
Abstract
Development of multicellular organisms is a highly orchestrated process, with cells responding to factors and features present in the extracellular milieu. Changes in the surrounding environment help decide the fate of cells at various stages of development. This review highlights recent research that details the effects of mechanical properties of the surrounding environment and extracellular matrix and the underlying molecular mechanisms that regulate the behavior of embryonic stem cells (ESCs). In this study, we review the role of mechanical properties during embryogenesis and discuss the effect of engineered microtopographies on ESC pluripotency.
Introduction
E
Previous studies have shown that stem cells respond to distinct properties of the extracellular matrix (ECM) that range from mechanical properties, such as stiffness, 11 adhesiveness, 12 geometric patterning,13,14 and topography, 15 to biochemical properties involving changes in the composition, 16 which all in turn affect the behavior of the cell. For an overview of the importance of the ECM and the underlying substratum in the context of mesenchymal stem cells, we refer the reader to a number of excellent articles and reviews.17–20
These extrinsic cues may also be harnessed to direct and control stem cell fate in the context of tissue engineering and regenerative medicine, in an attempt to deliver patient-specific cell therapies. However, we are still gaining knowledge about how in vitro culture conditions can better mimic the in vivo environment in which stem cells live. This review focuses on how properties of the ECM affect the pluripotency of ESCs while attempting to also draw correlates to the intrinsic behavior of cells residing within the developing mammalian embryo. In the following sections, we highlight examples detailing the role of the ECM in the developing mammalian embryo, the effect of mechanical stimuli that mimic the external substratum through topological cues on pluripotent stem cells, and the underlying molecular mechanism.
Embryonic Development and the Role of the ECM
The ECM is a dynamic structural component of all tissues, whose composition and constituents change through development, providing the functional cellular environment for the developing embryo. A number of studies illustrate the involvement of the ECM during early vertebrate development, and this has been particularly well characterized in the context of the chick embryo.21–25 While it is not possible to present all the instances of ECM involvement in early development, we highlight below the role of a few individual components of the ECM.
During early vertebrate embryo development (chick and mouse), fibronectin (FN) may be assembled in either a paracrine or autocrine manner. 26 FN, however, remains essential, as mouse embryos null for Fn1, present a shortened anterior–posterior axis, cardiovascular defects, and a general deficit in mesoderm, including impaired somite and notochord formation.27,28 The expression and localization of FN in mouse blastocysts, are known to be regulated in response to specific growth factors, such as insulin-like growth factor 1 (IGF-1), to enhance their attachment to endometrial cells in vitro. 29
Other ECM components such as Laminin are also expressed early during mammalian development,30,31 with Laminin β1 and β2 (LAMB1, LAMB2) chains expressed widely at different embryonic stages of human development. 32 Furthermore, the Laminin expression pattern was found to undergo major changes during the differentiation of human ESCs (hESCs), further supporting the idea that the ECM may be both instructive as well as responsive. 33 Perlecan, a five-domain heparin sulfate proteoglycan is expressed through different stages of human embryogenesis. In particular, its expression is detected at stages where an epithelial-to-mesenchymal transition is known to occur, such as during human gut development beginning at gestational week 8. 34
In addition to the composition of the ECM, physical properties of the substrate have also been shown to affect early embryonic development. Preimplantation mouse embryos cultured on softer surfaces such as collagen gels (stiffness of 1 kPa) that mimicked the in vivo uterine environment, developed faster and better from the two-cell stage to the blastocyst, with a significantly higher rate of zona hatching, compared with embryos cultured on stiffer substrates such as polystyrene dishes (stiffness of 1 GPa). In addition, embryos cultured on softer substrates developed better when transferred to recipient female mice, indicating that the physical properties of the preimplantation environment deeply affected development. 35 Preimplantation embryos were also sensitive to shear stress, resulting in an induction of phosphorylated MAPK8/9. 36 Mimicking of the in vivo environment in the fallopian tube in terms of shear stress in an in vitro culture system improved the development of embryos to the blastocyst stage, indicating that embryos are responsive to mechanical stimuli. 37 Thus, the influence of the external environment in terms of chemical composition and physical parameters is far reaching in terms of early developmental decisions.
Patterned Surfaces as In Vitro Mechanical Niche Models
In this section, we emphasize the importance of mechanical cues, such as different surface architectures, during embryonic development, and explore how this inherent property can be harnessed for ESCs for fundamental research and tissue engineering purposes. Topographies can be viewed as structures originating from a flat surface and ranging in the nano (1–1000 nm) or microtopographical (1–10 μm) range, with most research performed on mesenchymal stem cells (MSCs). Disordered nanoscale pits can stimulate the differentiation toward the osteogenic lineage, independent of the classical osteogenic supplement, dexamethasone. 38 On microtopographies, our group found improved osteogenesis of MSCs. 39 Also, MSC differentiation down the adipogenic 40 and chondrogenic 41 lineages have been previously demonstrated.
