affects its biological activity to select the appropriate expression platform.
Clinical applications
In the fields of regenerative medicine and tissue engineering, GFs are of prime interest to direct cell fate and control tissue formation. Their very short half-life in vivo is, however, a major drawback that limits the efficiency of bolus injections, 110 a problem that is sometimes addressed by their daily administration. 111 In an effort to prevent the diffusion of the GFs, while maintaining their bioactivity, chimeric proteins corresponding to GFs fused to a CBD have been developed (Fig. 2A). Among other applications, these fusion proteins can be administered to accelerate and extend the repair of cardiovascular wounds and skin wounds, such as ulcers and diabetic wounds. Accordingly, they can also benefit chirurgical reconstructions, such as the uterus, bladder, extrahepatic bile duct, urethra, and abdominal wall, as described in Table 2. Moreover, CBD-GF fusion proteins can be used to induce bone formation and thus to aid fracture healing or to improve the success of a bone graft. They can also be used in the treatment of critical-sized bone defects. For example, mandibular and calvarial or to induce vertical bone regeneration, critical in dentistry (Table 3).
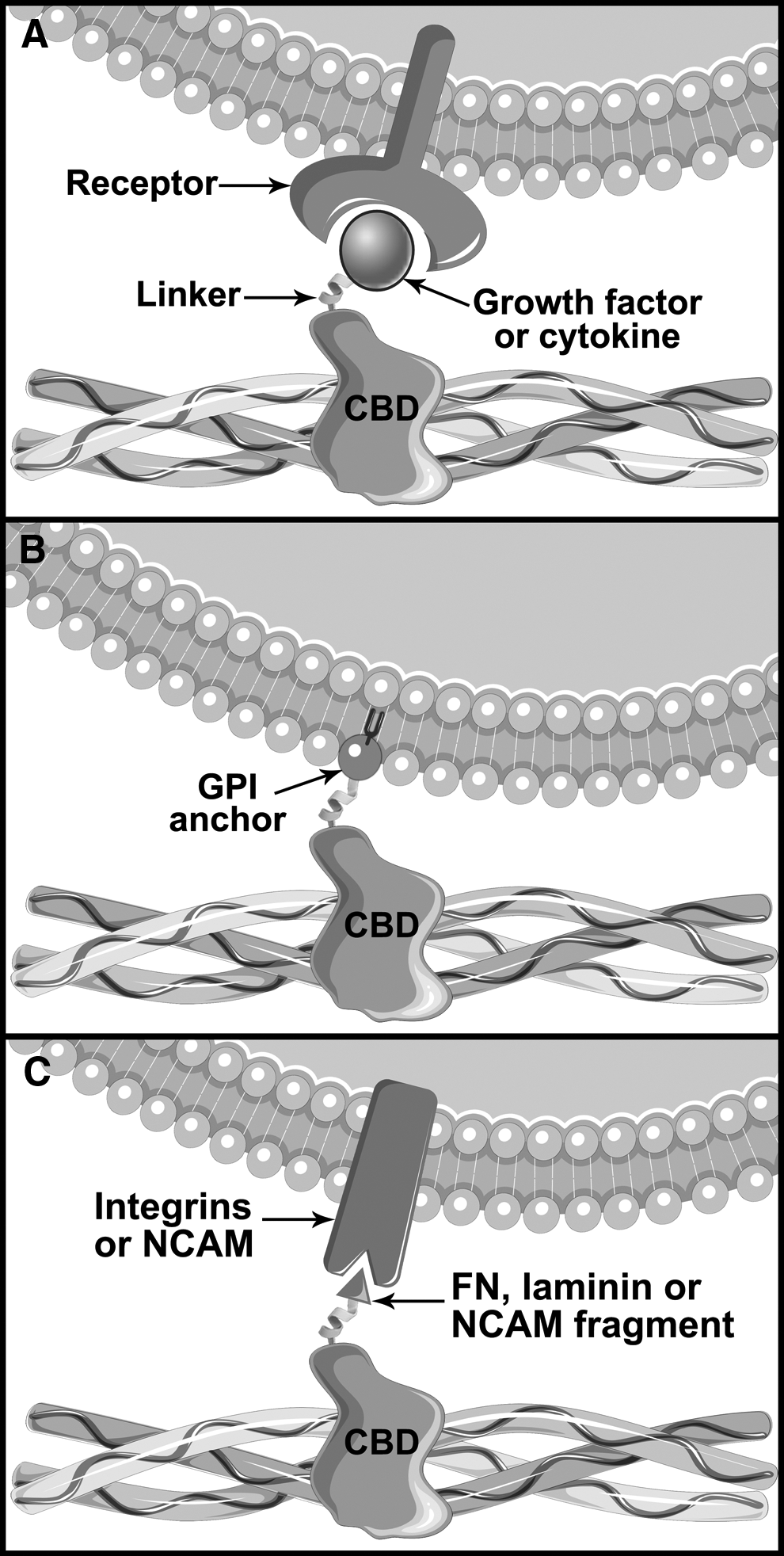
Biological effect of various chimeric proteins containing a CBD:
The tick and cross symbols indicate whether the fusion of a CBD domain to the protein under study was beneficial or detrimental when used instead of the wild-type protein, respectively.
EPC, endothelial progenitor cell; GF, growth factor.
The tick, cross and square symbols indicate whether the fusion of a CBD domain to the protein under study was beneficial, detrimental or had no impact when used instead of the wild-type protein, respectively.
DBM, demineralized bone matrix; ECM, extracellular matrix; MBM, mineralized bone matrix; MSC, mesenchymal stem cell; TGF, transforming growth factor.
GFs fused to a CBD have generally provided better clinical results when compared to their native counterparts, which can be attributed to the increased bioavailability enabled by the collagen-binding activity. In most cases, the delivery of the fusion proteins is improved when they are combined with a proper carrier, which sustains their retention at the site of administration for several days. Most of the carriers are collagen derivatives that are approved by the FDA, and they can meet most clinical requirements given their variety of forms: sponge, hydrogel, membrane, powder, demineralized bone matrix (DBM) or mineralized bone matrix (MBM), and even the CMP (POG)10 (Tables 2–4). Interestingly, no carrier seems to be required in the specific case of vascular applications (Table 2).
