
Abstract
Biomaterial cues can act as potent regulators of cell niche and microenvironment. Epigenetic regulation plays an important role in cell functions, including proliferation, differentiation, and reprogramming. It is now well appreciated that biomaterials can alter epigenetic states of cells. In this study, we systematically reviewed the underlying epigenetic mechanisms of how different biomaterial cues, including material chemistry, topography, elasticity, and mechanical stimulus, influence cell functions, such as nuclear deformation, cell proliferation, differentiation, and reprogramming, to summarize the differences and similarities among each biomaterial cues and their mechanisms, and to find common and unique properties of different biomaterial cues. Moreover, this work aims to establish a mechanogenomic map facilitating highly functionalized biomaterial design, and renders new thoughts of epigenetic regulation in controlling cell fates in disease treatment and regenerative medicine.
Introduction
B
Epigenetics refers to heritable changes in gene expression pattern that do not involve alterations in the DNA sequence. The epigenome helps to stabilize cell status, which can be perturbed during stem cell differentiation and somatic cell reprogramming. 25 Interestingly, biomaterial cues, which act as outside signals, can alter the internal epigenetic state through various epigenetic regulatory mechanisms, including chromatin remodeling, DNA methylation, and noncoding RNA. Chromatin remodeling is an essential step in cellular deformability, genome integrity, transcription control, and cellular memory. Besides transcriptional regulation by transcriptional factors, gene expression can also be turned on or off by changes in chromatin structure, which are modulated by a number of posttranslational modifications at the amino-terminal tails of nucleosomal histones, including acetylation, methylation, phosphorylation, and adenosine diphosphate ribosylation. The acetylation status of histones is a dynamic process, balanced by the activities of histone acetyltransferases and histone deacetylases (HDACs), whereas the methylation of histones is balanced by histone methyltransferase and histone demethylase. Generally, higher methylation levels of H3K4, lower methylation level of H3K9 and H3K27, histone acetylation, and DNA demethylation lead to gene activation. On the contrary, H3K4 demethylation, H3K9 and H3K27 methylation, histone deacetylation, and DNA methylation lead to gene repression. Noncoding RNAs are encoded by genes, but are not translated into proteins. 26 Instead, the structures of noncoding RNAs allow them to interact functionally with various biochemical processes, such as chromatin structure, transcription, and translation.
Mechanical tuning of cell geometry and the induced chromosome alterations could serve as an innovative approach to cross epigenetic barriers. A clearer mechanism of how biomaterials influence stem cell functions is essential for new biomaterial research and development. Biomaterial–cell interactions can be either utilized in tissue engineering to guide stem cell lineage commitment or in disease treatment such as tumor and fibrosis. One of the main challenges in the field of tissue engineering resides in finding the optimal cellular microenvironment for controlling cell fate and enhancing maturation.17,27,28 Similarly, in the field of oncotherapy, specific cellular shapes will lead to tumor necrosis or provide useful information for prognosis evaluation. 29 Due to the diversity of materials and the complexity of the signaling milieu, the mechanisms of how geometric cues direct stem cell fate remain vague. Thus, establishing a mechanogenomic map will provide a quantitative framework to understand how altering cellular homeostasis in local tissue microenvironments can lead to diseases in the field of oncotherapy, and to elucidate how to tailor materials for specific tissue engineering in the field of regenerative medicine.
Although a few recent studies have focused on how biomaterials direct cell behaviors, such as cell deformation, proliferation, differentiation, and reprogramming, the results lack correlation and summarization owing to the heterogeneity in biomaterials and diverse epigenetic modulations. Therefore, a systematic classification of the various material cues and the consequent epigenetic effects on different cells should be summarized to reveal the relationship between biomaterial cues and epigenetic regulation. In this study, we systematically reviewed the underlying epigenetic mechanisms of how different biomaterial cues influence cell functions, to establish a mechanogenomic map facilitating highly functionalized biomaterial design, and provides novel insights into epigenetic manipulation in controlling cell fates in disease treatment and regenerative medicine.
Methods
PubMed was used as the primary search database. The following keywords and their combinations were used: (topography OR biomaterial OR pattern OR micropattern OR nanopattern OR geometric) AND (epigenetic OR histone OR chromatin OR nuclear OR reprogramming OR DNA methylation OR noncoding RNA). The search resulted in 44,099 articles. The abstracts of the articles were screened based on the following inclusion criteria:
- Only original research articles, but not reviews, were included. - Articles should focus on how biomaterials influence cell behaviors at the epigenetic level. - Only studies with cells cultured on biomaterials other than tissue culture plastics were included. - We focus on the inherent influences from biomaterials; therefore, articles which investigated materials as a simple delivery platform of drugs, chemicals, or noncoding RNAs were excluded.
The abstracts of 44,099 articles were read and selected by two researchers, respectively. The full texts of 187 articles were further checked and 20 articles fulfilled the above inclusion criteria. Reference tracking was performed on full texts of the resulting articles to find missing articles that satisfied the inclusion criteria. The final number of included articles was 25 (Fig. 1).
