
Abstract
Mesenchymal stem cells (MSCs) are considered to hold great therapeutic value for cell-based therapy and for tissue regeneration in particular. Recent evidence indicates that the main underlying mechanism for MSCs' beneficial effects in tissue regeneration is based on their capability to produce a large variety of bioactive trophic factors that stimulate neighboring parenchymal cells to start repairing damaged tissues. These new findings could potentially replace the classical paradigm of MSC differentiation and cell replacement. These bioactive factors have diverse actions like modulating the local immune system, enhancing angiogenesis, preventing cell apoptosis, and stimulating survival, proliferation, and differentiation of resident tissue specific cells. Therefore, MSCs are referred to as conductors of tissue repair and regeneration by secreting trophic mediators. In this review article, we have summarized the studies that focused on the trophic effects of MSC within the context of tissue regeneration. We will also highlight the various underlying mechanisms used by MSCs to act as trophic mediators. Besides the secretion of growth factors, we discuss two additional mechanisms that are likely to mediate MSC's beneficial effects in tissue regeneration, namely the production of extracellular vesicles and the formation of membrane nanotubes, which can both connect different cells and transfer a variety of trophic factors varying from proteins to mRNAs and miRNAs. Furthermore, we postulate that apoptosis of the MSCs is an integral part of the trophic effect during tissue repair.
Introduction
T
In the tissue engineering field, researchers have sought ways to exploit the multilineage differentiation potential of these culture expanded MSCs to regenerate lost or worn out tissue. For example, a working hypothesis is that MSCs would generate de novo tissue themselves by incorporating specific biological cues into scaffolds that combined with the cells would stimulate their differentiation into the desired cell type (Fig. 1A). This strategy is currently explored in preclinical studies.11–13 In another strategy, MSCs are injected either at the site of the injured tissue or in the blood stream. It is believed that the MSCs would subsequently home to the diseased tissue and induce tissue repair. Indeed this concept is under investigation in a large number of preclinical trials for treating a wide range of diseases.14–16 While beneficial effects of this approach have been reported, there is much uncertainty with respect to the mechanism of action. In fact, only a small fraction of the injected cells are actually homing to damaged tissues and even less cells can be found back in the tissue in long-term follow-up of days to weeks.17–21 After transplantation into the heart, the exogenous MSC showed poor survival and do not persist in the infarcted area. 22 Other investigators have even failed to detect permanent engraftment of transplanted MSCs in infarcted hearts. 23 MSC has been found to home preferentially to the ischemic boundary in stroke studies. 24 However, it has been shown that cells disappeared progressively and were no longer detected 4 weeks after transplantation. 25 Similar findings were reported in another study in human adipose tissue-derived MSCs in a mouse model of hind limb ischemia. Indeed the injected cells improved neovascularization, but the incorporation of MSCs into the vascular structures resulted quite low (less than 1%), indicating that other effects, rather than direct differentiation of MSCs in neovasculature, likely accounted for the observed beneficial effects. 26 All this evidence suggests that in many tissue engineering approaches the contribution of MSC to support the repair process by directly differentiating in de novo repair tissue is fairly limited due to poor engraftment and survival of cells. 27 Since in many of these studies beneficial effects have been reported this raises the question how the injected cells exert their effect.

Mechanisms through which MSCs mediate tissue repair.
Besides their multilineage differentiation capacity, MSCs have shown to be capable of reducing inflammation by modulating the immune system,28–35 promoting angiogenesis,36–40 and inducing cell migration, proliferation,41–44 differentiation, and extracellular matrix (ECM) formation39,45,46(Table 1). Therefore, it has been proposed that the functional benefits observed after MSC transplantation in experimental models of tissue regeneration might be related to the trophic effects of MSCs (Fig. 1B).47–49 This specific role of MSCs is the main focus of this review. In particular, we will discuss the possible mechanisms behind this effect.
MSCs Act as Trophic Mediators in Tissue Regeneration
MSCs exhibit the capacity to migrate to injured sites and can, therefore, contribute to tissue repair. 50 The trophic effect of MSCs in tissue repair was first proposed by Arnold Caplan. 51 There is a dynamic regulation and interplay of stem cell derived cytokines that influence tissue survival, repair, and regeneration, including the activation of resident and circulating stem cells. 27 Currently, researchers are increasingly integrating the trophic roles of MSCs as a prominent feature in tissue repair strategies. 52 One engrafted MSC has the potential to modulate the activity of many surrounding cells through intercellular communication. This hypothesis is supported by the in vitro observation that MSC-conditioned medium (CM) improves tissue repair.27,53 MSC-CM acts as a chemoattractant for tissue-specialized cells. 54 Analyses of MSC-CM indicate that MSCs secrete many known mediators of tissue repair, including growth factors, cytokines, and chemokines. 53 This evidence suggests that the therapeutic effect of MSCs may largely depend on their capacity to secrete soluble factors that promote several key biological activities.
