
Abstract
The regenerative potential of tissue-engineered bone constructs may be enhanced by in vitro coculture and in vivo cotransplantation of vasculogenic and osteogenic (progenitor) cells. The objective of this study was to systematically review the literature to answer the focused question: In animal models, does cotransplantation of osteogenic and vasculogenic cells enhance bone regeneration in craniofacial defects, compared with solely osteogenic cell-seeded constructs? Following PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines, electronic databases were searched for controlled animal studies reporting cotransplantation of endothelial cells (ECs) with mesenchymal stem cells (MSCs) or osteoblasts in craniofacial critical size defect (CSD) models. Twenty-two studies were included comparing outcomes of MSC/scaffold versus MSC+EC/scaffold (co)transplantation in calvarial (n = 15) or alveolar (n = 7) CSDs of small (rodents, rabbits) and large animal (minipigs, dogs) models. On average, studies presented with an unclear to high risk of bias. MSCs were derived from autologous, allogeneic, xenogeneic, or human (bone marrow, adipose tissue, periosteum) sources; in six studies, ECs were derived from MSCs by endothelial differentiation. In most studies, MSCs and ECs were cocultured in vitro (2–17 days) before implantation. Coculture enhanced MSC osteogenic differentiation and an optimal MSC:EC seeding ratio of 1:1 was identified. Alloplastic copolymer or composite scaffolds were most often used for in vivo implantation. Random effects meta-analyses were performed for histomorphometric and radiographic new bone formation (%NBF) and vessel formation in rodents' calvarial CSDs. A statistically significant benefit in favor of cotransplantation versus MSC-only transplantation for radiographic %NBF was observed in rat calvarial CSDs (weighted mean difference 7.80% [95% confidence interval: 1.39–14.21]); results for histomorphometric %NBF and vessel formation were inconclusive. Overall, heterogeneity in the meta-analyses was high (I2 > 80%). In summary, craniofacial bone regeneration is enhanced by cotransplantation of vasculogenic and osteogenic cells. Although the direction of treatment outcome is in favor of cotransplantation strategies, the magnitude of treatment effect does not seem to be of relevance, unless proven otherwise in clinical studies.
Introduction
R
The bone tissue engineering (BTE) approach involves harvesting of osteogenic cells (most commonly mesenchymal stem cells [MSCs]) from an autologous source (e.g., bone marrow, adipose tissue), their in vitro expansion, and, in combination with an appropriate carrier scaffold, their in vivo implantation. 7 Thus, the triad of osteogenic cells, osteoinductive signals (growth factors [GFs] released by cells), and osteoconductive scaffolds replicates the properties of autogenous bone, without the need for invasive harvesting. 8 The prospects of BTE for alveolar bone repair are very promising, as demonstrated by several preclinical and some clinical studies (for reviews, see Shanbhag and Shanbhag, 7 Shanbhag et al., 9 and Padial-Molina et al. 10 ).
The promising results of preclinical BTE studies have not yet translated into significantly improved clinical outcomes in human trials.11–13 A major limitation of traditional BTE approaches is the reported lack of adequate and timely vascularization of the implanted construct—which is essential for oxygenation, nutrition, and waste elimination to/from the cells. 14 Absence of adequate vascular supply can result in premature cell death in regions of the construct distant from the host vasculature since diffusion of oxygen and nutrients is only limited to a distance of 150–200 μm.15,16 One strategy to overcome this limitation is to coculture (in vitro) and cotransplant (in vivo) osteogenic cells along with endothelial cells (ECs) or endothelial progenitor cells (EPCs) to simultaneously stimulate osteogenesis and vasculogenesis, that is, de novo formation of blood vessels, within bone constructs.17,18
EPCs and ECs are hematopoietic cells, which contribute directly to the process of new vessel formation, 17 and can be easily isolated from peripheral blood, bone marrow, and other tissues. 19 These cells also play a central role in emerging prevascularization strategies, which aim to generate primitive vessels within tissue constructs in vitro that functionally anastomose with the host vasculature when implanted in vivo.20,21 Recent studies have demonstrated enhanced performance of osteogenic cells (MSCs or osteoblasts [OBs]) when cocultured with EPCs or ECs in terms of in vitro gene expression and in vivo bone formation, suggesting a synergistic effect of vasculogenesis and osteogenesis—as discussed in several recent review articles.22–25
Preclinical testing of new regenerative therapies in clinically relevant animal models is an important aspect of translational research and, in most cases, a requirement of regulatory health agencies before initiating human clinical trials.26,27 Commonly reported animal models of relevance to craniofacial bone regeneration involve calvarial and alveolar critical size defects (CSDs). A CSD is the smallest size experimental defect in the cranial or alveolar bone of the animal that will not spontaneously and extensively regenerate with bone within a defined time frame without intervention.28,29
Following the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-analyses) guidelines, 30 the aim of the present study was to systematically review the available literature to answer the focused PICO (population, intervention, comparison, outcome) question: In CSDs of experimental animals, does a cotransplantation approach, that is, implanting osteogenic and vasculogenic cells seeded on biomaterial scaffolds, enhance bone regeneration compared with implanting only osteogenic cell-seeded scaffolds?
Materials and Methods
Inclusion and exclusion criteria
Inclusion criteria
(1) English-language articles.
(2) Randomized or nonrandomized, controlled preclinical studies with two or more experimental groups.
(3) Involving CSDs in the calvaria, maxillae, or mandibles of small or large animals.
(4) Cotransplantation of cultured autologous, allogeneic, or human-derived MSCs (or OBs) and EPCs (or ECs) seeded on biomaterial scaffolds in at least one experimental group.
(5) A control group receiving transplantation of only MSC- or OB-seeded scaffolds.
(6) Quantitative assessment of new bone formation (%NBF) 29 through (a) histomorphometry or (b) three-dimensional (3D) computed tomography (CT) or micro-CT. Quantitative histological or immunohistochemical assessment of new vessel formation (vasculogenesis) was considered as a secondary outcome.
Exclusion criteria
(1) Solely in vitro studies.
(2) In vivo studies reporting CSDs in other anatomical sites (e.g., long bones), ectopic models (e.g., subcutaneous), or systemic cell delivery.
(3) Absence of a cotransplantation group or an MSC-only group.
Search strategy, screening, and study selection
Electronic databases of MEDLINE (via PubMed) and EMBASE were searched for relevant English-language literature up to and including April 2016. Unpublished literature was searched through the Google and Google Scholar search engines. Bibliographies of the selected studies and relevant review articles were checked for cross-references. A specific search strategy was developed for MEDLINE (Table 1) and adapted for other databases. Titles and abstracts of the search-identified studies were screened by two authors (S.S. and A.S.) and full texts of all eligible studies were obtained. Uncertainty in the determination of eligibility was resolved by discussion with the other authors. Two authors (S.S. and A.S.) reviewed the selected full texts independently, and final inclusion was based on the aforementioned inclusion criteria. A summary of the screening process is presented in Figure 1.