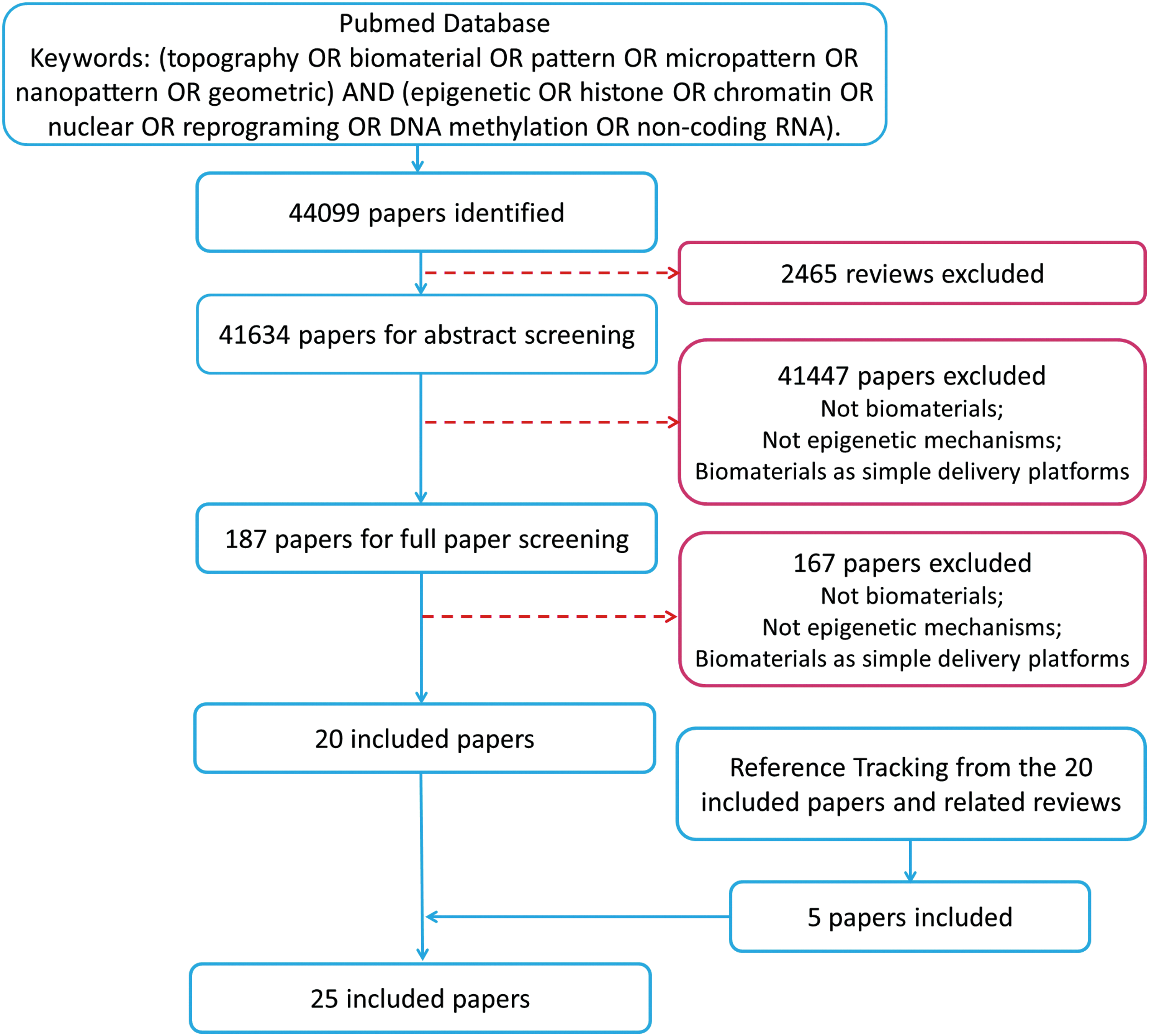
Flowchart of systematic review. Color images available online at www.liebertpub.com/teb
Results
Based on the included research articles, biomaterial cues can be divided into four categories: topography (including shape, dimension, microstructure, or nanostructure, and three-dimensionality), material nature/chemistry, stiffness/elasticity/rigidity, and mechanical forces/stimulus. Basic cell behaviors or functions guided by biomaterials include nuclear deformation, differentiation, reprogramming, and proliferation. Among the involving epigenetic regulators are chromatin remodeling and histone modification, DNA methylation, and noncoding RNA. This systematic review aims to elucidate the relationship among the three main factors, biomaterial cues, epigenetic regulation, and cell behaviors (Fig. 2).
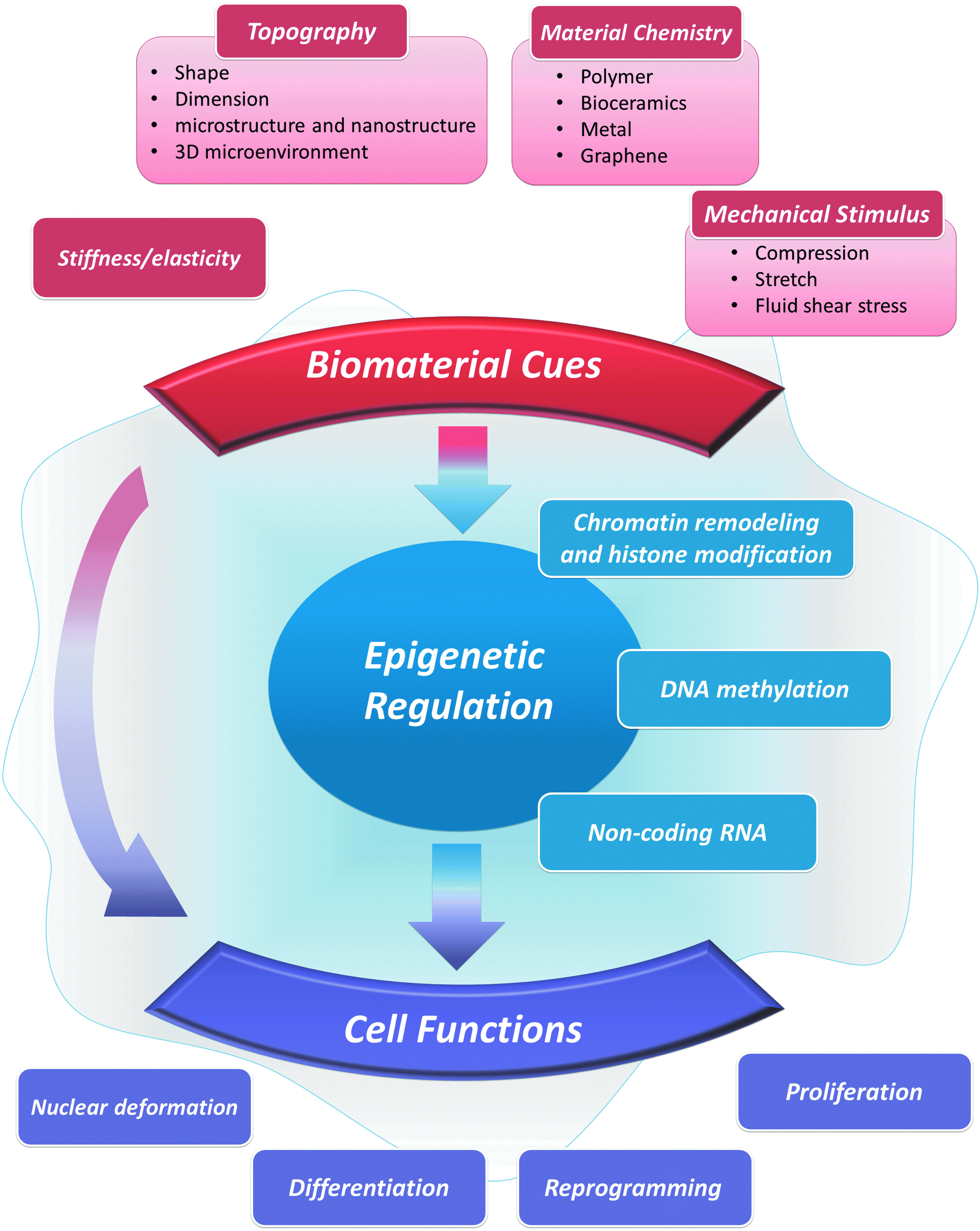
Network of systematic review demonstrating the basic relationships among biomaterial cues, epigenetic regulation, and cell functions. Color images available online at www.liebertpub.com/teb
With respect to cell behaviors (Fig. 3a), 14 out of the 25 articles investigated nuclear deformation1–7,9–12,15,30,31 (Table 1 and Fig. 4). Eight articles investigated stem cell differentiation3,8,13,14,32–35 (Table 2). Four articles investigated cell reprogramming 36–39 (Table 3 and Fig. 5). Four studies investigated cell proliferation3,6,11,31 (Table 4 and Fig. 6). Three articles investigated two factors, nuclear deformation and proliferation. One article investigated nuclear deformation, proliferation, and differentiation, and was included in the above three categories.