MSCs in osteoarthritis and other cartilage defect repair
Partial replacement of chondrocytes with stem cells in autologous chondrocyte implantation has been proposed as an effective strategy to limit or even omit the in vitro expansion phase of chondrocytes. 55 Indeed, in vitro studies have shown that cartilage matrix deposition and chondrogenic gene expression in MSCs could be improved by coculture of MSCs and chondrocytes.56,57 Based on these studies, it was assumed that the beneficial effects of coculturing MSCs and chondrocytes in cartilage matrix formation were largely due to the differentiation of MSCs into chondrocytes. However, using cell tracking experiments it was later shown that DiI-labeled allogeneic MSCs in cartilage defects were present after 1 week, but not after 4 or 12 weeks postimplantation, while the cartilage repair progress lasted for at least three months. 58 This implies that the transplantation of allogeneic MSCs could only have acted at the beginning of the repair process. Importantly, this experiment indicated that MSCs did not contribute significantly to wound healing by differentiating into the newly formed tissue.58–60
In a study performed by our group 61 human MSCs (hMSCs) were used in pellet coculture with human primary chondrocytes (hPCs) or bovine primary chondrocytes (bPCs), and the role of the individual cell populations in matrix formation was studied. This revealed that these coculture systems outperformed monoculture systems of either hPCs or hMSCs in terms of cartilage formation. Interestingly, the increase in matrix deposition was mainly achieved by increasing the matrix production of hPCs. Furthermore, we observed a significant decrease in MSC overtime, most probably caused by apoptosis, which may play an important role in the cartilage matrix formation of pellet coculture system.62,63 The beneficial effects of the pellet coculture model were largely due to the stimulation of proliferation and the matrix formation of chondrocytes induced by a trophic effect of the MSCs. This trophic role of MSCs in cartilage formation is furthermore supported by independent studies performed by others.64–66 These studies point to a dominant role of the MSCs in stimulating resident or coimplanted chondrocytes to initiate a regenerative response. One can interpret the role of the MSC in these systems as the initiator or even as the conductor of the repair process by activating cell and tissue autonomous repair mechanisms, such as the induction of chondrocyte proliferation and matrix production. Once activated, the cartilage regeneration becomes self-reliant and chondrocyte driven and is largely independent of MSC-based stimulation. Moreover, the trophic effect of MSCs is independent of culture conditions and the origin of the MSCs.66,67 These studies suggested that the supportive trophic role in tissue repair and regeneration might not be limited to MSCs, but could also be displayed by other stem cell populations or even non-stem cells such as dermal fibroblasts. 56 These findings starting from observations in vitro have been confirmed in animal studies and recently also in a clinical trial.68,69
MSCs in renal regeneration
The role of MSCs in the recovery of acute kidney injury (AKI) has been extensively studied. It has been reported that MSC administration effectively counteracted the detrimental effects of experimental AKI70–72 and induced functional improvements in chronic kidney disease (CKD). 73 Initially, it was thought that MSCs would home to the kidney to replace damaged renal cells. However, after initial accumulation of systemically administered MSCs in the injured kidney, only a few of these cells permanently engraft within the tubules and without obvious evidence of cell differentiation into tubular cells.70–72 In a rat AKI model, the iron-labeled MSCs were predominantly located in the glomerular capillaries, while kidney tubular cells showed no iron labeling, indicating the absence of transdifferentiation into tubular cells. 74 It has been suggested that MSCs do not replace renal tubular cells, but mitigate the damage by providing paracrine support for repair. MSCs achieve this by releasing growth factors that modulate the immune response and, subsequently, stimulate tissue repair.75,76 The kidneys of MSC-treated rats revealed a decrease in gene expression of pro-inflammatory cytokines and an increase in several growth factors with mitogenic, pro-survival, and antiapoptotic effects. 77 In particular, insulin-like growth factor (IGF-1) and vascular endothelial growth factor (VEGF) were suggested as critical mediators.42,78 In an in vivo model of lung injury, a decline in the levels of tumor necrosis factor (TNF), interleukin (IL)-6, and interferon-gamma (IFN-γ) was also detected in the group pretreated with MSCs. 79 This hypothesis is supported by a mouse model of tubular injury showing that injection of CM from murine MSCs resulted in a significant decrease of tubular cell apoptosis, increased survival, and renal function improvement. 80 Therefore, it can be concluded that the regenerative response of MSCs in this model depends on secreted bioactive factors.
MSCs in myocardial repair and regeneration
Injection of MSCs has been demonstrated to have therapeutic effects after myocardial infarction. Similar to the mechanism of MSCs promoting kidney repair, current data suggest that the direct differentiation of allogeneic MSCs into cardiomyocytes is very limited. Instead, the MSCs secrete factors and regulate the function of cardiomyocytes and immature cells during repair.81–84 In support of this paracrine hypothesis, many studies have observed that MSCs secrete cytokines, chemokines, and growth factors that could potentially repair damaged cardiac tissue. VEGF, transforming growth factor beta 1 (TGF-β1), fibroblast growth factor 2 (FGF-2), hepatocyte growth factor (HGF), and granulocyte colony stimulating factor (G-CSF) have all been identified as key factors released by MSCs. 85 These factors have been demonstrated to exert beneficial effects on the heart, including neovascularization, attenuation of ventricular remodeling, and increased angiogenesis. 39 Anti-inflammatory action is also exerted by MSC in a murine acute myocarditis model. 86 Furthermore, MSC-CM can reduce apoptosis and necrosis of cardiomyocytes exposed to low oxygen tension. 87 The injection of MSC-CM into animal myocardial infarction models limited infarct size, reduced cardiomyocyte apoptosis and ventricular remodeling, and resulted in improved cardiac function compared with controls.49,88,89 Meanwhile, extra cardiac administration of MSCs provided clear evidence that cardiac repair can be achieved through MSC's trophic actions and did not rely on their presence in the infarcted myocardium. 90 It should be emphasized that many of these positive effects of MSCs in AKI and myocardial infarction were found in animal models. Replication of these beneficial effects in human patients is the subject of many clinical trials and is subject of an intense scientific debate.