ESC self-renewal can be influenced by combining nanoroughness with geometric shapes. In this study, smooth surfaces (1 nm) support stemness, whereas nanorough surfaces (70 and 150 nm) resulted in a loss of pluripotency. 42 Furthermore, E-Cadherin (CDH1) presence was maintained on the smooth surface, a crucial regulator for ESC self-maintenance. 43 In a similar study, flat and nanorough surfaces outperformed microroughness when assessing self-renewal of ESCs. 44 Here, the flat and nanopatterned surfaces led to reduced ESC adhesion and spreading, suppressed FAK and downstream ERK signaling, improving ESC self-renewal. 45
In another study, microroughness and nanoroughness were combined (919 ± 22 nm) and compared against nanoroughness alone (68 ± 30 nm), a smooth surface, and against a feeder layer. Here, it was observed that in LIF media, the combined approach of utilizing microroughness and nanoroughness promoted formation, homogeneity, and long-term self-renewal of OCT4-positive colonies. 46 Ordered topographical features such as hexagonal (HEX) and honeycomb (HNY) pillars of 50–80 nm with a diameter of 30–40 nm fabricated in polystyrene with variable spacing, were used to study the effects of topography on ESC self-renewal. Growth on the HEX and HNY pillars was sufficient to maintain OCT4 expression and a higher proliferation rate, compared with flat surfaces without FGF-2 supplementation. 47
In another approach, silica colloidal crystals between 120 and 600 nm in diameter were able to improve both ESC self-maintenance and colony formation. 48 To assess if microtopographies can also guide self-maintenance of pluripotent cells, we used the TopoChip platform, containing 2176 unique topographical features at a constant 10 μm height profile. 49 Through machine learning approaches we observed that a low feature size was a major determinant for OCT4 expression in induced pluripotent stem cells (iPSCs), which also correlated with higher proliferation.
The BioSurface Structure Array (BSSA) is another microtopographical screening platform with square and round pillars on which 16 different combinations of lateral and gap dimensions were made. Together with a variation in height (0.6, 1.6, and 2.4 μm), a total of 504 topographical features were used to screen cellular responses, and it was found that ESC colony numbers increased with a decrease in pillar size. 50 The previous examples highlight the strength of high-throughput screening platforms, where through machine learning algorithms, the most optimal surface architecture can be found for either ESC phenotypic maintenance or differentiation. Figure 1 illustrates the diversity of the cellular response of MSCs and ESCs on the TopoChip platform (unpublished data).
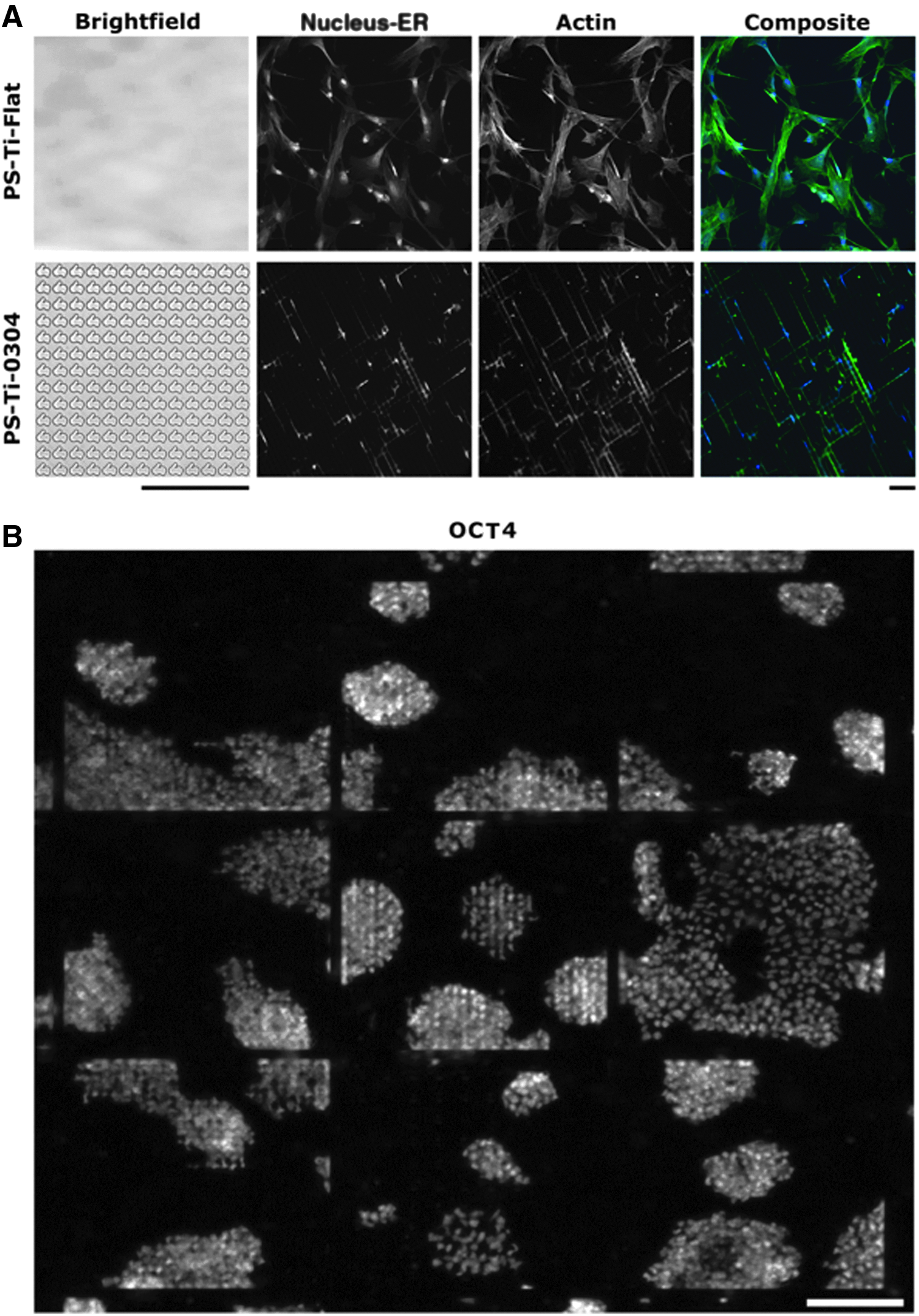
High-throughput screening tools can identify most optimal substratum properties for either ESC self-renewal or lineage differentiation.