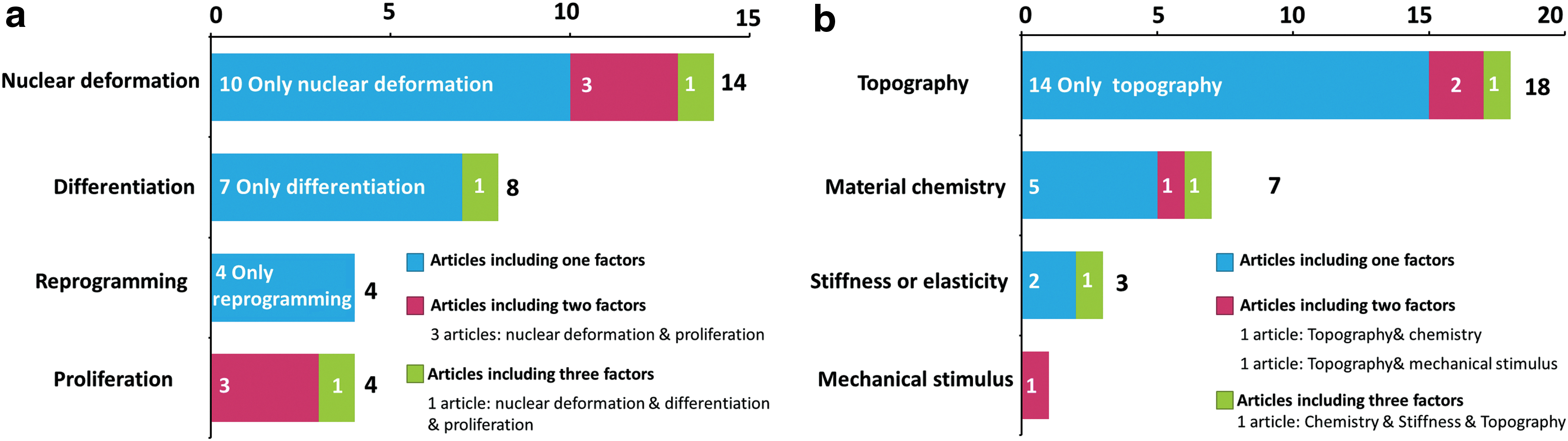
Number and distribution of investigated factors in the included articles.
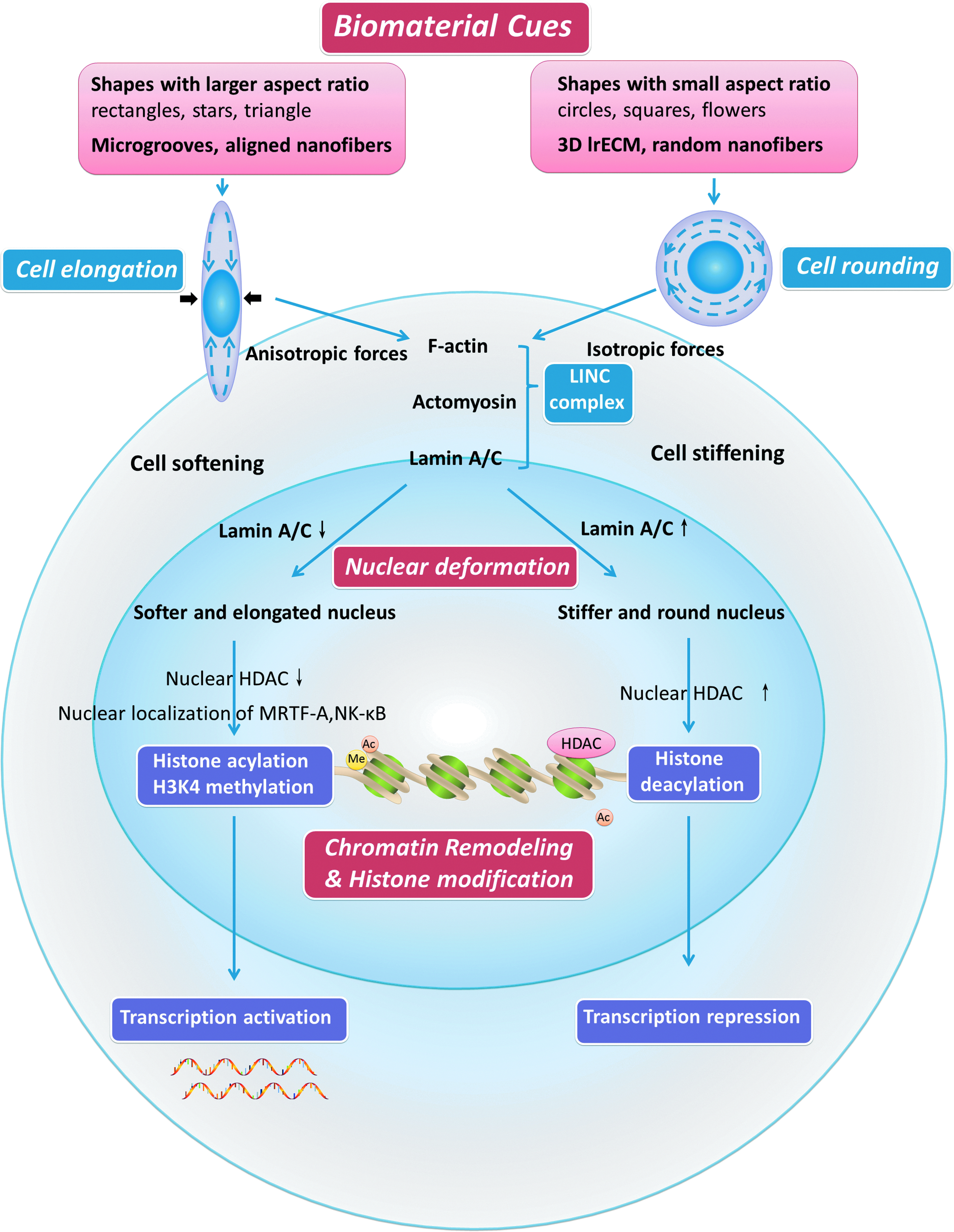
Biomaterial cues regulate epigenome through nuclear deformation induced by cell shape alteration and cytoskeleton reorganization. Elongated cell shapes directed by biomaterial cues lead to anisotropic forces of cytoskeleton, which transmit forces to nuclear protein lamin A/C through LINC complex, and consequently causing higher level of histone acetylation, H3K4 methylation, and transcription activation. Cell rounding leads to isotropic forces of cytoskeleton, higher lamin A/C level, and consequently causing decrease of histone acetylation, H3K4 methylation, and transcription repression. LINC, linkers of nucleoskeleton and cytoskeleton; HDAC, histone deacetylases; MRTF, myocardin-related transcription factor. Color images available online at www.liebertpub.com/teb
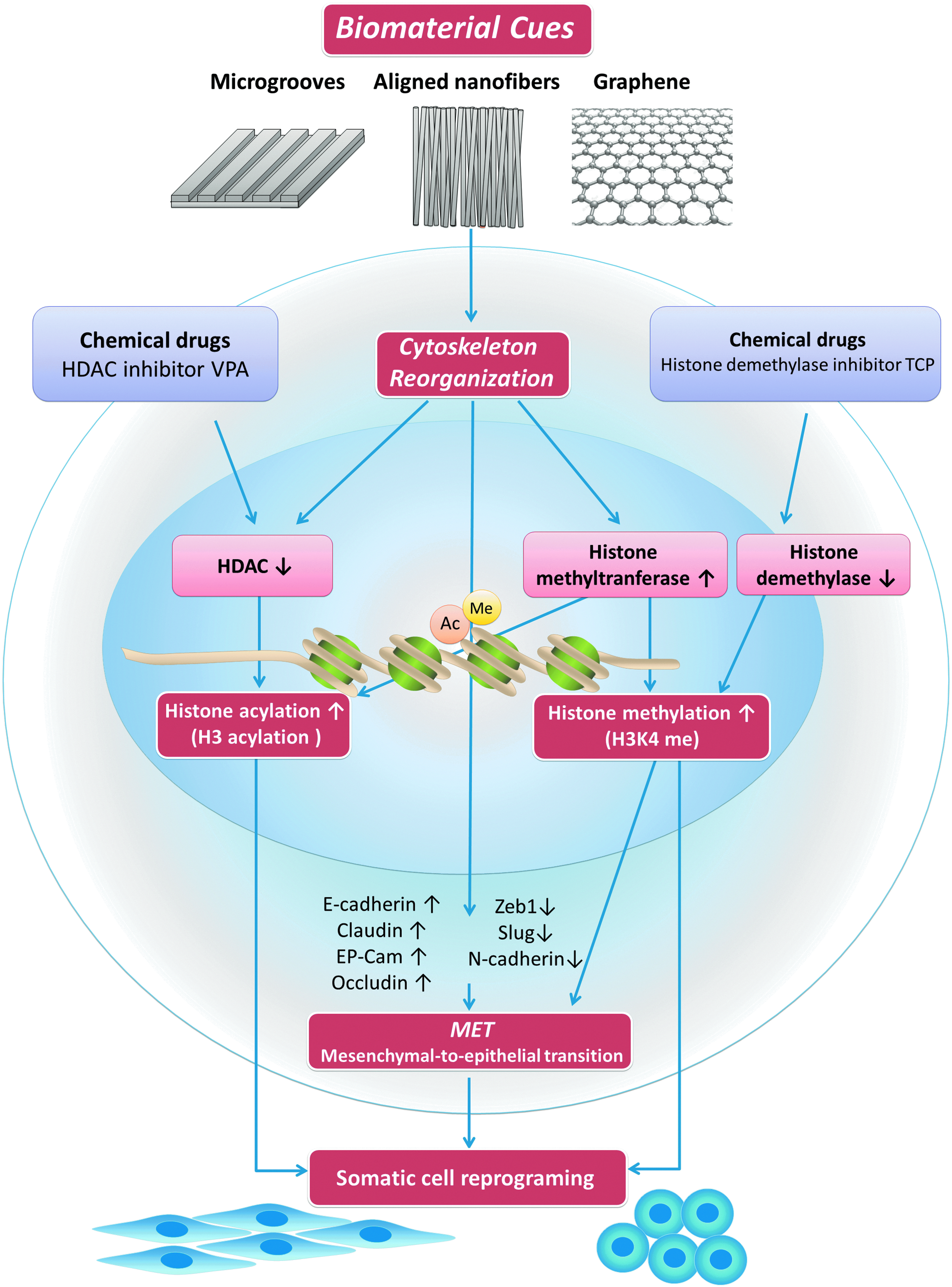
Biomaterial cues improve reprogramming efficiency through epigenetic regulation. Biomaterials, such as microgrooves, aligned nanofibers, and graphene, provoke an increase in histone acetylation and H3K4 methylation, through downregulation of HDAC activity and upregulation of histone methyltransferase, and also promote MET, thus, significantly enhancing reprogramming efficiency and replacing the effects of chemical inhibitors. HDAC, histone deacetylases; MET, mesenchymal-to-epithelial transition; VPA, valproic acid, a histone deacetylase inhibitor; TCP, tranylcypromine hydrochloride, a histone demethylase inhibitor. Color images available online at www.liebertpub.com/teb