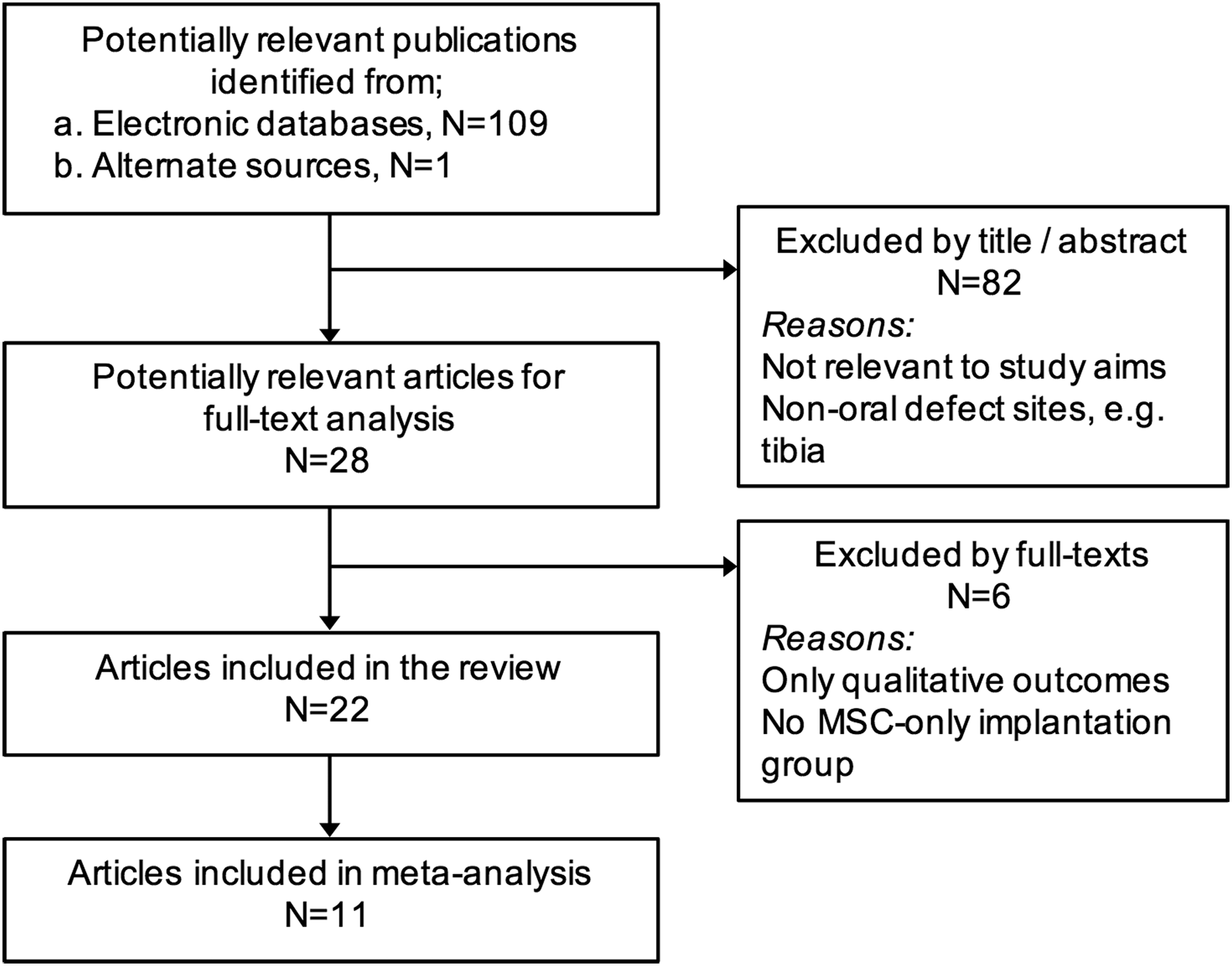
Flowchart for study screening and selection. N, number of articles.
Data extraction
Data were extracted from full texts of included articles on author(s), study design, animal species, model characteristics, number of animals, number of procedures, inclusion criteria, outcome(s), method(s) of outcome evaluation, main findings, and conclusions. Descriptive summaries of studies were entered into tables. Quantitative data regarding NBF, that is, regenerated bone volume as a percentage, and vessel formation, that is, vessel numbers/density per unit area, were extracted from studies for possible meta-analyses. When data were presented as ratios or volumes (mm3), they were converted to percentages using total defect volume data. Standard errors of means, when reported, were converted to standard deviations for analysis. If data were only expressed graphically, numerical values were requested from the authors, and if a response was not received, digital ruler software was used to measure graphical data (ImageJ; National Institutes of Health, Bethesda, MD). When studies reported outcomes at multiple time points, data from similar time points of different studies were pooled for meta-analysis.
Quality assessment and risk of bias
Reporting quality assessment of all studies was performed based on a modification of the ARRIVE (Animal Research: Reporting In Vivo Experiments) guidelines regarding relevant items.31,32 Compliance with the guidelines was evaluated using a predefined grading system applied to each of the 20 items 33 (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/teb). Reporting quality was judged as high, moderate, or low. Risk of bias (RoB) assessment was performed using a modification of the SYstematic Review Centre for Laboratory animal Experimentation (SYRCLE) RoB tool for animal studies 34 and judged as high, low, or unclear 35 (Supplementary Table S2). Any disagreement between the reviewers during study selection, data extraction, and quality assessment was resolved by discussion and consensus.
Meta-analysis
Meta-analysis was performed to compare the effectiveness of MSC/EC cotransplantation (experimental [Ex]) and MSC-only transplantation (control [Co]) using histomorphometric and micro-CT data (mean and standard deviation of %NBF and number of animals/defects [n] per group). Studies were pooled based on homogeneity regarding PICO and observation time. Subgroup analyses were performed at the level of animals and observation time(s), using the DerSimonian and Laird random effects model 36 and STATA Statistical Software (StataCorp LP, College Station, TX). In one study, 37 the design included multiple interventions per animal, that is, bilateral calvarial defects, for which a correlation coefficient (r) was calculated for between treatment-group measurements. 38 An r-value of 0.5 was used for this study; for parallel group studies, the r-value was set at 0 and thus both study types were included in the same analysis. 39 Pooled estimates of treatment effect (weighted mean differences [WMDs]) were calculated along with 95% confidence intervals (CIs), and the I 2 statistic was used as a measure of inconsistency of results across studies. 36
Results
Search results and study characteristics
Of the 110 search-identified titles, 22 studies reporting quantitative histomorphometric and/or radiographic (CT/micro-CT) outcomes from 5 different species and 474 animals were included in the review (Table 2). A list of excluded studies along with reasons for exclusion is provided in Supplementary Table S5. Small animal models included rats (number of animals [n] = 356), mice (n = 77), and rabbits (n = 27). Large animal models included minipigs (n = 6) and dogs (n = 8; Table 2). Sample sizes in individual studies ranged from 2 to 64, and observation times ranged from 4 to 12 weeks. Fifteen studies used calvarial CSD models, while seven studies used maxillary (two studies) or mandibular CSDs (five studies). No studies included a positive control group treated with gold standard autogenous bone.
Indirect coculture of MSCs and EPCs, that is, MSCs cultured in the presence or absence of EPCs were implanted.
μCT, microcomputed tomography; Allo, allogeneic; Auto, autologous; (b), bilateral CSD; CCSD, calvarial CSD; Cells, number of implanted MSCs; coratio, time, coculture (MSC:EC) ratio and coculture time; CSD, critical size defect; CT, computed tomography; EPCs, endothelial progenitor cells; Histo, histomorphometry; Human, human-derived; induction, osteogenic induction; IS, immune suppressed; MCSD, mandibular CSD; MxCSD, maxillary CSD; n, number of animals; NR, not reported; NZ, New Zealand; ON, overnight; SCID, severe combined immunodeficiency; SD, Sprague-Dawley; Vessels, vessel regeneration.