As mentioned before, most research aimed at establishing lineage differentiation utilizing topographical features has been performed with MSCs. Still, the knowledge gained from these studies can be useful for protocols utilizing stromal progenitor cells derived from pluripotent cells,51,52 to treat for example cartilage and bone defects. Besides direct clinical applications, MSCs can also be used as an autologous cell source for supporting ESC self-renewal as a feeder layer. 53 To allow ESC differentiation toward MSCs without the need for chemical induction, square patterned nanopits were able to upregulate mesodermal marker expression and downregulate pluripotent, ectodermic, and endodermal markers. 54 Of note, these surfaces were the same topographical structures used to induce osteogenic differentiation of MSCs without the need for dexamethasone, 38 offering a promise for ESC differentiation for bone tissue engineering applications using only topographical cues.
A large body of research involving differentiation of pluripotent cells through topographies focuses on the generation of cells of the neuronal lineage. hESCs seeded on gelatin-coated nanoscale groove patterns (spacing = 350 nm; height = 500 nm) induced the expression of NEUROD1 among other neurogenic markers. 55 A similar observation was made when iPSCs were seeded on Matrigel-coated nanoscale grooved patterns (spacing = 350 nm; height = 300 nm) fabricated on PDMS. Expression of neurogenic markers, such as TUBB3, NEUROD1, and NEUROG1 was higher on these nanotopographical dimensions compared with flat, 2 or 5 μm spacing.55,56 In another study, Matrigel-coated polystyrene gratings of 2 μm width were able to efficiently induce neural differentiation in the absence of neurotrophic-inducing chemicals at later stages of the culture. 42
Besides improving differentiation efficiency, induction of neuronal subtypes can be achieved using isotropic surfaces. 57 Studies on 250 nm grooved topographies linked ESC actomyosin contractility with neuronal marker expression. 58 It is interesting to mention that nanotopographical grooves of 350 nm have also been previously used to induce MSC differentiation toward the neurogenic lineage. 59 To summarize, these studies reveal that both micro- and nanotopographical cues can play a key role in formulating more efficient and xenofree protocols for ESC differentiation toward the neurogenic lineages.
To investigate endoderm differentiation, tall nanopillars with different aspect ratios were used, through which stiffness parameters could be altered. It was found that tall pillars, mimicking a softer substrate, allowed a higher efficiency of endoderm differentiation after chemical stimulation. 60 In another approach, 4.5 nm diameter gold particles were combined with different chemistries to study ESC behavior, with all nanoparticle films inducing the expression of FOXA2, a marker for early endoderm commitment. Furthermore, this study provided evidence that ESCs can sense topographical cues smaller than 5 nm. 61 Although soluble growth factors are still required in these protocols, these studies indicate that biophysical cues can influence the endodermal differentiation of pluripotent stem cells.