Biomaterial cues regulate cell proliferation through epigenetic regulation. Larger spreading area with the same cell shape leads to larger nuclear volume, chromatin decondensation, increased DNA synthesis, and increased cell proliferation. Cell elongation with the same spreading area leads to chromatin condensation, decreased DNA synthesis, and lower proliferation rate. Color images available online at www.liebertpub.com/teb
HDAC, histone deacetylases; PDMS, polydimethylsiloxane;
As for biomaterial cues (Fig. 3b), out of the 25 single studies, 18 studies1–13,31,33,34,37,39investigated topography, 7 studies4,14,30,33,35,36,38 investigated material nature and chemistry, 3 studies4,15,32 investigated material stiffness/elasticity, 1 study 9 investigated mechanical stimulus, such as compression or stretch stresses. Among the 18 studies which focused on topography, 7 articles investigated microgrooves or gratings or stripes,3,8–10,31,37,39 2 articles investigated micropillars,4,7 5 articles investigated micropatterns such as circles, rectangles, squares, and triangles,1,2,5,6,11 3 articles investigated nanotopography, such as nanofibers (which also investigated microgrooves), 37 or nanotubes, 34 or nanohydroxyapatite, 33 and 2 articles investigated three-dimensionality.12,13 Also, there were studies that investigated more than 2 or 3 factors, as shown in Figure 3b.
Materials used in included articles were mainly polymers, metal, bioceramics, and graphene, among which polymers were most investigated. Synthetic or natural polymers1–5,7,9,10,12,30,36,37,40 included polydimethylsiloxane (PDMS), PLGA, Poly-l-lactic acid or poly(l-lactide-co-caprolactone), polyacrylamide hydrogels, collagen, and fibronectin. Bioceramics included hydroxyapatite. 33 Meanwhile, graphene35,38 and metal materials 34 were also included.