MSCs in wound healing and vascularization
Neovascularization is another important biological process positively influenced by stem cells in a paracrine manner. Coculture of endothelial cells and MSCs can simulate and stabilize endothelial microvascular networks and promote cell functions. 91 A study with coapplication of MSCs and endothelial cells resulted in improved angiogenesis which was largely due to the secretion of angiogenic cytokines by the MSCs stimulating the endothelial cells. 92 Another mechanism by which MSCs contribute to wound healing is by reducing inflammation and by inducing migration and proliferation of resident tissue-specific cells. 27 Many of these effects can be replicated using MSC-CM both in vitro 93 and in vivo. 53 Indeed, the CM acted as a chemoattractant recruiting macrophages and endothelial cells to the wound. The MSC-CM has a similar effect as MSCs on a wound both accelerating epithelialization.53,94,95 Studies have indicated that many cell types in the wound area, including epithelial cells, endothelial cells, keratinocytes, and fibroblasts, are responsive to factors secreted by MSCs. 96 The trophic function of MSCs in the wound healing environment has been further elucidated in animal models. In a diabetic rat model of ulcerating wounds, researchers evaluated the function of MSCs in the context of improved collagen metabolism. 97 Overall, these data suggest that MSCs released soluble factors that stimulated proliferation and migration of the predominant cell types in the wound in therapeutically relevant concentrations.
MSCs in neurological disorders
MSCs have been reported to provide a potential therapeutic benefit in the treatment of neurological disease and injury. 98 MSCs are able to exert neurotrophic effects by releasing a variety of molecules that directly or indirectly promote endogenous repair. Such molecules may include neurotrophic growth factors, chemokines, cytokines, and ECM proteins. 99 Increased levels of IGF-1, as well as VEGF, bFGF, and epidermal growth factor (EGF), were observed in the brain of treated rats, compared with controls. 48 Paracrine effects of such factors involve direct neurotrophic and/or neuroprotective activity on resident progenitor cells, hence inducing neurogenesis, oligodendrogenesis, 100 neurite outgrowth,101,102 angiogenesis, 103 or antiapoptotic effects 48 on neurons and glia cells. Furthermore, MSCs have been shown to improve gain-of-functions in brain stroked rats without differentiating into a neuronal phenotype. 104 It is assumed that the MSCs were able to achieve this by secreting a cocktail of factors into the neural niche microenvironment that mediated neural repair and protection.
Mechanisms Behind the Trophic Effects
In each of these disease models discussed above, the MSCs secreted factors that stimulated neighboring cells. It has been proposed that the interaction of stem cells with their microenvironment, also named niche, has a critical role in defining the phenotype of the stem cell but also determines their secretory profile. 105 This has suggested that the selection in the production of the paracrine factors produced by the MSCs is, in part, orchestrated by the injured tissue. Indeed, MSCs appear to enhance the regenerative potential of multiple tissue types as a result of various mechanisms that become activated when exposed to the biochemical factors that are characteristic of an injured environment in vivo. 106
In a living tissue, cells constantly interact with each other and their ECM. This interaction influences the proliferation and differentiation of cells within the tissue and organ.107,108 Cell–cell communication is required to guarantee effective coordination among different cell types within tissues. Classical means of cell communication (Fig. 2) are represented by cell junctions, cell adhesion molecules, and signaling through secreted and soluble factors, which all connect neighboring cells acting in a paracrine or even endocrine manner.109–111

Classical forms of cellular communication.
More recently, communication by extracellular vesicles (EVs) and membrane nanotubes (MNTs) have been identified as possible mechanisms. Furthermore, we postulate that apoptosis is an integral part of the trophic effect of MSCs in tissue repair. It seems likely that each of these mechanisms can contribute to the trophic role of the MSC in tissue regeneration. The extent to which each of these mechanisms can contribute to the therapeutic effect is likely dependent on the context such as tissue type, type of tissue damage, and inflammation degree. Recent articles summarized the mechanisms of the more classical ways of cell communication.109,111 In this review, we will emphasize on the latter three mechanisms.
