
Abstract
In addition to mesenchymal stem cells, adipose-derived stem/stromal cells (ASCs) are an attractive source for a large variety of cell-based therapies. One of their most important potential applications is related to the regeneration of bone tissue thanks to their capacity to differentiate in bone cells. However, this requires a proper control of their osteogenic differentiation, which depends not only on the initial characteristics of harvested cells but also on the conditions used for their culture. In this review, we first briefly describe the preclinical and clinical trials using ASCs for bone regeneration and present the quantitative parameters used to characterize the osteogenic differentiation of ASCs. We then focus on the soluble factors influencing the osteogenic differentiation of ACS, including the steroid hormones and various growth factors, notably the most osteoinductive ones, the bone morphogenetic proteins (BMPs).
Impact statement
Adipose-derived stromal/stem cells are reviewed for their use in bone regeneration.
Introduction
Bone tissue engineering aims to induce regeneration of functional tissues using biomaterial constructs in combination with bioactive factors and/or stem cells 1 as an alternative to bone grafting. The use of stem cells is motivated by their active participation in native regenerative processes 2 and represents one of the main clinical strategies for bone regeneration. 3 Since their discovery, 4 adipose-derived stromal cells (ASCs) harvested from fat tissue, have been gaining increasing popularity in the field of regenerative medicine. The popularity of ASCs is based on a few inherent advantages that they possess over other types of adult stromal cells such as bone marrow-derived stromal cells (BMSCs), which are obtained from bone marrow aspirate.
For instance, ASCs can be obtained at high yield as they comprise up to 5% of the total number of nucleated cells in adipose tissues, compared with less than 0.01% for BMSCs in bone marrow. 5 The collection procedures for ASCs are relatively simple, painless, and minimize tissue morbidity. 6 Besides, ASCs maintain high proliferative potential during culture with average population doubling times of 2–3 days, 7 without genetic alteration or replicative senescence. 8 Even long-term cultures up to 20 passages do not induce significant chromosomal aberrations of ASCs and preserve their morphology, immunophenotype, normal division cycle, 9 as well as their apoptosis regulator function. 10 It was also shown that their long-term cryopreservation for more than 10 years does not impact their viability, differentiation potential, and immunophenotype characteristics. 11
Importantly, having demonstrated their capacity for multilineage differentiation, including adipogenic, chondrogenic, myogenic, and osteogenic lineages, 3 ASCs have been used in vivo to regenerate fat, 12 cartilage 13 and bone,14,15 and more complex tissues such as periodontium 16 and intervertebral disc. 17 Since they can also differentiate into endothelial cells, ASCs may serve as a single-cell source for the engineering of vascularized bone tissues,18,19 thus tackling a major bottleneck in the clinical translation of tissue engineering strategies. 3
Like other stromal cells, ASCs secrete several potent endogenous cytokines and immunomodulators 20 but at a comparatively higher rate, 21 thereby enhancing their paracrine function, most notably in angiogenic 19 and homing 22 activity toward endogenous progenitor cells in vivo.21,22 This paracrine function plays an important role in successful regenerative outcomes for bone and intervertebral disc regeneration.14,16,18 ASCs have been also shown to survive in hypoxic conditions, which makes them particularly suitable for surgical implantation 19 in oxygen-depleted environments. 23
The ability of ASCs to promote regeneration was originally considered to depend primarily on their osteogenic and chondrogenic differentiation, leading to the production of mature and functional bone cells and tissues. 24 However, their in vitro osteogenicity was demonstrated to be inferior in comparison with BMSCs. 19 While multiple studies have demonstrated bone differentiation of ASCs both in vitro25,26 and in vivo,27–29 there is a clear need to elucidate the basic mechanisms controlling this differentiation and to identify the factors and the experimental conditions influencing this process.
In this article, after a brief reviewing of the preclinical and clinical trials using ASCs in bone regeneration studies, we then focus on the soluble factors influencing the osteogenic differentiation of ACS, including the steroid hormones and various growth factors, including the most osteoinductive bone morphogenetic proteins (BMPs).
Preclinical and Clinical Trials Using Adipose-Derived Stem/Stromal Cells in Bone Regeneration
ASCs were studied in several preclinical animal models of bone regeneration. The most common application is related to the repair of critical size calvaria defects in rats,30–35 or rabbit, 36 where ASCs are used in association with different types of scaffolds, mostly made of synthetic polymers such as hydroxyapatite/poly(lactide-co-glycolide), 30 polycaprolactone, 31 poly(vinylidene-trifluoroethylene), 33 or bioceramics such as bioactive glasses, 32 membrane, calcium silicate, 34 and granulated bone substitutes, 35 In all the cases, a significant improvement of bone repair was observed. A few rat studies focused also on the repair of other bone fractures such as radii 37 where ASCs were used in combination with heterogeneous deproteinized bone and femur, 38 where they were used with tricalcium phosphate or a gelatin sponge.