Discussion
The main purpose of this review is to summarize the epigenetic effects of biomaterial cues on cell behaviors, to compare the differences and similarities between different biomaterial cues, to reveal underlying relationship among epigenetic alterations, mechanical transduction and chemical signaling pathways, and to provide important insights to biomaterial designing for clinical applications.
Definition and classification of biomaterial cues
There are various characteristics of biomaterials that can dictate cell behaviors in different manners. Hence, a clear definition and classification of biomaterial cues are urgently needed to summarize and categorize previous investigations. In this study, we define biomaterial cues into the following categories: material nature/chemistry, stiffness/elasticity/rigidity, mechanical forces/stimulus, and topography (including shape, dimension, microstructure or nanostructure, and three-dimensionality). Due to the diversity and complexity of biomaterial cues, exploring the mechanisms by which cells respond to inherent material characteristics remains challenging. Therefore, setting a single variable and controlling the other characteristics of a material is important in the researches to explore complex mechanisms. Basing on a clear plot of regulating mechanisms from each biomaterial cue, complex and personally designed biomaterials can be engineered to meet the different needs for clinical applications.
Biomaterial cues regulate epigenetic state
Biomaterial cues regulate epigenetic state in the manners of chromatin remodeling, histone modification, and noncoding RNA, whereas the influences on DNA methylation remain limited according to the existing researches. Mechanical transduction from membrane to nucleus is the basis of chromatin remodeling and histone modification induced by biomaterial cues, which is also supported by microarray analyses demonstrating large-scale changes in gene expression of nuclear morphology, actomyosin contractility, and histone acetylation.2,5 The moment a cell adheres to the substrate, their interactions begin. The cell spreads and thus cell shape changes under the guidance of material substrate. Material adhesiveness, micro- and nanotopography, and three-dimensionality will all lead to reorganization of the cytoskeleton. Forces generated by the cytoskeleton components, including actin and myosin can be transmitted to the nucleus and chromatin through physical links on the nuclear envelope and the lamin meshwork. The physical properties of the nucleus govern the spatiotemporal packaging of chromatin. Therefore, when active forces modulate nuclear morphology, chromatin remodeling takes place in the nucleus. Chromatin remodeling, including packaging and release of histones and DNA, and histone modification as well, regulates transcription activation and repression, thus influencing cell behaviors and stem cell lineage commitment. Histones, assembled with DNA to form nucleosomes, are the basic building blocks of chromatin. Therefore, histone modifications and chromatin decondensation are central to the remodeling process of chromatin, which is important for cell functions, such as proliferation, differentiation, and reprogramming.
A variety of topographies have been investigated in different studies. Collectively, micro- or nanogeometry or 3D microenvironment can generally guide cells into two types of shapes, cell shapes guided by anisotropic forces and cell shapes guided by isotropic forces. The former includes a variety of cell shapes, such as elongated shapes guided by parallel microgooves,9,31 aligned nanofibers, 37 and single cell topography such as rectangles with larger aspect ratios.5,6,11 The latter one is generally a round shape.12,30 For equal spreading, cell elongation leads to decreased cell stiffness, 11 elongated nuclear shape and decreased nuclear matrix protein lamin A/C. 3 Meanwhile, chromatin decondensed with a concomitant increase in histone acetylation caused by decreased HDAC activity, 37 and thus an increase in gene expression. Cytoplasmic actomyosin and nuclear matrix protein lamin A/C are important transducers in this process, since inhibition lamin A/C disrupts this process 11 and cytoplasmic-to-nuclear redistribution of HDAC depends on actomyosin. 2 On the contrary, 3D lrECM (laminin-rich ECM), 12 as well as nonadhesive materials 30 and circle micropatterned substrata, 2 leads to cell rounding. Cell rounding involves global histone deacetylation, specifically, deacetylation of histones H3 and H4, chromatin condensation and a decrease in nuclear size, a global increase in chromatin condensation, and a reduction in gene expression.2,12,30 Disruption of the actin cytoskeleton with cytochalasin D phenocopies the lrECM-induced cell rounding and histone deacetylation. 12 Based on these studies,9,37 elongated cell shapes directed by biomaterial cues lead to increased histone acetylation and H3K4 methylation, where the actin cytoskeleton transmits forces to nuclear protein lamin A/C, consequently causing transcription activation (Fig. 4 and Table 1).
Mechanical stimulus, such as compression, stretches, and fluid shear stress, which offers dynamic changes to cellular microenvironment, can induce nuclear deformation and changes in histone modifications. Cells can sense forces perpendicular to their spreading axis, which is similar to topography-guided cell elongation, where orientation and deformation of the nucleus are regulated by lateral compressive forces driven by tension in central actomyosin fibers. 9 Uniaxial mechanical forces, perpendicular to substrate guidance such as microgrooves, can cause not only changes in nuclear shape and nuclear orientation, but also a decrease in HDAC activity accompanied by increased histone acetylation. Similarly, in respect of fluid shear stress, which influences cell migration, significantly altered migration direction and increased migration tortuosity is observed when the flow is oriented perpendicular to the topography. 41 Therefore, similar to cell elongation, which generates internal forces perpendicular to cell spreading axis, external forces perpendicular to substrate topography also induce decreased HDAC activity and increased histone acetylation. 9 However, material stiffness or elasticity does not seem to have a dominant effect on nuclear deformation. 4
Biomaterial cues direct cell functions through epigenetic regulation
Cell proliferation
Biomaterial cues can alter cell shape, and the changes in cell shape and spreading area deform and orientate the nucleus, thus influencing cell proliferation. Endothelial cells and vascular smooth muscle cells are most studied in cell shape-directed proliferation, since they are important for vascular regeneration and remodeling, as well as wound healing. According to included articles related to cell proliferation,3,6,11,31 larger nuclear volume leads to increased cell proliferation, whereas chromatin condensation and less DNA synthesis lead to lower proliferation rate. To distinguish the effects of cell shape and spreading area, micropatterned islands of well-defined shapes and areas are fabricated by microcontact printing.6,11 In the two included single cell researches, spreading area and cell shapes are well controlled to investigate the influence of these factors, respectively, independent of cell–cell interactions. It is a consensus that larger spreading area with the same cell shape (nonelongated cells such as circle and square shape) leads to higher F-actin content and contractility, increased cell stiffening (higher storage modulus), larger nuclear volume, increased DNA synthesis, and increased cell proliferation (Fig. 6). With the same spreading area, Versaevel et al. report that cell elongation leads to a decreased nuclear volume, cell softening (lower storage modulus), a drastic condensation of chromatin, and dramatically decreased cell proliferation. 6 Roca-Cusachs et al. demonstrate a slight decrease in nuclear volume and proliferation rate in elongated cells, but there are no significant differences. 11 These slightly different results are likely caused by the different extent of elongation and control of spreading area (Table 4). The relationship between cell elongation and lower proliferation rate is also confirmed in the other two researches based on micro- or nanotopography. Microgrooves with 10 μm width lead to obvious cell elongation and a decrease in cell spreading compared with nonpatterned substrate. Elongated smooth muscle cells demonstrate lower proliferation rate compared with cells in a spread-out morphology. 31 DNA microarray profiling reveals a lower expression of neuron-derived orphan receptor-1 (NOR-1) in elongated smooth muscle cells. Knocking down NOR-1 suppressed DNA synthesis, suggesting that NOR-1 is a mediator of cell elongation effects. Human mesenchymal stem cells (MSCs) on nanogratings of 350 nm linewidth demonstrate a similar result. 3 Collectively, cell elongation leads to chromatin condensation, decreased DNA synthesis, and lower proliferation rate, whereas cell spreading leads to chromatin decondensation, increased DNA synthesis, and promote proliferation (Table 4 and Fig. 6).