The tick, cross and square symbols indicate whether the fusion of a CBD domain to the protein under study was beneficial, detrimental or had no impact when used instead of the wild-type protein, respectively.
GPI, glycosylphosphatidylinositol; OC, osteocalcin; PCL, polycaprolactone.
It is here worth mentioning that several GFs, such as the high-molecular-weight isoforms of VEGF, naturally possess a heparin-binding domain. 112 This feature can be used to improve their retention, be it on a heparinized biomaterial or via their capture by endogenous heparan sulfate. 113 Nevertheless, in most studies, the administration of CBD-VEGF fusions yielded better results than that of native VEGF165 (Table 2). Similarly, bFGF naturally binds to collagen and gelatin 114 but CBD-bFGF fusions provided better outcomes (Tables 2 and 3).
Chimeric collagen-binding GFs for wound healing
Wound healing is a complex process comprising (1) an initial inflammatory phase that involves the clotting cascade and the recruitment of neutrophils and macrophages, (2) a secondary proliferative phase that is characterized by a strong angiogenesis, fibroplasia, formation of granulation tissue, and collagen deposition, and (3) a remodeling phase. 115 Throughout this process, GFs play crucial roles such as chemoattractants (EGF, FGF-1/2, PDGF, TGF-β, SDF-1α),116,117 mitogens (EGF, FGF-1/2, HGF, PDGF),116,118 and promoters of angiogenesis (FGF-1/2, NGF-β, PDGF, TGF-β, VEGF).111,119 All of these factors are released in a highly controlled manner for an optimal spatiotemporal distribution leading to the appropriate modulation of cell response. 118 Hence, providing exogenous GFs may accelerate or improve the quality of the wound healing process, although the success of the approach greatly depends on the carrier used for their topical delivery. 120 For example, it has been long known that the topical application of EGF may accelerate the rate of epidermal regeneration in the cases of partial-thickness skin wounds or chronic wounds.12,121 Several designs of collagen-binding GFs are reported in Table 2, which highlights their typical application and the advantages of their use over native GFs.