Extracellular vesicles
EVs are cytosol fragments with a spherical morphology, surrounded by a membrane composed of a lipid bilayer and hydrophilic proteins, similar to the cell plasma membrane (Fig. 3). 112 Such vesicles are released from most cells and can be isolated from most body fluids such as serum, plasma, urine, and cerebrospinal fluid.113–115 EVs are a heterogeneous group of vesicles and include exosomes, microvesicles, and apoptotic bodies. It has proven to be a challenge to experimentally distinguish between these vesicles due to their overlapping biophysical characteristics and lack of discriminating markers.116,117 Therefore, the term EVs will be used in this review to refer to a mixed population of vesicles.
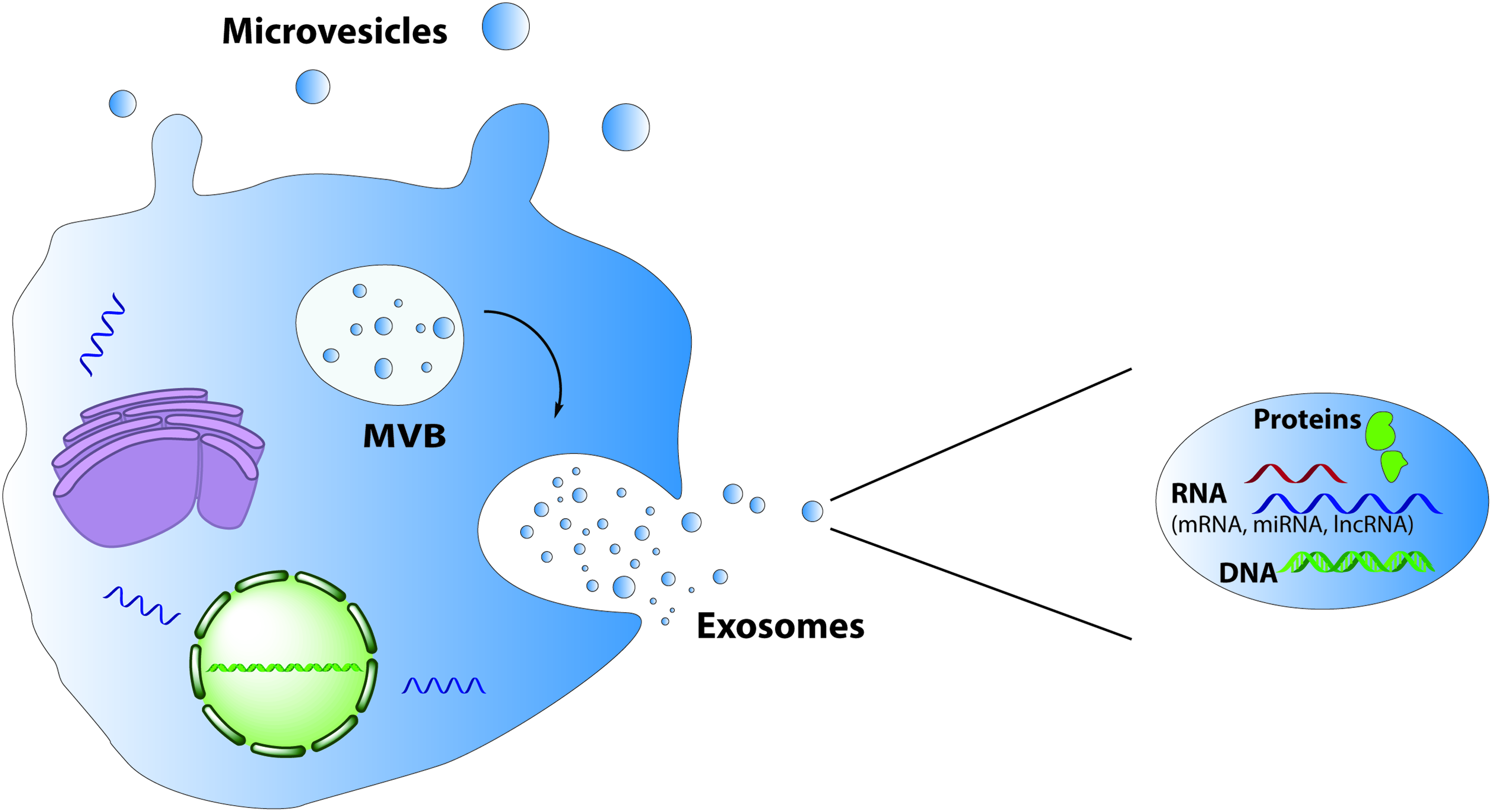
Mechanisms involved in the formation of MVs and exosomes. Exosomes are formed from MVBs and are released by fusion of MVBs with the plasma membrane, whereas MVs bud directly from the plasma membrane. The EVs contain a variety of proteins, RNA, and DNA species. EV, extracellular vesicle; MV, microvesicle; MVB, multivesicular body. Color images available online at www.liebertpub.com/teb
Formation of EVs
Exosomes are 30–100 nm in size and are secreted when multivesicular bodies, a late endosomal compartment, fuse with the plasma membrane.118,119 Exosome production is generally regarded as a constitutive membrane vesicle pathway, although it can increase as a result of stimulation with, for example, Ca2+ ionophore. 112 Microvesicles, also termed shedding vesicles, tend to be 100 nm to 1 μm in diameter and are directly derived from the cell membrane of activated cells through the disruption of the cortical cytoskeleton. These vesicles shed into the extracellular space in a calcium flux- and calpain-dependent manner. 120 Apoptotic bodies are larger than 1 μm and are derived from dying cells. They result from budding of the plasma membrane and contain cytoplasm with organelles. 121
These apoptotic bodies are a special class in the EVs. In tissue regeneration studies, cell death was considered as a consequence of injury, lack of oxygen and not as a regenerative factor, until recently. Researchers proposed the concept of “altruistic cell suicide” based on their observation that dying cells could induce proliferation of their neighboring cells. 122 The apoptotic cells release a variety of signals, including apoptotic bodies, which induce cellular responses over short and/or long-range distances. During apoptosis, apoptotic bodies containing biological information are transferred from apoptotic to nonapoptotic cells. Indeed, endothelial cell-derived apoptotic bodies convey paracrine signals to recipient vascular cells that trigger the production of CXCL12. 123 These data indicate that the apoptotic bodies likely play an important role in paracrine signaling supporting tissue repair and regeneration. Thus the disappearance over time of MSCs from the injured tissue could reflect the death of MSCs. The extent to which MSC's cell death contributes to tissue regeneration, as well as the underlying mechanism of this “altruistic cell death” deserves further study. As mentioned before, also in the pellet coculture model of MSCs and primary chondrocytes an association was found between MSC disappearance from the pellets likely by apoptosis and the stimulation of chondrocyte proliferation and matrix formation. It is presently unclear whether the cell death of MSCs in these pellet cocultures with primary chondrocytes contributes to the improved cartilage formation in this model.
EVs biological activities