Somatic cell reprogramming
Cell reprogramming represents a major advancement in cell biology and has wide applications in regenerative medicine, disease modeling, and drug screening. Patient-specific somatic cell reprogramming is likely to have a large impact on medicine by providing a source of cells for disease modeling and regenerative medicine. In 2006, terminally differentiated mammalian cells were reprogrammed for the first time to a pluripotent state by transfer of a combination of defined genetic factors. 42 Now it is well recognized that induced pluripotent stem cells (iPSCs) can be produced from somatic cells by the forced expression of four or fewer transcription factors (for example, Oct4 (O), Sox2 (S), Klf4 (K), and c-Myc (M), or Oct4, Sox2, Nanog, and Lin28).43–45
However, low reprogramming efficiency and safety consideration has hindered its clinical transformation. Barriers for reprogramming are known to be attributable in epigenetic state, reflecting the remarkable stability of the differentiated state of somatic cells. The success of nuclear reprogramming is limited by epigenetic mechanisms that stabilize the state of gene expression in somatic cells and thereby resist efficient reprogramming. Transcription factors, chromatin modifications, and noncoding RNAs can regulate the efficiency of reprogramming. 46 Histone H3 acetylation (AcH3) is known to be a permissive environment for reprogramming by stemness factors, thus HDAC acts as an inhibitor of reprogramming. HDAC inhibitor chemical compounds, such as valproic acid (VPA), have shown great promise in enhancing the efficiency of iPSC formation and even eliminate the need for some oncogenic transcription factors such as c-Myc and Klf4. Meanwhile, one of the first noticeable changes during the reprogramming of somatic fibroblasts is their transformation into tightly packed clusters of rounded cells in a process that resembles mesenchymal-to-epithelial transition (MET), which plays an essential role in initiating reprogramming into iPSCs. 47 Higher H3K4 methylation level is correlated with MET process, thus histone demethylase inhibitors, such as tranylcypromine hydrochloride, can also increase the efficiency of cell reprogramming (Fig. 5).
Interestingly, biomaterial topography has been discovered to replace the effects of small-molecule epigenetic modifiers and significantly improve reprogramming efficiency, providing a safer and more convenient alternative to cell reprogramming. Specifically, topographies of microgrooves or aligned nanofibers provoke an increase in AcH3, H3K4me2, and H3K4me3, through downregulation of HDAC activity and upregulation of the expression of WD repeat domain 5 (WDR5), a subunit of H3 methyltransferase, thus significantly enhancing reprogramming efficiency in fibroblasts. Blebbistatin, inhibitor of myosin-II disrupting actomyosin contractility, can eliminate these effects. Meanwhile, parallel microgrooves have also drawn attention in the field of cardiac tissue engineering. No single cocktail or reprogramming strategy has yet become a gold standard for conversion of somatic cells to cardiomyocyte-like cells because of the low reprogramming efficiency. Under the existence of myocardin, Tbx5, and Mef2c, microgrooves mimic the effects of VPA and markedly augmented the expression of cardiomyocyte-specific proteins. In addition, the anisotropy resulting from parallel microgrooves can induce cellular alignment, mimicking the native ventricular myocardium and augmenting sarcomere organization. 39 Moreover, graphene can also improve cellular reprogramming efficiency by inducing MET process, which is known to affect H3K4me3 levels. 38 Compared with small-molecule drug, using biomaterials to enhance cell reprogramming efficiency is safer and, therefore, more potential in the clinical use of human cells (Table 3 and Fig. 5).
Stem cell differentiation
Stem cells demonstrate preferential lineage biases during differentiation by responding to material cues including topography, material chemistry and substrate elasticity. Therefore, tailored biomaterials play important roles in osteogenic, chondrogenic, neurogenic, and cardiogenic differentiation. Biomaterials influence stem cell lineage commitment and facilitate differentiation of stem cells especially under the existence of chemical induction medium. The structural composition of both the nucleus and the cytoskeleton alters in differentiated stem cells, but whether nuclear stiffness increases or decreases with onset of differentiation remain controversial. Some researches propose that with the beginning of differentiation, nuclear stiffness increases, which means increased lamin A/C level.2,48,49 While the others hold the idea that nuclear stiffness decreases with lower lamin A/C level. 3 Unlike abundant studies on cell shape and nuclear deformation, researches on how epigenetic regulations modulate stem cell linage commitment remain limited. The stem cell transcriptome also exhibits variable expression of key regulatory gene products that is utilized for distinct differentiation potential and possibly provides heterogeneous mechanical properties to stem cells. In addition, activation of myocardin-related transcription factor, an actin-dependent transcription cofactor, have been shown to upregulate its target genes involved in actin cytoskeleton pathway to stabilize nuclear morphology during cellular differentiation.
Histone modification is an important epigenetic regulation method which influences gene expression and cell differentiation. Our research group has revealed how nanotubes promote osteogenic differentiation through epigenetic regulation. Titanium dioxide (TiO2) nanotubes with the dimension of 70 nm diameter promote the osteogenic differentiation especially under the condition of osteogenic medium, and this promotion is based on enhanced H3K4 methylation level at the promoter regions of osteogenic-related genes by inhibiting histone demethylase retinoblastoma protein 2 (RBP2). 34 Later, we discover similar phenomenon in single-layered graphene. 35 Another research also demonstrates that retinoblastoma protein are downregulated in MSCs cultured on PDMS substrates with nanogratings of 350 nm linewidth, 700 nm pitch, and 280 nm depth compared with planar substrates. 3 Therefore, common mechanisms underlie how biomaterials promote osteogenic differentiation of cells (Table 2).