Cells (osteogenic): ASC, adipose tissue-derived MSC; BMSC, bone marrow-derived mesenchymal stem cell; HOB, human osteoblast; POC, periosteal cell.
Cells (vasculogenic): ASC-EC, ASC-differentiated EC; AT-EC, adipose tissue-derived EC; BMSC, bone marrow-derived MSC; BMSC-BMP2, BMP2 gene-modified BMSC; BMSC-EPC/EC, BMSC-differentiated EPC/EC; BMSC-SMC, BMSC-differentiated smooth muscle cell; DMEC, dermal microvascular EC; EC, endothelial cell; EPC-BMP2, BMP2 gene-modified EPC; HUVEC, human umbilical vein EC; PB-EPC, peripheral blood EPC; PO-EC, periosteum-derived EC; UCB-EPC, umbilical cord blood-derived EPC.
Scaffolds: DBBM, demineralized bovine bone mineral; HA-PLA-Col/Fn (ML), HA-PLA-collagen/fibronectin (multilayered scaffold); HA-PLGA, hydroxyapatite-PLGA; Nano-CS/Alginate, nano-calcium sulfate/alginate; PCL/PLGA/TCP, polycaprolactone/PLGA/tricalcium phosphate; PDO/PLF, polydioxanone/pluronic F127 copolymer; PLGA, poly-lactide-co-glycolide; Poly(LLA-co-DXO), poly(
Quality assessment and RoB
Overall, the included studies were of unclear to high RoB and moderate reporting quality (Supplementary Tables S2 and S3). Most studies provided adequate information regarding title, abstract, introduction, study objectives, ethical approval, experimental design, and procedures. However, information regarding experimental animals and their housing and husbandry was generally inadequate. No studies provided information on sample size calculation or baseline characteristics of the animals. In seven studies, animals (or defects) were randomly allocated to treatment groups to minimize selection bias, although no details of the randomization procedure were reported. Four studies reported blinding of outcome assessors to treatment groups to minimize detection bias. All studies reported detailed outcome evaluation, including statistical analyses. Finally, information regarding study limitations and implications for translation to human models was limited. None of the studies reported implications for the 3Rs principle (replacement, refinement, and reduction) of animals in research.31,40
Characteristics of cell cotransplantation strategies
Osteogenic cells
Included studies reported on the use of autologous, allogeneic, xenogeneic, or human bone marrow-derived MSCs (BMSCs), adipose tissue-derived MSCs (ASCs), periosteal cells (POCs), or human osteoblasts (HOBs) as the osteogenic cell population. Eleven studies reported the use of human-derived cells, that is, BMSCs,41–44 ASCs,45–47 HOBs, 48 or POCs.49–51 MSCs were used in early passages (2–5) and most often osteogenically induced before implantation (66% studies). One study 52 reported the use of bone morphogenetic protein-2 (BMP-2) gene-modified allogeneic BMSCs in rats.
Vasculogenic cells
ECs and EPCs in the included studies were obtained from commercial, autologous, allogeneic, or human sources—most commonly, bone marrow. Commercially obtained human umbilical vein endothelial cells (HUVECs) were reported in three studies,42,45,47 while EPC isolation from human umbilical cord blood was reported in two studies.43,51 In six studies, ECs were differentiated from a fraction of the isolated MSCs by endothelial induction.37,52–56 EC/EPCs were characterized by expression of endothelial-specific markers, CD31, CD146, or CD34, through flow cytometry, immunostaining, or polymerase chain reaction (PCR) analysis.
Coculture
A majority (60%) of studies reported in vitro coculture of MSCs and EPC/ECs overnight or for 2–17 days before implantation; in three studies,41,42,53 MSCs and ECs were coseeded on scaffolds only 0.5–1.5 h before implantation. One study 57 reported cell sheet preparation through BMSC-EPC coculture for 10 days, while another study (by the same group) 58 reported indirect coculture of BMSCs and EPCs using a transwell system—to investigate the influence of EPC secretory products on BMSCs. Coculture conditions varied between studies; MSCs and EPC/ECs were cocultured in standard, osteogenic, endothelial, or combination culture media. In two studies,47,52 the optimal seeding ratio of MSC:ECs and the influence of coculture on in vitro osteogenic potential of MSCs were evaluated by alkaline phosphatase (ALP) activity or calcium staining assay after 7 or 28 days, respectively. Coculture enhanced MSC osteogenic differentiation compared with MSC-only culture, and an MSC:EC seeding ratio of 1:1 (vs. 1:2, 2:1, 5:1, etc.) was identified as being most optimal. The number of implanted cells ranged from 1 × 103 to 12 × 106 each for MSCs and EPC/ECs. In one study, 52 coculture of BMP-2-modified BMSCs and EPCs enhanced ALP activity and OB and endothelial marker expression compared with monoculture of BMP-2-modified BMSCs or unmodified BMSCs. In vitro prevascularization, that is, formation of primitive capillary-like networks, following 1-week dynamic coculture of BMSCs and ECs on copolymer scaffolds was reported in one study. 37
Scaffolds
In most studies, alloplastic copolymer or composite scaffolds were used as carriers for cells. Other common biomaterials such as β-tricalcium phosphate (β-TCP), hydroxyapatite (HA), and bovine bone mineral (BBM) were also used as scaffolds. One study reported the use of (rat) calvarial allografts. 54 One study 59 reported the use of a multilayered composite (HA-copolymer) scaffold comprising MSCs and ECs in a layered construct design. Another study 60 reported incorporation of vascular endothelial growth factor (VEGF) within β-TCP scaffolds in some experimental groups. Scaffolds were prepared according to defect dimensions and seeded with cells either well before (2–14 days) or shortly before (1–1.5 h) surgery.
Qualitative outcomes
Overall, minimal signs of inflammatory response, infection, or adverse reactions were observed across all studies, even after transplantation of allogeneic, xenogeneic, or human-derived cells. Detection of newly formed vessels was performed through immunohistochemical staining for endothelial-specific markers such as CD31, CD34, von Willebrand factor (vWF), VEGF, and so on. One study 45 reported PCR analysis of retrieved tissues to identify endothelial marker genes (CD31, vWF, and vascular endothelial-cadherin). A general observation across studies was the detection of vascular structures consistently in areas of NBF. In four studies,41,42,44,45 detection of human-specific endothelial markers in retrieved tissues confirmed the contribution of implanted ECs in vasculogenesis.