Although this review highlights the influence of topographical cues on stem cells, research investigating differences in their response to topography of the two majorly studied pluripotent cell types (ESCs and iPSCs), is fairly limited. Research does indicate distinct gene expression profiles between iPSCs and ESCs, 62 which can lead to differences in differentiation efficiencies. 63 Furthermore, the cellular origin of iPSCs influences lineage differentiation propensity. 64 Therefore, the possibility arises that iPSC and ESC cell lines may have a variable response to topographical cues. In this context, it has been shown that different ESC lines respond slightly different on microtopographical cues in terms of self-renewal and differentiation. 50 Furthermore, the use of two different ESC lines show differences in marker expression when differentiated toward neuronal lineages on different topographical structures. 57 Recently, Abagnale et al. showed that submicrometer groove-ridge structures can modulate the shape of iPSC colonies, regulate cell polarity, and guide the orientation of actin fibers. 65
Altogether, these studies indicate that both ESCs and iPSCs respond to topographical cues, yet not necessarily in a completely similar manner. Future research, where the influence of topographical cues on both ESCs and iPSCs in the same experimental setting is investigated, might explore this observation more deeply.
Molecular Mechanism of Sensing the Underlying Substratum
The previous sections clearly demonstrate the influence of the ECM and topographical cues on ESC behavior and fate commitment. However, the exact molecular mechanisms that underlie these phenomena are unknown. In the following sections, we aim to summarize current knowledge and identify open questions.
Integrins and Downstream Kinases
A number of molecules sense changes in the underlying substratum and help convert this into a definitive transcriptional output that regulates cell fate. Integrins are one of the important substrate- and ECM-sensing proteins.66,67 Integrins are fundamental components of focal adhesions. These are heterodimeric receptors made up of one β subunit with one α subunit, clustered in different combinations in response to specific ECM proteins. 68 Integrins play a major role in mechanosensing, and sensing differences in ECM composition.69,70 Hayashi et al. demonstrate that the expression of integrins is dependent on the composition of the ECM. They further demonstrate that the overexpression of integrin subunits (specifically those subunits that are induced in ESCs upon culturing them on Type I collagen), result in the differentiation of mESCs, whereas inactivation of specific integrin subunits helps promote mESC self-renewal. 69 Furthermore, in the context of mesenchymal stem cells, it is known that the expression and clustering of integrin receptors changes in response to the stiffness of the underlying matrix. 71
Specific integrins were shown to be activated in response to different ranges of stiffness. In response to medium stiffness (10.2 kPa), MSCs differentiated into myocytes through β3 receptor-mediated signaling, whereas MSCs switched to ITGA2 in response to stiffer matrix (40.7 kPa) resulting in differentiation down the osteocyte lineage. This integrin switching in response to substrate stiffness also resulted in a change in the size of focal adhesions. 72
Interactions between integrins and the ECM result in a number of downstream signaling events, some of which involve the SRC family kinases. 73 Integrin-mediated focal adhesion kinase (FAK)-SRC signaling regulates cell adhesion dynamics by regulating the activity of the small GTPase, RHO. RHO functions as a major target molecule involved in mESC differentiation by activating downstream kinases such as RHO kinase (ROCK), resulting in cell spreading. 74 Significantly, the presence of a pharmacological inhibitor of ROCK not only blocked mESC spreading and differentiation and promoted colony formation, but also resulted in maintenance of OCT4 and NANOG expression even in the absence of LIF, although mESC numbers were lower than when cultured in the presence of LIF. 75 The function of RHO and ROCK in hESCs appears to depend heavily on the cellular context. Dissociated hESCs were susceptible to apoptosis due to actomyosin contractility, and this could be blocked in the presence of a ROCK inhibitor.76,77 Apoptosis was driven in these cells under conditions where RHO activity was high compared with RAC. 77 However, contrary to this, in intact hESC colonies, RHO was essential for the survival and propagation of hESCs. 78
The role of FAK in the maintenance of stem cell survival and pluripotency has been shown to vary between human and mouse ESCs. In fact, even within hESCs, the role of FAK remains controversial. Vitillo et al. report that integrin-associated FAK is active in hESCs, and that this signaling is important for protecting cells from apoptosis upon detachment. Furthermore, this signaling is required for the maintenance of the pluripotent state. 79 However, contrary to this, Villa-Diaz et al. report that FAK remains inactive in hESCs due to the expression of ITGA6. During the differentiation of hESCs, the levels of ITGA6 decrease and ITGB1 gets activated resulting in an activation of FAK, and differentiation of hESCs, 80 whereas specific combinations of integrins have been shown to maintain stemness in mESCs, even in the absence of LIF. 81 In mouse ESCs, FAK remains inactive and the expression of integrins is shown to undergo a switch in response to differentiation. 69
Some reports also show that FAK/SRC interacting partners may play an important role in integrin signaling. Knockout of one such interactor, Paxillin (Pxn), in ESCs shows delayed cell spreading, reduced FAK phosphorylation, and differentiation. 82 Interestingly, proteomic analysis comparing ESCs grown in serum with cells cultured in 2i (a defined medium containing inhibitors against GSK3β and MEK), showed higher levels of integrins and actin-binding proteins, such as talin, vinculin, and filamin in cells maintained in serum. 83 It is well known that serum contains many ECM components with prime differentiation, indicating that serum induces changes in the molecular ECM-sensing mechanism. Thus, the sensing of the ECM through integrins, coupled with the activation of specific downstream kinases and actin regulation may play a key role in maintaining specific cell fate (Fig. 2).