Meanwhile, with the recent progress of micro-RNAs (miRNAs) containing highly conserved noncoding RNAs with lengths ranging from 18 to 23 nucleotides, 50 it becomes clear that the signaling networks are even more precise and complex with miRNAs participating in almost every cellular process. Given the pivotal role of miRNAs in regulatory circuitries that control self-renewal and pluripotency of stem cells, the important roles of miRNAs in cell–substrate interactions need to be systematically investigated. Now, there are only a few researches. PHBHHx promote the chondrogenesis of mouse MSCs with the existence of chondrogenic medium. According to high-throughput analysis, differentially expressed genes include miR-29a and miR-29b, which directly target 3′UTR of Col2a1 encoding type II collagen, and their activities are regulated by Sox9, the SRY-related high-mobility group box gene 9. 14 On the other hand, 10 μm-wide microgrooved PHBHHx promote osteogenic differentiation of MSCs with increased expression of osteogenesis-related marker genes, including Cbfa1, Col1a1, and Bmp2. The miRNA microarrays reveal that 18 differentially expressed miRNAs on microgrooved pattern had multiple target genes, contributing comprehensively to the cellular regulation process, including MAPK and Smad signaling pathways 8 (Table 2).
DNA methylation leads to gene suppression. DNA methylation refers to addition of a methyl group to a cytosine base residue in DNA, which often localizes next to a guanine base. Methylated cytosines can be further modified by hydroxylation. Methylated cytosines can lead to the recruitment of specific methyl DNA-binding proteins, which may lead to transcriptional repression. The influence of biomaterials on DNA methylation remains limited. Although it is well recognized that substrate elasticity directs stem cell lineage commitment, DNA methylation profiles remain unaltered as the elasticity of the hydrogel substrate varies. 32 Nano-HAp is usually considered to promote osteogenic differentiation of MSCs, but a significant increase in ALP promoter methylation leading to unexpected suppression of ALP, BSP, and OSC RNA levels is reported. 33 Therefore, further researches on biomaterial cues and DNA methylation are expected.
Similarities and differences of cell functions guided by various biomaterial cues
Topography
Topography, including shape, dimension, microstructure or nanostructure, and three-dimensionality, is one of the most investigated factors in material–cell interactions, and can dictate various cell behaviors, including cell proliferation,6,11,31 differentiation,51,52 and reprogramming.37,39 Topography regulates cell behaviors by influencing cell adhesion, spreading, and subsequent cytoskeletal reorganization leading to changes in cellular mechanical transduction, nuclear alterations, and consequently changes in gene expressions. These regulations depend on cytoskeletal mechanical transduction system, and interruptions to actomyosin or lamin A/C block these substrate–cell interactions. With the same cell shape, larger spreading area induce larger nuclear and transcription activation, and smaller area lead to chromatin condensation and a global reduction in gene expression 12 (Fig. 4). With the same spreading area, different shapes, such as circle versus square or rectangles, circle versus triangle, and flowers versus stars, lead to isotropic forces or anisotropic forces and consequently influence cell proliferation and differentiation. Topographies which guide cells into elongated shapes, such as microgrooves and aligned nanofibers, lead to global enhancement of histone acetylation and H3K4 methylation, thus facilitating the process of reprogramming and neural differentiation. Topographies which guide cells into more spreading and polygonal shapes, such as nanotubes 34 and random nanofibers, 22 generally promote the process of osteogenic differentiation and lead to higher proliferation rate. Meanwhile, microtopography and nanotopography regulate cell behaviors from different signaling pathways. In 3D environment, round cells demonstrate decreased histone acetylation level. Microtopography influences Rho/RhoA signaling pathway.21,53 As for nanotopography, such as nanotubes and nanogrooves, leads to differential expression of genes mainly involved in Wnt/β-catenin signaling pathway,23,53,54 whereas nanogratings can influence gene expression in both Rho/RhoA and Wnt signaling pathways. 55
Stiffness or elasticity
Stiffness or elasticity influences cell differentiation, nuclear deformation, and cell spreading. Stiffer substrates lead to decreased nuclear height 56 and larger cell spreading area.16,56 As for differentiation, stem cells commit to specific lineage whose original niches are similar with the existing matrix stiffness. For example, soft matrices (0.1–1 kPa) that mimic brain are neurogenic, stiffer matrices (8–17 kPa) that mimic muscle are myogenic, and comparatively rigid matrices (>34 kPa) that mimic collagenous bone prove osteogenic. 17 Stiffness or elasticity is usually controlled under the same material substrate by changing the thickness or component ratio of the material. Elastomeric polymer networks have been commonly used, as their elasticity can be controlled by simply adapting the density of crosslinks in the network. Cells on the substrate with low elastic modulus cannot form stable focal adhesions and differentiate as a result of decreased activation of the extracellular signal-related kinase (ERK)/mitogen-activated protein kinase (MAPK) signaling pathway. 16 Interestingly, stiffness-induced mechanical dosing can be memorized with YAP/TAZ acting as an intracellular mechanical rheostat—that stores information from past physical environments and influences the cells' fate. 57 Moreover, similar as topography-induced cell function alterations, inhibition of nonmuscle myosin II also blocks all elasticity-directed lineage specification, but without strongly perturbing many other aspects of cell functions and shapes.17,32 Meanwhile, different elasticity of the substrate has little effect on DNA methylation pattern. 32 Therefore, elasticity-induced lineage commitment depends on cytoskeletal mechanical transduction, resulting in heritable changes in YAP/TAZ and ERK/MAPK signaling pathways, but the changes inside the nucleus remain uncovered.
Mechanical stimulus
Mechanical stimulus, such as compression, stretches, and fluid shear stress, offers dynamic changes to cellular microenvironment. Cells can sense forces perpendicular to their spreading axis, which is similar to topography-guided cell elongation. 9 Similar to cell elongation, which generates internal forces perpendicular to cell spreading axis, external forces perpendicular to substrate topography also induce decreased HDAC activity and increased histone acetylation resulting from tensions in central actomyosin fibers.
Material chemistry
Material chemistry influences cell behaviors in a more complex manner. Polymers, ceramics, and metals have historically dominated the application of materials in medicine. Polymers enjoy the advantages of good controllability of both stiffness and topography. Therefore, it can be used in a variety of studies, including elasticity, mechanical stimulus, and topography. Metal materials enjoy the advantages of good rigidity, but the fabrication of precise topography is relatively difficult. Methods of anodic oxidation, 34 micro-arc oxidation, 58 and femtosecond laser59,60 have been used for the fabrication of micro- or nanotopography of metal materials. In spite of conventional metal materials, such as titanium, biodegradable magnesium alloys61–64 and metal ions incorporated materials58,65 have attracted a lot of attention. It is reported that Li+ promotes osteogenic differentiation by influencing canonical Wnt signaling pathway, 65 whereas Ca2+ influences noncanonical Wnt signaling pathway. 66 Other metals, such as tantalum67–69 and strontium,58,70–73 have been reported to enjoy appraisal biocompatibility recently, and their underlying mechanisms are expected to be revealed before their clinical applications. Bioceramics enjoy good biocompatibility for bone tissue engineering,74,75 but its shape or topography is relatively difficult to control. Carbon-originated materials, including graphene and its derivatives,76–80 as well as carbon nanotubes or nanofibers, form another kind of newly discovered biocompatible materials. Graphene, a two-dimensional crystalline allotrope of carbon, is one of the most attractive carbon materials that can be described as a one-atom-thick layer of graphite. Recently, graphene has been considered to be a suitable material as a strong, noncytotoxic, implantable platform for use in biomedical engineering. Graphene can influence both signaling pathways and histone modification,35,81 and it is still controversial which characteristics (material chemistry, topography, or its special characteristic of electric charges) cause these changes in epigenome and cell functions.81–83 With the development of new materials, a deeper insight into their mechanisms and safety evaluations are of paramount importance before their clinical applications.