Chimeric collagen-binding GFs for bone regeneration
Bone regeneration is an intricate process regulated by numerous cytokines and GFs. Similarly to wound healing, these bioactive molecules play different roles, from chemoattraction (BMP, FGF, PDGF, VEGF, TGF-β) 122 and cell proliferation (FGF, IGF, PDGF, TGF-β) 122 to osteoinduction, that is, differentiation of mesenchymal stem cells (MSC) into mature bone cells (BMP, IL-11, TGF-β). 122 To promote osteogenesis, exogenous GFs can thus be used to functionalize scaffolds such as DBM (made of spongy bone treated with an acid) or MBM (made of spongy bone treated with an oxidizing process).33,123 It is here worth noticing that some of these GFs, such as BMP2, along with several resorbable biomaterials based on type I collagen, are already approved by the FDA for bone regeneration purposes.124–126 In this context, collagen-binding fusion proteins are very appealing to better control the localization of GFs, and numerous chimeras were shown to improve bone regeneration when compared to their native counterparts (Table 3).
Side effects of chimeric collagen-binding GFs
Diffusible GFs feature a short half-life in vivo due to their sensitivity to denaturation, oxidation, or proteolysis. 23 Their translation as clinical tools thus requires high doses, sometimes repeated on a daily basis, to be provided to the site of action to induce the desired effect. 111 Supraphysiological concentrations of native GFs are, however, known to trigger unwanted side effects, both locally and systemically, limiting the amount that can be administered.14,127 For instance, heterotopic bone formation and hypercalcemia were observed with BMP 33 and PTH 128 injections, respectively, and both GFs increased the risks of cancer. Similarly, intimal hyperplasia can arise when using HGF, 129 whereas VEGF is prone to cause hypotension and edema, see heart failure. 130
Given their biology, the use of recombinant CBD-GF fusions may trigger similar side effects. Nonetheless, their binding to collagen fibers hinders both their diffusion and degradation, thus allowing for higher local concentrations, lower systemic distribution, and prolonged activity than their native counterparts. Lower doses can thus be administered to obtain a similar effect. For instance, Visser et al. were able to trigger bone formation in a rat model with 400 ng of a CBD-BMP2 fusion protein, 75 while it is generally admitted that 460 ng is the required minimum. 131 More interestingly, the fusion of a GF with a CBD may even abrogate systemic effects, as observed with VEGF and PTH (Tables 2 and 3).
Influence of the CBD
The collagen-binding ability of a fusion protein greatly depends on which CBD is fused to the GF. Several studies have shown that using a CBD that features a strong affinity for collagen, that is, a low KD, will remain longer at the site of administration, hence providing better healing potential. For example, TKKTLRT-bFGF was shown to promote a stronger neovascularization than WREPSFMALS-bFGF, 132 in good agreement with TKKTLRT displaying a higher affinity for collagen than WREPSFMALS (Table 1). Similarly, bFGF fused to the S2b+S3 domains of ColH markedly enhanced bone formation in comparison with bFGF fused to the S3 domain alone, 133 which has a lower affinity for collagen (Table 1).