Cell communication by means of EVs is considered to be a universal way for cells to interact with each other and influence the behavior of other neighboring cells by exchanging material and information. Indeed, EVs can stimulate target cells and induce alterations in the phenotype and behavior of recipient cells.124,125 EVs influence the fate of target cells in numerous ways. First, they may directly stimulate the cells by interaction with membrane bound receptors or ligands. 126 Indeed, EVs express surface molecules and several types of receptors, including transforming growth factor β (TGFβ), CD95 L, MHC class I/II molecules, and the CCR5 chemokine receptor.126,127 After ligand interaction, they may then modulate the functional target cell by delivering intracellular proteins.128,129 Second, EVs can transfer receptors, bioactive lipids, or nucleic acids between cells after fusion with target cell membrane. Of particular interest is the involvement of EV in the horizontal transfer of genetic information. Subsets of mRNAs, miRNAs, and long noncoding RNAs can be transferred through EVs to recipient cells, inducing functional and phenotypic changes.130,131 In particular, the horizontal transfer of miRNAs has been proposed as a new form of intercellular communication, representing a means by which donor cells can regulate the gene expression of recipient cells. 132 EVs extend the trophic repertoire of MSCs from the classical secreted growth factors to now include as well mRNAs, miRNAs, lipids, and membrane bound proteins.
Stem cells and differentiated cells communicate with each other to regulate the self-renewal and differentiation processes related to tissue repair. 133 In this process, stem cells and differentiated cells may establish bidirectional communication during the reparative process. EVs, by transferring selected patterns of proteins, lipids, mRNAs, and miRNAs to recipient cells, may be considered as potent paracrine mediators of signaling between stem cells and differentiated cells. 110 In the first scenario, EVs released from injured tissue may help to reprogram the phenotype of stem cells to acquire tissue-specific features. The other way around, EVs derived from stem cells may induce cell cycle reentry of cells that survived an injury allowing tissue regeneration. 110 In this context, the stem cell-derived EVs could participate in the repair process 105 by provoking genetic and epigenetic changes in target cells thereby activating critical cell processes for tissue regeneration, for example, by inducing cell proliferation, differentiation, and ECM production. Meanwhile, EVs released from damaged tissues may organize adult resident stem cells into a reparative program. 134 With the progress of tissue repair, the demands of the regenerating tissue will change and may request a different type of support from the MSC. Paracrine signaling, for example, by EVs back and forth between tissue specific cells and the stem cells may keep the trophic role of the MSCs in pace with the regenerative process. EVs may exert these functions next to classical ways of communication by means of secreted growth factors. In fact, the beneficial effect of MSC-CM as reported in many studies cannot only be attributed to the presence of secreted growth factors but could have been mediated at least, in part, by the presence of EVs which are in most protocols also present in the CM. It would be of great interest to repeat the CM experiments with medium deprived from EVs to determine the contribution of either secreted growth factors or the EVs to the beneficial effects in tissue repair.
EVs contain diverse species of RNAs that reflect the functional state of the cell of origin and could represent an important therapeutic tool, since these RNA subsets impact directly or indirectly protein translation when taken up by the target cells. 135 Therefore, several studies have evaluated whether stem cell-derived EVs perform similar to MSCs in tissue regeneration in vivo.136–138 The use of MSC derived EVs rather than the cells themselves may avoid possible long-term mal-differentiation of engrafted cells and attenuate many of the safety concerns related to the use of living cells.