Meta-analyses
Meta-analysis was performed if there were three or more studies in each comparison group. Eleven studies reporting histomorphometric or micro-CT-based %NBF following MSC/EC cotransplantation versus MSC-only transplantation, in calvarial CSDs of rats and mice, were included in two separate meta-analyses (Fig. 2). A comparison was also made between cotransplantation and EC-only transplantation (Fig. 3). Finally, vessel regeneration was compared between cotransplantation and MSC-only groups (Fig. 4). To minimize any confounding influence of exogenous GFs, experimental groups receiving gene-modified cells and/or GFs were excluded. Since there were fewer than 10 studies in each meta-analysis, publication bias through funnel plots or statistical testing was not assessed due to the lack of power to distinguish chance from real asymmetry. 61
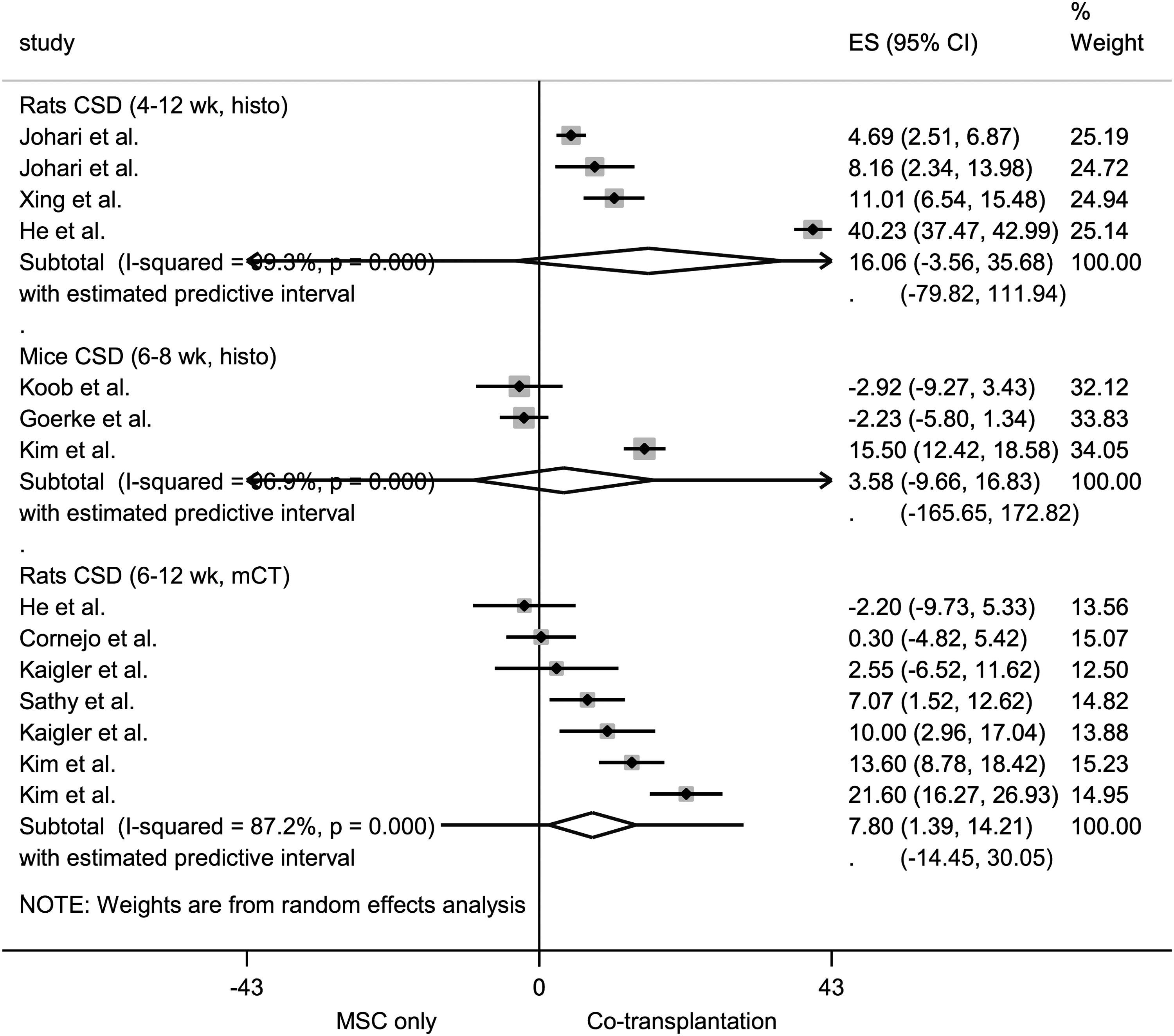
Forest plot for the comparison cotransplantation versus MSC-only transplantation for bone formation in rodent CSD models. The forest plot displays relative weight of the individual studies, the estimates of treatment effect (ES, effect size) expressed as WMDs, 95% CIs, and a predictive interval for the outcome %NBF. The diamond indicates the pooled estimate and its 95% CI. μCT, microcomputed tomography-based %NBF; CI, confidence interval; CSD, critical size defect; histo, histomorphometric new bone formation (%NBF); MSC, mesenchymal stem cell; wk, weeks; WMDs, weighted mean differences.
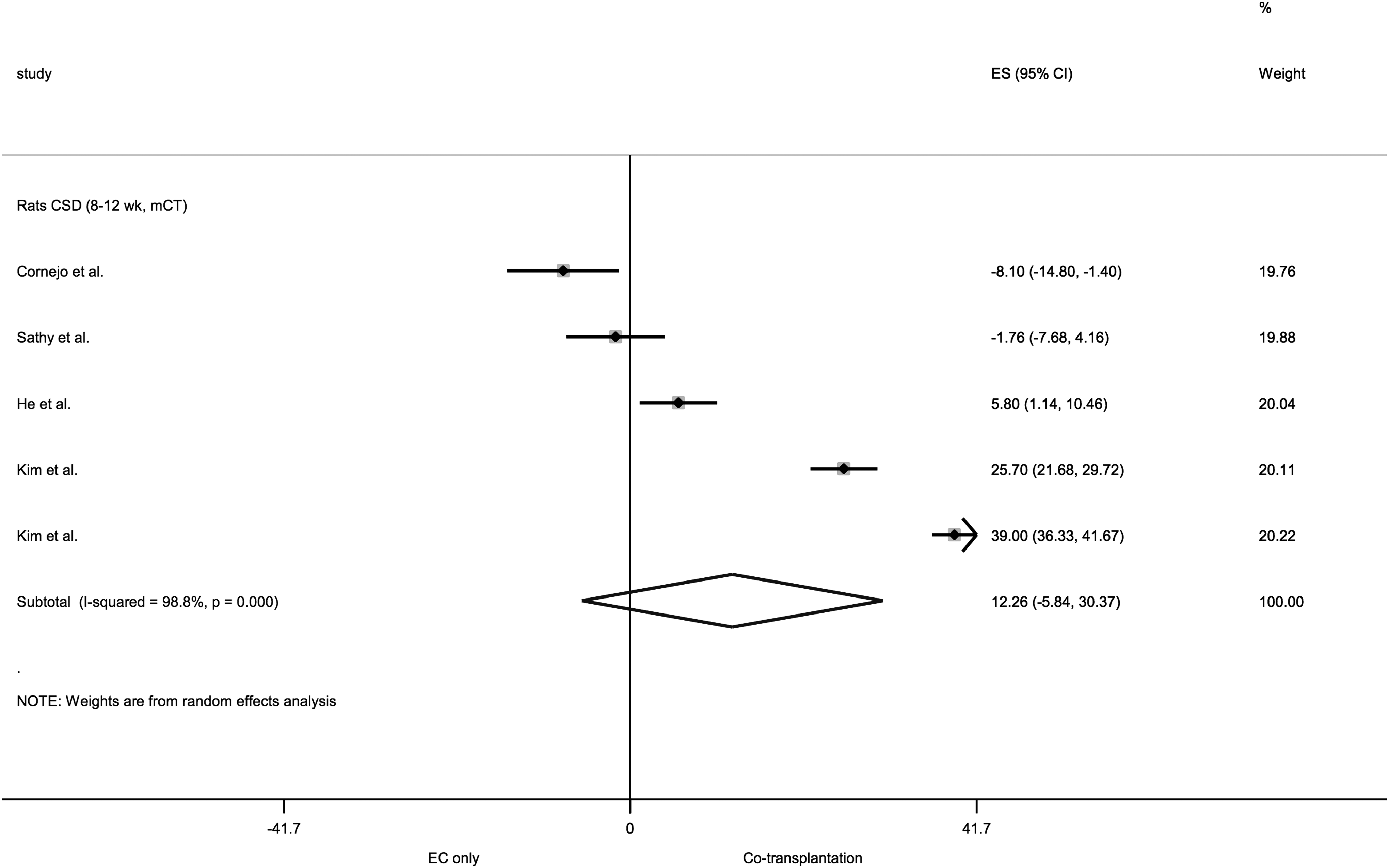
Forest plot for the comparison cotransplantation versus EC-only transplantation for bone formation in rat CSD models. The forest plot displays relative weight of the individual studies, the estimates of treatment effect (ES) expressed as WMDs, 95% CIs, and a predictive interval for the outcome %NBF. The diamond indicates the pooled estimate and its 95% CI. EC, endothelial cell.