Although different biomaterial cues influence cell functions from various regulating methods, the similarities among different kinds of biomaterial cues (topography, elasticity, and mechanical stimulus) lie in cytoskeletal reorganization and cellular-to-nuclear mechanical transduction. In addition, breaking the epigenetic barriers is the common first step of cell differentiation and reprogramming.
Applications
Establishing a mechanogenomic map for tailored material design
Tailored biomaterials will be widely applied not only in the field of tissue engineering, but also disease treatment. In the field of tissue engineering and regenerative medicine, biomaterials are designed to provide the most favorable microenvironment for seed cells to adhere, spread, communicate, and differentiate into expected linages. 84 Meanwhile, in the field of disease treatment, biomaterials are used to predict prognosis and induce apoptosis of cancer cells. 29
A mechanogenomic map of how biomaterials influence cell functions is essential for tailored and highly functionalized biomaterial design and engineering for clinical applications, therefore, a summarization of both epigenetic alterations and signaling pathways of each biomaterial cues will provide an important basis for mapping the whole picture. Several studies have investigated the changes in genomic profiling under the guidance of biomaterial cues, and most of differentially expressed genes are members of Rho/ROCK, MAPK, ERK, Wnt, YAP/TAZ, and JNK signaling pathways according to KEGG and Gene Ontology analysis.5,19,54,55 However, an epigenomic profiling of how biomaterial cues influence the epigenetic state of cells are expected to be revealed.
Epigenetic alterations as an important safety evaluation tool for biomaterials
Epigenetic regulation of biomaterials not only plays an important role in regenerative medicine but also can be a potential tool for safety evaluation of biomaterials. Epigenetic alteration is a mechanism which can explain the long-lasting and memorable effects of biomaterial cues. Meanwhile, epigenetic regulation is an important step that breaks through the barrier of cell lineage commitment. Recent technological advances have led to rapid progress in the characterization of epigenetic modifications that control gene expression in a generally heritable way, and are likely involved in defining cellular phenotypes, developmental stages, and disease status from one generation to the next. Epigenomic profiling has the potential to provide mechanistic information in toxicological assessment.85–87 The assessment of epigenetic effects may also be approached with new model systems that could directly assess transgenerational effects or potentially sensitive stem cell populations. These would enhance the range of safety assessment tools for evaluating newly designed materials.
Meanwhile, inflammation is an important and inevitable process of material–host interaction, which may alter epigenetic state. Proinflammatory activities of cytokines, such as interleukin (IL)-1, IL-6, and tumor necrosis factor-alpha (TNF-α), are lower on TiO2-coated titanium alloy (Ti6Al4 V) implants compared with silica (SiO2)-coated implant and stainless steel. 88 Meanwhile TiO2 nanotubes attenuate macrophage inflammatory activity through inhibition of MAPK and NF-κB pathways. 89 It is also reported that collagen membranes coated with graphene oxide demonstrate lower PGE2 levels, and less TNFα and COX2 expression in human dental pulp stem cells. 90 But whether these biomaterial-related inflammation process leads to epigenetic changes remain undiscovered. Therefore, epigenetic changes induced by inflammation process after biomaterial implantation are expected to be uncovered, and this will also become a paramount parameter for the evaluation of biomaterial compatibility.
Outlook
Noncoding RNAs play important regulating roles in various biochemical processes. 26 It has been reported that various microRNAs regulate cell functions under the existence of biomaterial cues. For example, it has been well recognized that microRNAs regulate important signaling pathways, such as Wnt, MAPK, Smad, and BMP signaling pathways. Meanwhile, microRNAs act as switches for MSCs to differentiate into either osteogenic or adipogenic lineage. 8 Different miRNAs have been reported to regulate a master transcription factor for osteogenesis, such as Runx2, as well as molecules in the Wnt or BMP signaling pathway, and control the balance between osteoblast and adipocyte differentiation. 91 With the discovery of long noncoding RNA and circular RNA, the researches on noncoding RNAs will go deeper and more regulating mechanisms remain to be investigated.
High-throughput biomaterial selection92,93 offers us an opportunity for mass screening when the underlying mechanisms are vague. This was a breakthrough for material discovery, but we are eager to embrace the era for material design and customization. Further studies are expected to investigate how epigenetic regulations correlate with chemical signaling pathways. A clear picture of the network among epigenetic alterations, signaling milieus, and cell functions will provide paramount information for disease treatment and regenerative medicine. Only by revealing the mechanism of how different biomaterial cues direct cell functions, can we design more intelligent and highly functionalized biomaterials from material chemistry chosen, to topography design, and then 3D construction to meet complex clinical needs.
Conclusion
Biomaterial cues, including material nature/chemistry, stiffness/elasticity/rigidity, mechanical forces/stimulus, and topography (including shape, dimension, microstructure or nanostructure, and three-dimensionality) can influence cell behaviors by epigenetic regulation. Elongated cell shapes lead to cell softening, decreased nuclear lamin A/C, decreased HDAC activity, and increased histone acetylation and H3K4 methylation. Meanwhile, spreading cell shapes causes higher actin level, cell stiffening, increased nuclear volume, chromatin decondensation, increased DNA synthesis, and higher proliferation rate. Biomaterials, such as microgrooves, aligned nanofibers, and graphene, can act as promising facilitators for somatic cell reprogramming. By increasing histone acetylation, H3K4 methylation, and MET, biomaterials can replace the role of chemical drugs in the process of reprogramming. Biomaterial cues can direct stem cell lineage commitment through epigenetic regulating methods, including histone modification, DNA methylation, and noncoding RNAs.
Footnotes
Acknowledgments
This work is supported by grant (31600787, 81371118) from the National Natural Science Foundation of China, the grant of Young Elite Scientist Sponsorship Program by CAST (2015QNRC001), the Project for Culturing Leading Talents in Scientific and Technological Innovation of Beijing (Z171100001117169), and the grant of Peking University School and Hospital of Stomatology (PKUSS20150107).
Disclosure Statement
No competing financial interests exist.