Role of EVs in tissue regeneration
Proteomics analysis of MSC-derived EVs has shown that they contain the factors influencing angiogenesis such as FGF, VEGF, HGF, EGF, and IL-8.139,140 Thus MSCs release not only growth factors directly into medium but also as an integral part of EVs. It is at present unclear whether growth factors embedded in an EV have distinct biological activity compared to the same growth factors directly released into the extracellular space. This requires more research. Nevertheless it is clear that release of growth factors in EVs represents another way for cellular communication and transportation in the extracellular space. 141
EVs have been shown that they are the mediators of the cardioprotective effects of MSCs and that they diminish the size of the infarct in an ischemic mouse model. 142 A similar model showed that MSC-derived EVs enhanced blood flow recovery, reduced the infract size, and preserved cardiac systolic and diastolic performance by promoting angiogenesis. 143 Currently it is hypothesized that the cardioprotective effects of EVs are resulting from a combination of improved angiogenesis, antiapoptotic effects, anti-inflammatory effects, and anticardiac remodeling factors showing a clear overlap with the proposed actions of MSC-CM. Recently, researchers showed that CXCR4 overexpressing MSCs produced EVs with increased concentrations of VEGF and IGF-1α. They loaded MSCs with EVs, which were produced in CXCR4 overexpressing MSCs, and proved that these exosomes were delivered in vivo upon implantation of a cell sheet, leading to increased angiogenesis, reduced infarct size, and improved cardiac remodeling. 144
Bruno et al. wrote an extensive review on the use of MSC-derived EVs on renal regeneration. 119 MSC-derived EVs were first tested for their renal regenerative potential in a model of AKI. 134 A meta-analysis on the studies using MSC-derived EVs in AKI concluded that MSC-derived EVs have a more profound therapeutic potential than CM and that early administration of EVs in AKI has the most effect. 145 Besides AKI, MSC-derived EVs have been tested in chronic diabetic nephropathy, where they were found to be as effective as CM. 146 The exosomes were found to suppress the invasion of bone marrow-derived cells (the source of inflammatory cytokines) by downregulating their adhesion molecules (ICAM-1). This resulted in a decrease in the expression of TNF-α. Besides this, activation of renal inflammatory pathways through p38-MAPK was reversed. TGF-β1 expression was downregulated, and tight junction protein expression (e.g., ZO-1) was persevered. Consequently, TGF-β mediated epithelial-to-mesenchymal transition was inhibited.
Recent studies compared the effect of injected MSCs and MSC-derived EVs on the neural functional recovery after ischemia or apoplexy.137,147 Both studies found equally positive therapeutic effects of MSCs and EVs. Improvement of neurological impairment, neuroprotection, and neurogenesis and angiogenesis were equally stimulated by the presence of either MSCs or EVs. Although the composition of the EVs was not within the scope of these studies, other research has shown that the neurotrophic effect of injected MSCs results from a variety of molecules that directly or indirectly promote endogenous repair (reviewed in chapter 2.5). Hofer and Tuan reviewed the role of MSC secreted trophic factors in never repair. 148 Neurotrophic brain-derived neurotrophic factor, nerve growth factor, glial cell line-derived neurotrophic factor, ciliary neurotrophic factor, as well as angiogenic VEGF and angiopoietin-1, work as neuroprotective factors. It is assumed that the MSCs are able to achieve their therapeutic effects by secreting a cocktail of these factors in EVs into the neural niche microenvironment that mediates neural repair and protection. However, to date, the molecular definition of their cargo remains unknown. Also these studies suggest a large overlap in the activity of MSC-CM and MSC-derived EVs, which may suggest that EVs rather than secreted growth factors and cytokines play a more dominant role in the mechanism of action by which MSCs induce tissue regeneration.
Based on the anti-inflammatory, antiapoptotic, antifibrotic, and pro-regenerative properties of MSC-derived EVs, it was long speculated that EVs might exert a positive effect on osteoarthritis (OA) or rheumatoid diseases.149,150 Zhang et al. showed that the addition of MSC-derived exosomes led to complete restoration of cartilage and subchondral bone. Hyaline cartilage with a good surface regularity and complete bonding to the adjacent cartilage was formed after 12 weeks in 5 out of 6 rats. 150
Based on all examples above EVs can have numerous positive effects on tissue regeneration due to their antiapoptotic, angiogenesis promoting, and pro-proliferative effects. Despite all these positive results many of the animal models used do not resemble a realistic pathological situation. Many of the models inject the EVs directly after injury, to prevent development of tissue damage, even when the disease is chronic and progressive like OA.119,143,144,150 There are studies where the administration of EVs is delayed, but these are the minority. In the studies for CKD there is one article using EVs to cure already established damage. 151 In this study they compared EVs and CM and found that EVs do not have a curative effect. One other article describes the regenerative effect of EVs in established chronic diabetic nephropathy. 146 The exosomes were found to suppress the invasion of bone marrow-derived cells by downregulating their adhesion molecules. Clearly more work is needed to find whether MSCs, MSC-CM, or MSC-derived EVs can play a role in the reversion or even regeneration of established disease and whether there are specific windows for opportunities in which treatment is most beneficial.