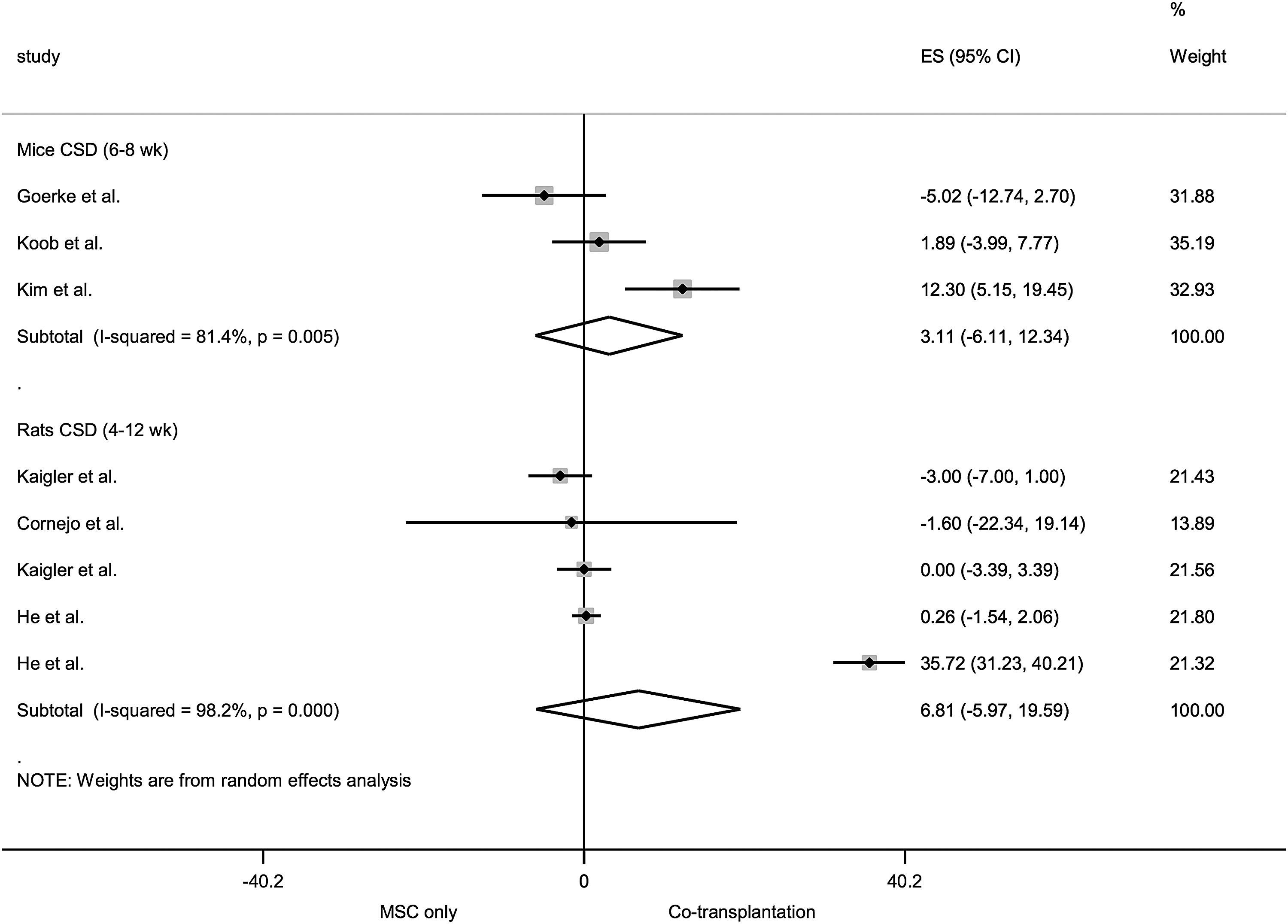
Forest plot for the comparison cotransplantation versus MSC-only transplantation for vessel formation in rodent CSD models. The forest plot displays relative weight of the individual studies, the estimates of treatment effect (ES) expressed as WMDs, 95% CIs, and a predictive interval for the outcome vessel density per unit area. The diamond indicates the pooled estimate and its 95% CI.
The meta-analyses revealed a statistically significant effect in favor of cotransplantation versus MSC-only transplantation for radiographic bone regeneration in rat calvarial CSDs—pooled estimate (WMDs): 7.80% NBF (95% CI 1.39–14.21). Results for histomorphometric bone regeneration and vessel regeneration were inconclusive. Heterogeneity in most cases was very high (I 2 > 80%, p < 0.05). Pooled WMDs with 95% CI and measures of heterogeneity for each of the subgroups are presented in Supplementary Table S4, along with an example for interpretation.
Discussion
The aim of this study was to systematically review the preclinical in vivo evidence for cell cotransplantation strategies for craniofacial BTE. Recent reviews of preclinical in vivo studies have revealed favorable outcomes of BTE approaches for periodontal 35 and alveolar bone defect regeneration. 9 However, a translational limitation of traditional BTE strategies, including implantation of osteogenic cells seeded on biomaterial scaffolds, is the inadequate vascularization of constructs in human clinical defects. 62 Preclinical studies have demonstrated benefits of coculturing and cotransplanting osteogenic and vasculogenic cells in experimental animal defects to overcome this limitation. For the present review, the most commonly reported animal models of relevance to maxillofacial bone regeneration, that is, calvarial and alveolar CSDs, employing cotransplantation strategies, were considered.