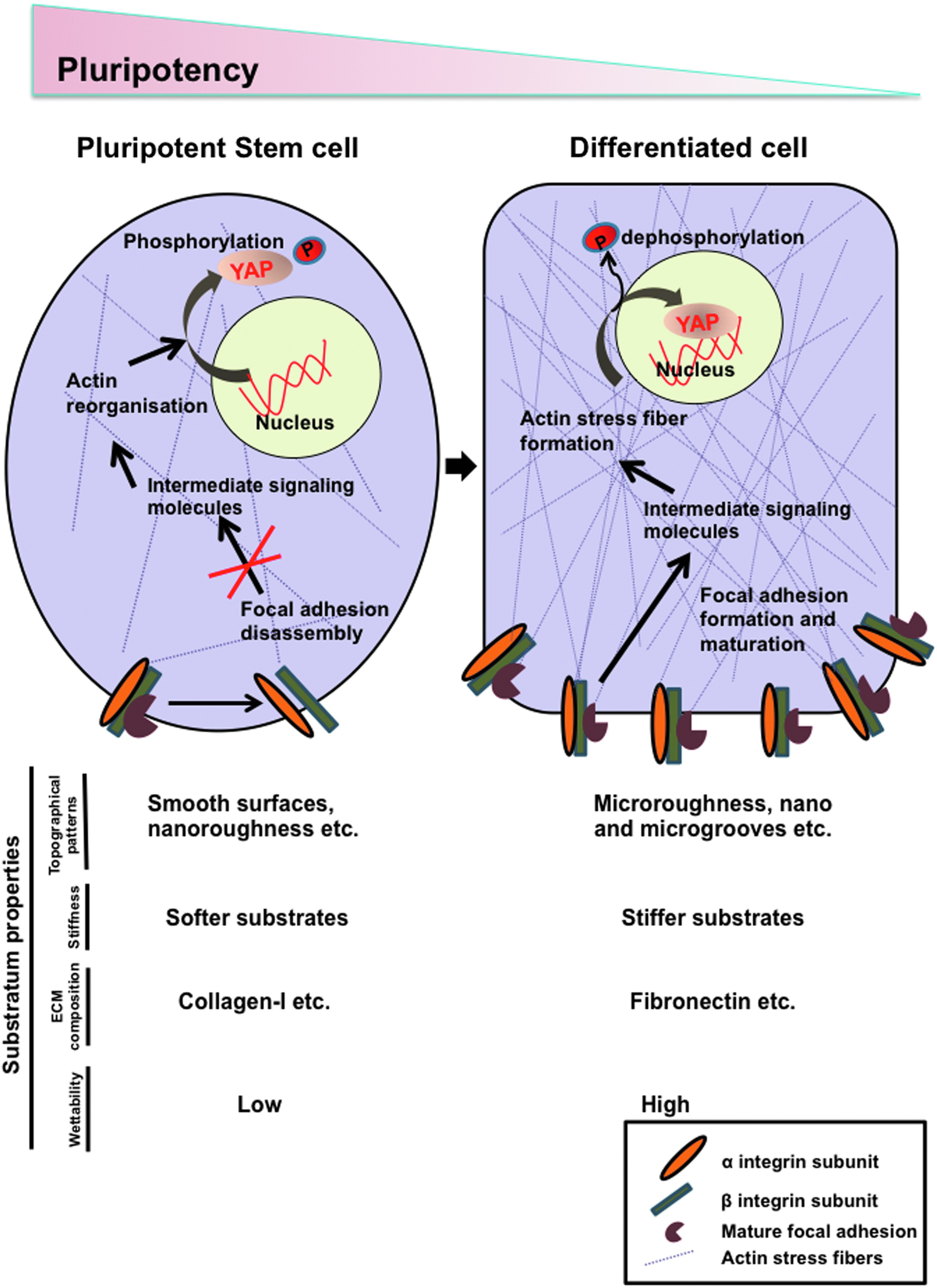
Molecular sensing of substratum properties by stem cells. Stem cells respond to the underlying substratum properties such as topography, stiffness, ECM composition, and wettability by changing the expression and engagement of integrins in a context-dependent manner. A change in the integrin composition affects the assembly and disassembly of focal adhesions and the recruitment of intermediate signaling molecules. These signals ultimately connect either directly or indirectly with the actin cytoskeleton and regulate the translocation of YAP/TAZ from the cytoplasm to the nucleus, resulting in a change in pluripotency.