Membrane nanotubes
Rustom et al. 152 were the first to describe an as yet unknown type of intercellular connection. They noticed some straight lines connecting two adjacent cells that were not in direct cell–cell contact. With regards to dimensions and morphology, these structures were termed MNTs or tunneling nanotubes (TNTs). MNTs are long thin F-actin-based membranous channels connecting cells (Fig. 4). These structures were observed between many eukaryotic cells,153–156 suggesting that these connections represent common mechanism of functional intercellular connectivity and cell-to-cell communication. MNTs are widely considered a neotype of cell communication through its ability to transport many components and signals from one cell to another. 157
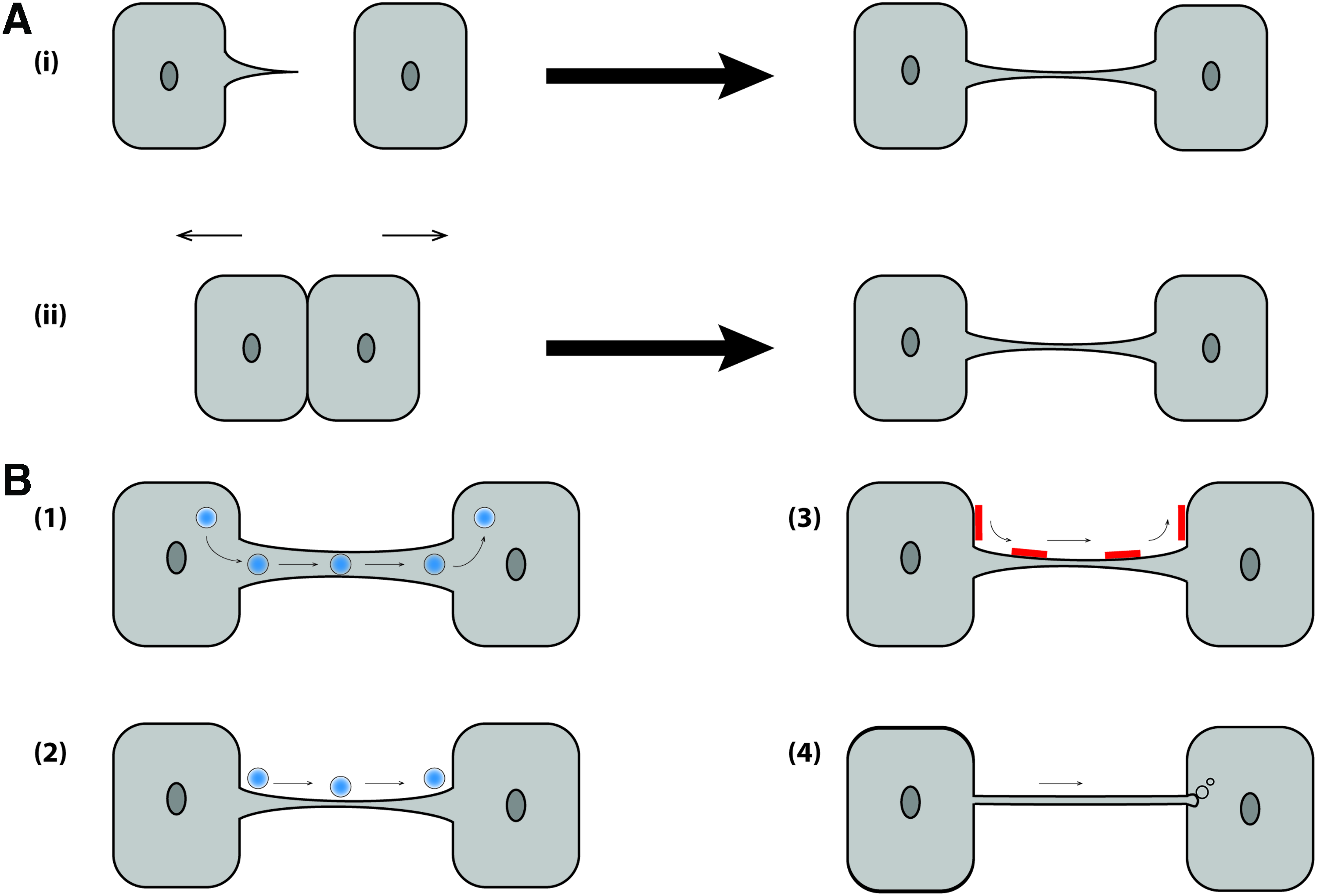
Mechanisms of MNT formation and potential functions.
Structural characteristics of MNTs
Electron transmission and scanning microscopes were used to reveal the surface and formation of MNTs. Meanwhile, fluorescence studies can give information about the structure of MNTs, as well as their function in intercellular transfer.158,159 To study the latter process, dyes such as DiD, DiO, fluorescent proteins like GFP, and selected cell compartment markers are widely used. The diameter of MNTs ranges from 50 to 200 nm and some are as long as the diameter of a few cells. 152 Findings suggest that the thickness and length of MNTs might be related to the components exchanged through the tunnels, as well as their quantity. 160
Two types of MNTs have been distinguished, open ended and closed ended. For open-ended MNTs, intercellular cargo or larger organelles such as mitochondria or vesicles may be transported from the MNTs to the target cell.160,161 For the closed-ended structures membranous cargo can enter into the target cell as a result of endocytic forces. 162 Their functions could apply to various specific topologies, depending on the types of signals.