Animal models
Small animal models
A majority of studies included in the present review reported on small animal models, that is, rodents and rabbits. Generally, small animal models constitute a starting point for proof-of-principle or feasibility studies before clinical modeling and efficacy testing in larger animals.26,63 Rodent models are often preferred over larger animals due to significantly lower costs, easier housing and handling, and minimal social concern. 64 The calvarial CSD model has been well characterized in rodents—a circular defect of 5 mm diameter is generally considered to be of critical size.29,65 The present meta-analysis revealed significantly higher radiographic bone regeneration following cotransplantation versus MSC-only or EC-only transplantation in calvarial CSDs of rats; results for histomorphometry were inconclusive. Calvarial bone is reported to physiologically resemble the mandible and therefore calvarial CSDs represent a reliable experimental model to test regenerative therapies for maxillofacial applications.66,67 Moreover, rodent calvaria are considered as a challenging environment for bone regeneration due to poor blood supply and limited bone marrow 68 ; thus, the role of vascularization in bone graft healing is even more critical in this zone. 69 In context, recent studies demonstrated that cotransplantation of EPCs with MSCs enhances vertical bone regeneration, compared with cell-free β-TCP scaffolds, in a rat calvarial dome model—corresponding to the guided bone regeneration (GBR) technique.70,71
Rabbits, like rodents, provide advantages of small size and easy handling; additionally, they provide larger volumes of bone tissue and creation of more reliable CSDs than in rodents. 72 The included rabbit studies reported significantly greater histomorphometric %NBF in cotransplantation versus MSC-only groups in either calvarial 55 or mandibular 73 CSD models. However, the significant differences in structure, composition, and physiology of rodent, rabbit, and human bone (e.g., trabecular content, metabolic rate, remodeling) must be considered when extrapolating results from these studies to avoid, for example, overestimation of clinical performance.74,75
Large animal models
Large animal models are especially pertinent in BTE research as defects with relevant diffusion distances simulating the clinical setting can be generated, which allows evaluation of the influence of mass transport, hypoxia, and vascularization on transplanted cell survival. 75 Four studies included in the present review reported data from large animal models—minipigs and dogs. Dogs and pigs, due to the similarities in structure, composition, and physiology between canine/porcine and human bones, are extensively used in musculoskeletal research 76 ; the rate of remodeling in pigs (1.2–1.5 μm/day) is comparable with that in humans (1.0–1.5 μm/day), but slower compared with that in dogs (1.5–2.0 μm/day). 74 Minipigs are often preferred due to easier handling than domestic pigs and morphological similarities to human bone. 77 In the present review, all three minipig studies reported superior radiographic bone regeneration in mandibular CSDs augmented with copolymer scaffolds loaded with human POCs and ECs (derived from adipose tissue, periosteum, or umbilical cord blood) versus scaffolds loaded with only POCs after 12 weeks.49–51 Interestingly, no immunological reactions were reported following implantation of human-derived cells in pigs. The biocompatibility of MSCs within and across species can be attributed to their hypoimmunogenic, immunomodulatory, and anti-inflammatory properties, which have been discussed in detail elsewhere. 78
A single included study 60 in dogs reported significantly greater histomorphometric %NBF following 8 weeks of coimplantation of autologous MSCs and EPCs in composite (β-TCP-PLGA) scaffolds versus only MSC-seeded scaffolds (45.21% ± 0.16% vs. 34.59% ± 1.49%). Interestingly, the investigators also included VEGF—a key regulator of angiogenesis, in some of the experimental groups. The highest %NBF was observed in the MSC/VEGF group (63.42% ± 1.6%). However, no significant differences were observed between the MSC/EPC/VEGF (47.80% ± 1.87%) and MSC/EPC groups (45.21% ± 0.16%), suggesting that no additional benefit of VEGF could be expected when EPCs were used. 60 In context, a recent study reported significantly greater %NBF in femoral defects of sheep, following cotransplantation of dynamically cultured autologous BMSC-derived OBs and ECs (∼88.10% ± 5.22%) versus solely OBs on β-TCP scaffolds (∼17.65% ± 4.50%) after 16 weeks. 79
In the present review, only studies employing craniofacial defects were included; due to differences in the embryonic development, structure, biomechanics/chemistry, and regenerative capacity between craniofacial and long bones, findings from studies using long bone models are not directly transferable to the craniofacial skeleton.80,81 In context, similar to the findings of the present review, several studies using long bone defects in both small82–87 and large animal models79,88 have reported significantly greater bone and vessel regeneration following MSC-EC cotransplantation versus MSC-only transplantation (including human-derived cells).
Coculture: translational considerations
MSCs and EPCs represent the most favorable progenitor cell populations for osteogenic and vasculogenic differentiation within a coculture system. Bone marrow has traditionally been considered the gold standard source for MSCs and EPCs (a fraction of the hematopoietic stem cell population); however, the morbidity and procedural burden associated with marrow harvesting have led to identification of relatively less invasive sources of MSCs (adipose tissue, oral tissues, etc.) and EPCs (peripheral blood, umbilical cord blood, endothelial-induced MSCs, etc.). The ex vivo differentiation of MSCs into functional ECs by endothelial induction—as reported by six included studies—represents a particularly attractive strategy since both cell types are obtained from the same primary source, minimizing the need for additional tissue harvesting. 89
Predifferentiation or induction of progenitor cells before implantation is considered to enhance their in vivo efficacy. 47 The choice of culture media plays an important role in determining the optimal conditions for inducing osteogenic and endothelial differentiation of MSCs and EPC/ECs, respectively, without exerting negative effects on either cell type. 22 In the present review, coculture of MSC-EPC/ECs was performed for 2–14 days before implantation in standard culture medium (SM), osteogenic medium (OM), a 1:1 mixture of SM or OM and endothelial medium (EM), or sequential culture first in EM and then in OM. In one study, 58 a transwell indirect coculture system was used, in which MSCs were exposed to the secretome of EPCs before implantation. Although all studies reported favorable in vivo outcomes, no studies correlated in vivo outcomes with different in vitro culture conditions. However, recent evidence suggests that optimal results may be achieved with coculture of MSC-ECs in purely OM in terms of both in vitro and in vivo bone formation and vascularization.90–92
MSC expansion is commonly performed using basal culture media plus supplements to provide GFs, proteins, and enzymes to support cell growth. Fetal bovine serum (FBS) is commonly used to supplement culture media because the fetal milieu is enriched with growth factors and poor in antibodies. 93 However, for clinical-grade therapy, it is important to substitute animal-derived products because MSCs can internalize xenogeneic proteins and thus there is a risk of infection and immunoreactions. 93 In addition, there are concerns regarding sample-to-sample inconsistency and ethics of animal welfare. Alternatives to FBS include chemically defined media or human blood platelet-derived supplements (e.g., platelet lysate [PL]) to provide the necessary factors for cell growth and function. 93 One study 47 reported on the use of human PL for ASC and ASC/HUVEC cultures and that significantly improved in vitro mineralization (osteogenesis) in PL- versus FBS-supplemented cultures was observed. These results are consistent with reports of BMSC/HUVEC cocultures with PL supplementation. 94
Only one study 37 reported the use of a dynamic coculture system for rat BMSCs and ECs seeded on copolymer scaffolds, with formation of capillary-like structures in vitro after 1 week, suggesting a possible strategy for prevascularization. 95 In context, a recent study 79 in a large animal (sheep) model reported bone regeneration in femoral defects following dynamic coculture of autologous OBs and ECs in a perfusion bioreactor system. Bioreactor systems (spinner flasks, perfusion systems, etc.) are reported to enhance the proliferation, homogeneous distribution, and differentiation of MSCs in scaffolds compared with conventional, static two-dimensional (2D) culture by providing the appropriate inductive and mechanical cues in a controlled environment.96,97 Additionally, bioreactors ensure more homogeneous distribution and perfusion of cells and culture media throughout the scaffold and can reportedly overcome the problems associated with nutrient gradients that develop in static cultures, that is, when cells nearer to the scaffold surface consume more nutrients than cells in the center, resulting in internal cell death. 96
Scaffold materials chosen to deliver cells and/or GFs play a critical role in the performance of BTE constructs. Biocompatible and biodegradable scaffolds can support and facilitate the attachment, proliferation, and differentiation of progenitor cells, ingrowth of vascular structures, and mass transfer while also providing structural stability, space maintenance, and osteoconductivity at the regeneration site.16,22 Commercial bone substitute materials, for example, β-TCP, HA, BBM, and so on, have been shown to be clinically effective scaffolds for MSCs. 7 However, alloplastic copolymer scaffolds have gained significant importance since each of their properties integral to performance (architecture, biodegradability, porosity, pore size, etc.) can be optimized during the fabrication process. 98 Especially in cotransplantation strategies—where the presence of multiple cell types and complex simultaneous processes demand differential scaffold design, copolymer scaffolds provide several advantages. 99 In the present review, a majority of studies reported the use of copolymer composite scaffolds to support MSC monocultures and MSC-EC cocultures with adequate in vitro efficacy and subsequently successful in vivo outcomes.