In addition to integrins, mechanosensitive ion channels are also emerging as a novel player in sensing mechanical force and tension.84,85 A specific family of eukaryotic mechanosensitive channels, PIEZO, appears to play a role in maintaining homeostatic cell numbers in epithelia. 86 Interestingly, PIEZO1 activity was triggered by traction forces and played an important role in differentiation of neural stem cells to either an astrocytic or a neuronal lineage. 87 Thus, sensing of the external environment through the abovementioned molecules and mechanisms results in signal transduction within the stem cell for regulating cell fate choices.
Actin Dynamics and the Hippo Pathway
The mechanism of signal transduction within the stem cell requires a connection between the ECM and the nucleus. This is mediated by a complex set of interactions involving the integrins, actin cytoskeleton, actin-binding proteins, and other proteins that play a critical role in regulating downstream signaling. As mentioned above, the underlying substrate properties, such as stiffness and surface topography, lead to the activation of several kinases such as the SRC kinase, which further results in the activation of RHO. This further regulates the formation of F-actin, thus promoting actomyosin contractility, and leading to the translocation of YAP to the nucleus (Fig. 2). 88 YAP (Yorkie ortholog) is a part of the Hippo pathway that was first elucidated in Drosophila through genetic mosaic screens for tumor suppressors. 89 Briefly, upstream signals result in the activation of downstream kinases, such as LATS1/2, which is responsible for the phosphorylation of YAP. The phosphorylated form of YAP is sequestered in the cytoplasm, and is thus unable to activate downstream transcription. Hypophosphorylated forms of YAP/TAZ (Yorkie orthologs) are able to enter the nucleus and initiate transcription together with TEAD1-4, which contain DNA-binding domains. 89
YAP/TAZ has been implicated in playing a major role in sensing mechanical changes. Their phosphorylation status and subsequent shuttling between the nucleus and cytoplasm act to regulate gene expression changes in response to changes in substratum properties, such as substrate stiffness, topography, and cyclic stretching. 88 The Hippo pathway was shown to function as a barrier during the reprogramming of human somatic cells to induced pluripotency, as the knockdown of LATS2 enhanced the reprogramming efficiency. 90 In support of this, overexpression of YAP also promoted the conversion of hESCs to a naive state. 91 Conversely, in mESCs, loss of YAP could continue to support the pluripotent state, whereas YAP functioning was required for proper differentiation. 92
Actin and RHO-processing factors directly or indirectly regulate YAP/TAZ translocation by affecting the stress fiber stability in response to several physical and mechanical clues. 88 In intact hESCs, Rho and its activator, AKAP-Lbc, maintain the nuclear function of YAP/TAZ by regulating actin filament organization. 78 Interestingly, a different study demonstrated that the knockdown of specific kinases, including TESK1 or LIMK2 promoted the reprogramming of somatic cells to induced pluripotency by decreasing cofilin phosphorylation and disrupting actin filament structures. 93
Overall, the dynamic shuttling, transcriptional activity, and in some cases the expression of YAP/TAZ are tightly linked to matrix stiffness. 88 Within developing embryos as well, it has been shown that tensile forces generated by the actomyosin network are responsible for the positioning of cells and for embryo compaction, while also regulating the localization of YAP.94,95 YAP has also been shown to be involved in the maintenance of both naive as well as primed pluripotency, and for the regulation of ESC differentiation.78,91,92,96–98
In addition to the involvement of the Hippo pathway, the nucleus is also physically linked to the ECM due to the continuous network of intracellular and extracellular fibers. As such, topographical cues can directly affect the nucleus mechanically, resulting in altered gene expression through epigenetic modifications or chromatin and laminar reorganization. 98 The nuclear morphology of mesenchymal stem cells was more sensitive than differentiated cells to the architecture of nanofibrous scaffolds on which they were seeded. Application of tension to the scaffolds further caused changes in nuclear morphology of stem cells that were mediated by the actin cytoskeleton. 99 Kulangara et al. showed that mesenchymal stem cells cultured on 350 nm gratings showed a reduction in levels of LMNA and RB and a reduction in cell proliferation. 100 Changes in gene expression patterns were also observed in cells due to seeding on substrates with geometric patterning through actomyosin contractility. 101 Despite this wealth of literature, the precise mechanism connecting the ECM to the nucleus through the actin cytoskeleton, and alterations in gene expression still remains to be worked out in pluripotent stem cells.