Notwithstanding, opposite results were obtained with CBD-PTH chimeras: the PTH-S2b+S3 fusion protein was not as effective as the PTH-S3 fusion protein in improving the bone mineral density of mice. As an explanation, the authors suggested that either the fusion protein was cleaved in vivo within the S2b domain, thus causing its early release, or that the increase in collagen-binding activity caused its sequestration on collagen, thus reducing the bioavailability of PTH to its receptor. 134
Applications of Other Proteins Fused to CBDs
For regenerative medicine or tissue engineering applications, notwithstanding the fact that GFs can promote cell growth, proliferation, and differentiation, the efficient recruitment of the cells to the tissue may require an additional stimulus. In that endeavor, cell adhesion peptides can be fused to a CBD and administered to promote the colonization of a collagen-based biomaterial. Indeed, the migration of cells such as MSC is enhanced when their integrins bind to partners such as the RGD sequence135,136 and various laminin domains. 137 Interestingly, besides providing anchorage, this type of interactions can also promote cell survival, differentiation, and proliferation. 136
As opposed to the recruitment of endogenous cells toward an implant, cell therapy focuses on the administration of exogenous cells. One of the main challenges in the field is to deliver those cells at a specific site, which proves to be challenging when the cells are administered intravascularly. Consequently, providing the cells with a collagen-binding ability could allow for their guidance into the bloodstream toward exposed endogenous collagen, for example, at the site of a vascular injury to improve re-endothelialization. Such a feat could be achieved thanks to a fusion protein that comprised a CBD and a glycosylphosphatidylinositol (GPI) anchor (Table 4). More specifically, when cells are incubated with a CBD-GPI chimera, the latter penetrates their lipid bilayer, thus providing them with a collagen-binding capacity, as depicted in Figure 2B. 138 Interestingly, CBD-adhesion peptide fusions can also be used for cell therapy when combined with a collagen-based carrier, as a mean to increase the survival rate of transplanted cells, for example, in the case of neural stem cell implantation in the brain. 139
Conclusion
In this review, we have highlighted the therapeutic potential of fusion proteins comprising a CBD, with a focus on GFs combined with collagen-based or collagen-coated scaffolds, for wound healing and bone regeneration applications. Numerous in vivo studies have indeed indicated that the fusion of a given GF to a CBD led to a drastic improvement of its therapeutic effect when compared to that of the native GF administered in its soluble form. This enhancement is most likely attributable to a prolonged bioavailability of the GF, be it due to the fact that the GF is preserved from degradation or that it is present for a longer period of time and at a higher local concentration in the vicinity of its cell surface receptors. Of interest, the use of a chimeric CBD-GF may also limit the occurrence of the systemic side effects that are observed with several diffusible GFs, which can again be explained by their efficient sequestration at the site of injury. The best example of this beneficial effect is arguably VEGF, which fusion to a CBD enabled the uncoupling of its angiogenic role from its hyperpermeability activity.
In most of the references collated in this review, researchers report the benefits of a given CBD-GF fusion protein when added to a single collagen-based scaffold. Despite all the promising results presented therein, these data remain hard to interpret as a whole to establish the importance of the biophysical traits of the interaction between the CBD and the collagen-based scaffold (e.g., GF loading capacity, binding affinity, and stability) on therapeutic outcomes.
Indeed, the amount of the chimeric protein bound to the scaffold or the impact of varying this quantity has seldom been reported. On the same note, the effect of replacing a CBD by another is merely addressed. A significant amount of work thus remains to be done to better characterize the interactions occurring between the various types of collagen and the different CBDs identified so far. These experiments are of prime importance, not only to resolve the discrepancies between several of the values listed in Table 1 but also to provide the research community with a broader knowledge of these interactions. We believe that such a characterization, if exhaustive, would guide the design of novel tailored CBD-GF chimeras for optimal therapeutic effects.
Footnotes
Acknowledgments
We apologize to authors whose relevant articles were not cited due to space limitations. This work was supported by the Canada Research Chair on Protein-Enhanced Biomaterials, by the Natural Sciences and Engineering Research Council of Canada, and by the MEDITIS training program.
Disclosure Statement
No competing financial interests exist.