Efficacy of human-derived cells
The main translational implication of the in vitro findings is that cotransplantation of previously cocultured osteogenic and vasculogenic cells could enhance in vivo bone regeneration either by directly inducing osteogenic differentiation of MSCs or by enhancing neovascularization and cell survival within the constructs.47,86 In the present review, significantly better in vitro cell survival, metabolic activity, and differentiation were observed in human-derived MSC and EC cocultures compared with MSC or EC monocultures. This is consistent with recent reports of the contribution of ECs to enhanced osteogenic differentiation of MSCs.92,100 Cotransplantation of human BMSCs or ASCs with EPC/ECs (with or without prior coculture) in rodent calvarial CSDs yielded significantly greater bone formation in two studies,41,45 while others42–44 observed no differences between cotransplantation and MSC-only groups. In one study, 47 human ASC monocultures demonstrated superior in vitro activity and in vivo bone formation than 1-week ASC-HUVEC cocultures on titanium mesh scaffolds. Therefore, the evidence for bone regeneration by cotransplantation of human MSCs and ECs is conflicting. Interestingly, one study, 101 excluded from the current review due to lack of quantitative outcomes, reported a codifferentiation strategy where human ASCs cultured in a combination of osteogenic and vasculogenic induction media acquired both phenotypes—positive for CD31 and osteopontin. These codifferentiated ASCs, when seeded on HA scaffolds and implanted into rat calvarial CSDs, resulted in superior (qualitative) bone and vessel formation than scaffold-only controls. 101
It is of relevance to consider the sources and types of vasculogenic cells used in a coculture/cotransplantation system. EPCs, under physiological conditions, are largely restricted to bone marrow and peripheral blood and consist of a heterogeneous population of cells.102,103 Recent evidence suggests that EPCs may also be isolated from umbilical cord blood. 104 ECs are an EPC subtype committed to EC differentiation. 102 In the included studies, relatively less variation in human EPC/EC sources was observed compared with osteogenic cell sources (Table 2). EPCs were derived from bone marrow, peripheral blood, or umbilical cord blood, while ECs were most often derived through endothelial differentiation of MSCs. The contribution of implanted human EPCs or ECs to vessel formation was confirmed by identification of human-specific endothelial markers in regenerating tissues.41,42,44,45 Vessel counting was generally performed on stained histological sections of the central region of tissue explants and therefore represents vasculogenesis in the central part of the construct. This finding is of clinical relevance since a limitation of traditional BTE constructs is the lack of vascularization within the central regions.16,22
Vascularization in BTE
Implanted EPC/ECs are reported to contribute to neovascularization through direct endothelial differentiation (vasculogenesis) and/or stimulation of the host regenerative response, for example, by release of trophic factors to recruit host EPC/ECs. 86 Recent evidence suggests that exogenous EPCs are also involved in homing of MSCs to defect sites, 105 while MSCs, in turn, may stimulate EPC differentiation through release of trophic factors. 106 Moreover, a fraction of MSCs exposed to endothelial factors reportedly differentiate into pericytes, which line and stabilize newly forming vessels.92,95 One study reported differentiation of human BMSCs into smooth muscle cells (SMCs) before coculture and cotransplantation with EPCs—the hypothesis was that SMCs would support EPC survival and stabilize the regenerating vessels, thus improving vascularization. 44 However, no significant additional benefit of cotransplantation versus MSC-SMC-only or EPC-only transplantation was observed in terms of bone and vessel regeneration. 44
Initiating prevascularization within BTE constructs before implantation is another emerging strategy. 107 One included study 37 reported the identification of nascent microvascular networks in vitro following 1-week dynamic coculture of rat BMSCs and ECs on copolymer scaffolds. Upon implantation into rat CSDs for 8 weeks, slightly higher amount of vessel formation was observed in the regenerating tissues of cocultures than MSC-only implants. 37 In context, a recent study 79 in a sheep model investigated bone regeneration in femoral defects implanted with prevascularized (MSC+EC) constructs dynamically cultured for 14 days in a perfusion bioreactor. Histological analysis of the prevascularized constructs at 16 weeks revealed microvessel-like structures containing red blood cells, suggesting a functional connection of primitive networks with host vasculature. 79 The ability of primitive vessels in prevascularized constructs to anastomose with host vasculature has previously been demonstrated.20,108–110
In the present review, one study 59 reported the use of a radiopaque contrast agent to quantify vessel regeneration within rat calvarial CSDs through micro-CT. At 4 weeks, infiltration of the contrast agent revealed vessel formation and/or infiltration (from the periphery) throughout the defect—no significant differences in vessel quantity (%) were observed between the central and peripheral regions. 59 Such novel imaging techniques, which can assist in tracking the in vivo behavior of primitive vessels in prevascularized constructs, should be utilized in future studies of vascularized BTE in orthotopic defect models.
The primary rationale for cell cotransplantation is that ECs can form primitive microvascular networks and improve cell survival in BTE constructs. 111 Although the presence of ECs may enhance vascularization in CSDs, transplantation of solely ECs (vs. solely MSCs) does not appear to enhance bone formation; indeed, the presence of an osteogenic cell population within the construct is reportedly necessary for osteogenesis. 112 In the present review, three studies42,43,45 included a control group of solely human EC transplantation and observed significantly lower bone formation compared with MSC-only or MSC+EC groups, while one study 44 reported no differences between groups. A similar trend was observed for vessel formation. However, in one study, 54 transplantation of rat ASC-derived ECs yielded significantly greater bone and vessel formation than MSC-only or MSC+EC transplantation. Therefore, the combination of MSCs with ECs, at least for human-derived cells, appears to be the most optimal strategy for vascularized BTE than any one cell type alone.