The Influence of Topographical Cues on Integrin and Hippo Signaling
Cells are cultured in a multifactorial environment, which makes it difficult to fully assess the influence that topographical cues exert on stem cell fate. Utilizing materials with different stiffness, chemistry, surface coatings, and culture media can affect cell culture outcome and potentially mask topographical influences. This is further complicated by the interconnectedness of topographies with wettability and even stiffness, 102 both known to be major influencers in stem cell self-renewal and differentiation.103–105 Nevertheless, the influence of topographies can be independent from wettability as shown in the studies of Jaggy et al. and Jeon et al.45,46
Despite this environmental complexity, trends can be seen across multiple studies, for example, comparing different material roughness levels on stem cell maintenance shows that a smooth surface performs equal or better then nanoscale levels,42,44–46 and outperforms microscale roughness. 44 However, when combining both micro and nanoroughness, stem cell self-renewal was promoted, compared with a flat and nanorough surface. 46 On the nanoscale level, promotion of stem cell pluripotency is possible through the use of repetitive crystal structures 48 or pillars, 47 illustrating that besides size, other feature parameters such as symmetrical patterns can have an important effect on cell fate. Microtopographical cues can support a pluripotent phenotype, yet this is very dependent on feature parameters. In the study of Markert et al., higher features are supportive for self-renewal among the distance of the patterns, 50 whereas in the study of Reimer et al., a lower pattern density is associated with pluripotency. 49 The use of grooves or ridges in both micro and nanoscale dimensions are generally associated with promoting differentiation of pluripotent cells.55–58,65,106
In the previous sections, we emphasized the role of integrin signaling and specific topographical cues on ESC and iPSC cell fate. Activation of integrin signaling is associated with loss of pluripotency, 69 and topographies could influence this signaling through the focal adhesion kinase (FAK). 107 Cells grown on surfaces with low roughness promote pluripotency, show less focal adhesions, and reduced aligned actin stress fibers (Fig. 2). 47 Furthermore, increased phosphorylation of FAK, vinculin, and increased actomyosin contractility is associated with the induction of differentiation through the activation of its downstream signaling pathways.45,46,108–110 Topographical cues can also alter the spatial arrangement of FAKs and negatively influence pluripotency. 42 Furthermore, spreading of pluripotent cells is associated with increased integrin–matrix interactions and loss of self-renewal.69,111,112 The wettability of the substrate is associated with cell spreading and increased FAK formation,113,114 making this an important parameter in cell culture studies. Considering this, micro- and nanotopographical surfaces are known to influence adhesion characteristics of pluripotent stem cells.42,45,48,50,115
FAK signaling can regulate MSC differentiation as well, and proves to be important for differentiation toward the osteogenic lineage through the activation of canonical and noncanonical signaling pathways mediated by cytoskeleton reorganization,108–110,116 whereas a decrease in FAK presence is associated with a higher potential to differentiate toward the adipogenic lineage.107,117 Also in this context, differentiation can be mediated by nanotopographical cues.107,118
We previously elaborated on the role of actin–myosin dynamics and Hippo signaling in ESC pluripotency and differentiation. The downstream transcriptional co-activators, YAP and TAZ, are mechanosensitive barometers of the cell118,119 and are affected in MSCs by both surface stiffness and forced spreading, 88 influencing their differentiation capability. 120 The effect of these substrates and topographical cues on YAP and TAZ in ESCs and iPSCs has been less investigated. Topographical features are capable of enhancing the osteogenic differentiation of MSCs by increasing cell attachment, actin rearrangement, and promoting the activation of the YAP/TAZ signaling pathway. 121 Specific nanopatterns that are optimal for inducing osteogenic differentiation showed enhanced YAP activity when MSCs were cultured on such surfaces. 122 Nanotopographical features were also shown to promote the differentiation of hESCs and iPSCs into pancreatic endocrine cells 123 and neuronal cells, 124 through the regulation of the nucleocytoplasmic localization of YAP/TAZ.
In recent work by Abagnale et al., where nanoscale ridges support BMP4-induced differentiation of iPSCs, the spatial distribution of TAZ was colocalized with actin filaments and cell material adhesion sites when culturing iPSCs on nanogrooves. 65 These studies support the notion that YAP/TAZ pathways can play a role in transmitting topography-induced mechanical cues for pluripotent stem cells.