Formation of MNTs
Until now, two mechanisms of MNT formation are known162–164 (Fig. 4A). In the first mechanism, linkages form after a cell extends a de novo filopodia-like bridge that is then bound and tightly anchored to a neighboring cell. Filopodia are dynamic exploratory and sensory organelles that can direct cell migration toward specific sources, including nearby cells. These structures can be maintained to form a long-lived stable filopodial bridge, or alternatively, function as structural intermediates to more complex cell–cell interfaces. 165 The second mechanism of MNT formation is connected with prior cell–cell contact, after which the cells are separated and a nanotube is formed between them. These mechanisms may vary per cell type. Indeed, most immobile cell types, such as neuronal-like PC12, form tubes by the outgrowth of filopodia. 166 Mobile cells, such as kidney cells and T cells, which easily connect with other cells, form tubes by the second method.156,167 However, it is important to note that the two mechanisms could occur in the same cell type. Despite the two possible mechanisms of formation, it is accepted that actin polymerization is required for MNT formation. This specific feature of MNTs helps to distinguish them from other similar structures such as membrane tubules formed by neutrophils. 168
Little has been done to investigate the signaling pathways involved in MNT formation. Since actin polymerization is required, it has been proposed that some proteins involved in actin polymerization, such as Cdc42, will also be important for nanotube formation. Indeed, Cdc42, Rac1, ezrin, and N-WASP are all present in MNTs, but the requirement of these factors in MNT formation has so far not been assessed. 169 Using expression of dominant negative constructs in HeLa cells, inhibition of Cdc42 resulted in decreased MNT formation, but Rac1 inhibition showed no effect. 170 It suggests that the signaling requirements for MNT formation may be different depending on the mechanisms of formation.
Biological significance of MNTs
Many studies have investigated the biological function of this recently discovered structure that connects two distant cells (Fig. 4B). A common function of MNTs is the propagation of an electrical signal. 167 Organelles and protein transport through MNTs also seem to play an important role especially when cells are damaged or undergo premature senescence.171–173 Ca2+ and intercellular vesicles could also be transported through MNTs between cells.161,174,175 Transfer of miRNA through MNTs has been described in cancer cells, but has to be investigated if this function is also present in immune cells or in MSCs. 176 MNTs were found to transport porous silicon microparticles, so that they may be involved in the dispersion of therapeutic agents at the site of diseased tissue. 177 MNTs have also been suggested to provide cell-contact dependent communication over long distances. 158 Recently, MNTs have been observed between MSCs and vascular smooth muscle cells (VSMCs), which promoted the proliferation but not differentiation of MSCs by transfer of mitochondria from VSMCs into MSCs, 178 as well as between MSCs and osteoclast precursors, which was found to be essential for osteoclastogenesis. 179 Overall, MNTs are novel candidates to explain how direct cell-to-cell communication occurs and play an important role in many (patho)physiological processes by transporting molecular signals and cell organelles from one cell to another. 180 We propose that communication through nanotubes could be an additional mechanism by which MSCs exert their positive effect in tissue regeneration.
Conclusions
The beneficial function of transplanted MSCs has already been proven in many therapeutic domains. Recently, the trophic effect of MSCs is being considered as the most important player in the observed reparative effects of these cells. In the case of severe tissue damage, MSCs can be attracted to the site where they secrete a broad repertoire of trophic factors that function to assist the repair and regeneration process. 51 The trophic hypothesis of MSCs action changed the perspective of the therapeutic use of MSCs in regenerative medicine. 181 The mechanisms governing this trophic activity are quite distinct from those used in tissue engineering functional substitutes for replacement of damaged or lost tissue. There is strong evidence showing that the effect is at least partly mediated by secretion of growth factors, cytokines, and other secreted proteins affecting resident cells stimulating tissue repair, but it is likely that more mechanisms exist. In this review, we have discussed two additional mechanisms that could be used by the MSCs in support of their trophic role, EVs and MNTs, which can connect different cells and deliver their trophic factors. These structures are defined as important forms of intercellular communication and are involved in many physiological processes. As they may influence the behavior of recipient cells by delivering their bioactive cargo, it may be possible to exploit this effect in tissue regeneration and repair. The extent to which these additional mechanisms contribute to the trophic effect of MSCs in tissue repair and regeneration deserves further study. Indeed gaining a solid understanding of these mechanisms may lead to development of new strategies and/or therapeutic tools for tissue regeneration.
Footnotes
Acknowledgments
This work was supported by a long-term grant of the Dutch Arthritis foundation and Innovative Research Incentives Scheme Veni #14328 of the Netherlands Organization for Scientific Research (NWO) and by the European Fund for Regional Development.
Disclosure Statement
No competing financial interests exist.