Alternative strategies
Alternative strategies to enhance vascularization in BTE constructs have been proposed, of which local delivery of exogenous osteogenic and/or angiogenic GFs has been extensively documented. 16 GFs are molecular signals that coordinate wound healing and regeneration. Local delivery of exogenous (recombinant) or autologous GFs can direct cellular behavior by initiating specific osteogenic cell signaling cascades and mobilizing host progenitor cells (MSC, EPC) to the regeneration site.113,114 Delivery of pooled (e.g., autologous platelet concentrates) or single GF (e.g., recombinant human bone morphogenetic protein-2 [rhBMP-2] or growth and differentiation factor-5 [rhGDF-5]) combined with biomaterial scaffolds has been shown to significantly enhance bone regeneration in clinical studies of contained or limited bone defects, for example, sinus augmentation, saddle defects, and extraction sockets.114,115 However, for larger defects, combined delivery of osteogenic and angiogenic GFs may be more beneficial. 116 VEGF plays a critical role in angiogenesis during bone formation and is an ideal candidate for combination GF delivery strategies. 14 Recent preclinical studies have reported conflicting results of bone and vessel regeneration, following implantation of scaffolds loaded with BMP-2 or VEGF either alone or in combination, in calvarial CSD models.117–119 In the present review, delivery of VEGF through β-TCP-PLGA scaffolds resulted in significantly lower %NBF than delivery of VEGFs with MSCs and/or EPCs in canine mandibular CSDs, 60 suggesting that GF delivery may compliment, but not substitute, the regenerative action of implanted cells. Recent studies, however, have shown that by using an appropriate delivery mode, it could be possible to lower the dose and delivery of GFs and minimize adverse reactions.120,121 Determination of the optimal dosage, delivery method, temporo-spatial release profile, and reliance on host cell responses are some of the reported limitations of current GF delivery strategies.16,22
Another proposed strategy is the implantation of genetically modified MSCs encoding for angiogenic and/or osteogenic GFs. 122 MSCs are modified ex vivo, most commonly through adenoviral transduction, to express favorable GFs (e.g., BMP-2, VEGF) or transcription factors (e.g., HIF-1α) and have been shown to enhance bone regeneration in craniofacial CSDs compared with unmodified MSCs.123–126 In the present review, one study 52 reported significantly greater bone and vessel formation in rat calvarial CSDs following coculture and cotransplantation of BMP-2-transduced MSCs and ECs compared with unmodified cocultures or unmodified MSCs after 5 weeks. However, additional sensitive in vitro procedures relating to GF gene transduction must be considered in these strategies.
Meta-analyses and heterogeneity
A random effects model was chosen for the present meta-analysis to account for the expected between-study variance. 127 Heterogeneity (I 2 ) was found to be very high within all categories in the meta-analyses. This could be due to biological factors related to the animals and/or methodological differences between the studies. Biological factors may include the animals' species, gender, age, immunological status, or the sources of MSCs and ECs used, and so on. In particular, sex and age of animals are reported to affect morphological, physiological, immunological, and behavioral parameters and thereby influence the outcomes of experiments. 128 Methodological differences in the study design (e.g., sample sizes, randomization), nature of interventions (e.g., use of autologous vs. allogeneic or human cells, osteogenic induction of cells before implantation), and outcome evaluation (e.g., methods of bone histomorphometry or micro-CT) may have also contributed to heterogeneity.
Although histomorphometry is considered the gold standard method for evaluation of 2D bone structure, micro-CT is emerging as a reliable alternative for assessing 3D microarchitecture with high resolution and accuracy in a fast and nondestructive manner. Several studies have reported high correlation between micro-CT and histomorphometry.129–131 However, to minimize heterogeneity in the present meta-analyses, results of micro-CT and histomorphometry studies were analyzed separately.
Quality of reporting
The reliability of results of meta-analyses directly depends on the quality of the primary studies. 127 As assessed by compliance with the ARRIVE guidelines and SYRCLE RoB tool, the included studies were judged to be of moderate reporting quality, but an unclear to high RoB. The ARRIVE guidelines aim to improve the reporting quality of animal studies and have been widely used for assessment of preclinical research in implant dentistry, 32 while the SYRCLE tool addresses particular aspects of bias that play a role in animal experimental studies. 34 Nevertheless, a clear need for more standardized reporting of animal studies was identified herein to allow reliable future reproduction and synthesis.
Clinical relevance
Although no clinical studies have reported direct coculture or cotransplantation of cells for bone regeneration in humans, one study 132 evaluated the efficacy of bone marrow aspirate containing populations of both hematopoietic (CD34+ HSC) and mesenchymal (CD44+, CD90+, CD105+ MSC) progenitor cells for mandibular reconstruction. Patients (n = 20 per group) received bone marrow grafts with either standard (54 ± 38 cells/mL) or high (1012 ± 752 cells/mL) concentrations of CD34+ HSCs and similar concentrations of MSCs (15 × 106 cells) together with rhBMP-2 in an allograft scaffold. Significantly higher %NBF was observed in patients receiving higher concentrations of CD34+ HSCs (67% ± 13% vs. 36% ± 10%) after 6 months, suggesting an important and complimentary role of HSC to MSC in alveolar bone regeneration and the benefits of implanting heterogeneous cell populations at regeneration sites.132,133
Due to the large inherent variation in animal studies, preclinical meta-analyses aim to disclose the direction rather than size of the effect of an intervention.127,134 Fundamental limitations of animal models that must be considered when interpreting the findings are (1) the underestimation of clinical variation, with regard to both local (defect size, morphology, mass transfer, etc.) and systemic (age, comorbidities, etc.) aspects; and (2) the overestimation of clinical performance since the CSDs utilized are uniform surgically created defects with sound surrounding tissues and generally uncompromised blood supply and most often involve healthy animals, which is often not the case in clinical scenarios. 75 Moreover, the dimensional differences between experimental defects in small animals and the clinical situation must be considered, especially with regard to vasculogenesis. Although it may be inferred that bone and vessel regeneration observed within calvarial CSDs in the included studies was largely due to the direct and/or indirect regenerative effects of exogenous cells functioning in a truly demanding environment (due to the highly cortical structure [low bone marrow] and poor local vascularity [absence of a primary nutrient artery] of the calvarium28,135,136), the numerical values or magnitude of response from the present meta-analysis cannot be directly translated to the human situation. Thus, the ∼8% additional NBF observed in defects augmented with cotransplanted cells would not translate to 8% (or x-times 8%) more bone if the same procedure was performed in humans. 27 Moreover, considering the magnitude of estimates of treatment effect, the clinical relevance (benefit) of cotransplantation procedures for oral and maxillofacial indications may be questioned.
Conclusions
The reviewed preclinical in vivo evidence can be summarized as follows:
(1) Based on the results of small animal (rodent) models, cotransplantation of osteogenic (MSCs or OBs) and vasculogenic cells (EPCs or ECs) seeded on biomaterial scaffolds results in greater radiographic bone regeneration than transplantation of solely MSC-/OB-seeded constructs. (2) Coculture of MSCs and EPC/ECs under specific conditions in vitro can enhance their regenerative potential when cotransplanted in vivo. (3) MSCs and ECs from human sources demonstrate synergistic activity when cocultured in vitro. However, the evidence for bone and vessel regeneration following in vivo cotransplantation of human cells is inconclusive. (4) The magnitude of estimates of treatment effect was relatively small and thus the clinical relevance (benefit) of cotransplantation procedures for oral and maxillofacial indications may be questioned. (5) The results should be interpreted with caution due to large heterogeneity between studies as a result of biological and methodological factors. (6) There is a need for more standardized reporting of well-designed and adequately powered animal studies to allow reliable future reproduction and synthesis.
Footnotes
Acknowledgments
The authors would like to thank all the corresponding authors of the primary studies who provided additional information for the review.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.