Relevance to Tissue Engineering and Regenerative Medicine
Human tissues are made up of many types of cells, which in turn interact with a variety of ECM proteins. The organization of the ECM in each tissue and organ varies from nano- to microscale proteins with different feature sizes. Micro- and nanoscale proteins from the ECM transmit chemical and physical cues to cells controlling various behaviors, including their adhesion, proliferation, migration, and differentiation. Therefore, topography-guided approaches that attempt to mimic the natural ECM has become a field of topical interest, with the aim of optimizing the generation of 3D artificial tissue grafts from stem cells.125–128
Substrates with topographical features can be beneficial in the field of tissue engineering, with noteworthy examples, including improving tissue–implant interaction,129,130 generation of cardiomyocytes from iPSCs, 131 improving the retention time of transplanted cells and their integration into the host tissue,132,133 generation of mature muscle patches, 134 and improving the process of skin regeneration. 135 Altogether, these reports strengthen the notion that topographical cues can help direct cell fate down a specific lineage and that patterned surfaces can be employed efficiently in the field of regenerative medicine.
Open Questions and Future Directions
During development, ESCs do not develop in an independent and isolated fashion. Instead, they respond to and interact with various biological, chemical, and physical cues. To further our fundamental understanding of embryonic development (and its relation to particular pathologies), as well as design improved in vitro culture methods to advance the field of regenerative medicine, more in-depth studies are needed to unravel the role of the ECM in ESC behavior. It is important to note that due to the complex interplay between ESCs and ECM and the large number of ways in which the ECM can be modified, combinatorial, high-throughput screening approaches, typically used for drug discovery in the pharmaceutical industry, will offer novel opportunities (defined as “materiomics”). 136 Recently, with the emergence of micro- and nanotechnologies, novel platforms have been developed to systematically probe the cell–substrate interface using cell size-limiting adhesive islands from fibronectin, 14 patterned surface pits, 15 patterned cell shapes, 101 or varying matrix elasticity or stiffness.11,137 Taking this a step further, others have developed platforms to independently and simultaneously manipulate two cell–ECM interfacial properties, such as cell size and cell shape, 13 or ECM stiffness and pore size, 138 and investigate their influence on cell behavior. Moreover, to systematically study large numbers of surfaces, the platforms mentioned above are being miniaturized, enabling high-throughput screening of surface topographies50,139 or chemistry.140,141
Altogether, with the development of combinatorial high-throughput techniques, developing advanced data mining techniques and system biological models are essential tools to identify patterns in the data, generate novel hypotheses, and to perform in silico experiments for a wide range of different conditions. Several examples in the literature exist such as (dynamic) models of gene regulatory networks that allow exploring reprogramming strategies,142–144 computational models of autocrine and paracrine signaling to determine the soluble factors essential for mESC survival in vitro, 145 and computational fluid dynamics models to optimize the culture conditions to promote efficient cardiogenesis of hydrogel-encapsulated ESCs in a rotating bioreactor. 146 For more detailed information on computational modeling, we refer the reader to the following excellent reviews.147–149
In summary, although it is clearly recognized that the ECM plays an important role during embryonic development, several outstanding questions remain. What are the molecular mechanisms that underlie ECM sensing and how do they interact? How do these (mechanobiological) pathways change during embryonic development and lineage specification? Are these pathways conserved among different species, and if so, what can we learn from this? Finally, how can we harness these mechanobiological cues to advance current scientific and clinical models of embryonic development?
Conclusions
A key component to successfully translating regenerative therapies from the bench to the bedside is an in-depth understanding of early embryonic development. In this review, we have detailed how mechanical properties of the ECM affect the pluripotency of the ESCs, in addition to a myriad of well-studied soluble factors, including growth factors and cytokines. It appears that the effect of mechanical cues (such as topography) that replicate the conditions within the developing embryo, favor ESC maintenance. Interestingly, although the influence of mechanical cues is well recognized, the exact underlying molecular mechanisms are poorly understood in the context of stem cells. However, as the field progresses through the development of new tools to study the cell–material interface, details of particular signal transduction pathways will emerge. As such, we believe that interdisciplinary collaborations of material scientists and developmental biologists will have great potential to illuminate the fundamental questions on ESC behavior and improve existing regenerative therapies.
Footnotes
Acknowledgments
The authors regret their inability to cite all the articles in the field due to space constraints. S.K. is a recipient of a Senior Research Fellowship from the Department of Biotechnology, India. J.d.B., A.C., B.V., and S.V. acknowledge the financial support of the Dutch province of Limburg. A.C. gratefully acknowledges the Dutch Science Foundation (NWO) for a VENI grant (15075), S.V. acknowledges the financial support of the European Union's Horizon 2020 Program (H2020-MSCA-ITN-2015; Grant agreement 676338). The authors thank members of the Subramanyam Laboratory and the de Boer group for critical comments.
Disclosure Statement
No competing financial interests.