Interestingly, ASCs implanted into a rat mandibular defect for 6 weeks, 39 led to visible bone tissue formation as early as 1 week and promoted faster and greater bone regeneration compared to the dental pulp stem cell group. Other studies focused on periodontal bone regeneration, which are reviewed in Dziedzic et al. 40
Miettinen and colleagues recently reviewed the perspectives for clinical translation of adipose stromal/stem cells. 41 They notably discussed the strength and weaknesses of the use of autologous versus allogenic ASCs for cell therapies. 41 Adipose stem cells were shown to be suitable candidates for allogenic cell therapies due to their low immunogenic profile. Thanks to their characteristics listed above (i.e., obtention at high yield, simplicity of isolation, efficient cryopreservation, multicellular differentiation, and secretion of cytokines and immunomodulators), ASCs are increasingly used in clinical trials for the regeneration of both soft42–44 and hard14,45 tissues, including bone tissue regeneration24,46,47: they are the most studied to treat disease related to the digestive system, followed by muscle, bone, and cartilage. 41 The team of Miettinen did the first study of good manufacturing practices compliant nonhematopoietic nonhematologic application for autologous ASCs in the treatment of hard-tissue defects at various sites of the craniomaxillofacial skeleton. 15 This case series assessed the results of 13 patients reconstructed with ASC-seeded resorbable scaffolds based on bioactive glass or β-tricalcium phosphate (β-TCP), at four very different anatomic sites within the craniomaxillofacial skeleton, namely frontal sinus (three cases), cranial bone (five cases), mandible (three cases), and nasal septum (two cases). The criterion for the success of the graft at these four implantation sites was whether the hard-tissue grafts in their recipient bed were functioning according to the demands of their new native sites during the follow-up period. 15
Globally, the results were good or fair, exept for the results of the cranial defects that were disappointing in three cases, as the authors found that the resorption of the constructs made of β-TCP was more important than expected. For these cranial constructs, it seems that the mesh layers must be made of rigid, nonabsorbable material (such as titanium), perhaps to sustain the constant dural pulsations to which these cranial wounds are exposed. 15
A search in the ClinicalTrials.gov registry (NIH) in August 2019 with keywords “adipose stem cell” and “bone [disease]” revealed eight registered trials targeting ASC-based bone regeneration and intervertebral disc treatment (Table 1). The clinical trials using ASCs are often comparing autologous or allogenic ASCs with mesenchymal stem cells (MSCs) harvested from different sources. In most studies, the stem cells were injected, either alone or in combination with a hydrogel. These trials can be classified according to their main application.
Summary of Registered Clinical Studies Found at ClinicalTrials.gov Using “Adipose Stem Cell and Bone [Disease] Not Withdrawn” Keywords
ASC, adipose-derived stromal cell; BMSC, bone marrow derived-stromal cell; MRI, magnetic resonance imaging; MSC, mesenchymal stem cell; N/A, not available.
Three studies aimed to treat fractures. This was achieved either by injections of ASCs and MSCs (NCT02307435, NCT02140528) obtained from bone marrow or umbilical cord, or by adding the stem cells in a composite graft made of hydroxyapatite granules in fibrin gels (NCT01532076).
Three studies focused on the stabilization of intervertebral disc degeneration by tissue grafting (NCT02529566), either by injection of cells within a hyaluronate gel (NCT02338271) or by direct injection of stem cells (NCT02097862), the results being published. 48 In this last study, Comella et al. compared the delivery of a stromal vascular fraction that contained ASCs and growth factors and that was obtained via a mini-lipoaspirate procedure of fat tissue, to the injection of platelet rich plasma that was obtained from peripheral blood. They evaluated the safety and efficacy of administering the vascular fraction and platelet rich plasma intradiscally into patients with degenerative disc disase. 48 Patients demonstrated statistically significant improvements in several parameters, including flexion and pain ratings. No severe adverse events were reported during a 12 month follow-up period with any incidence of infection or complication.
One study investigated the treatment of the avascular necrosis of the femoral head (NCT01643655).
One study aimed to heal spinal cord injury (NCT02981576).
Unfortunately, in all these studies, the detail of stem cell preparation before the injection is not described, nor is indicated the possible addition of soluble factors to prime the cells. Besides, it is currently not possible to conclude about the success of the majority of these trials due to the lack of information provided. In most cases, no result was reported for the completed studies and the terminated study (i.e., a study that was terminated earlier and for which the patients are no longer being examined or treated).
Osteogenic Differentiation of ASCs
As it is currently understood, the process of osteogenic differentiation of ASCs starts with the activation of osteogenic signaling pathways, usually triggered by the binding of specific factors to the complementary cell receptors. 20 It then proceeds in stages, followed first by the upregulation of osteogenic genes such as runt-related transcription factor 2 (RUNX2) and osterix (OSX) and simultaneous downregulation of genes associated with other lineages such as adipogenic peroxisome proliferator-activated receptor-γ (PPARγ) and CCAAT-enhancer-binding protein α (C/EBP-α). These variations in gene expression lead to the expression of relevant proteins, including alkaline phosphatase (ALP), the long-term deposition of extracellular matrix proteins, including osteopontin (OPN), osteonectin (OCN), and matrix mineralization. It is only when a combination of genes and proteins are expressed at a sufficiently high level in comparison to a negative control (i.e., a condition where cells are not expected to differentiate) that differentiation can be ascertained.
Up to now, two major osteogenic signaling pathways have been identified, which involve Smad and non-Smad proteins, 49 notably the mitogen-activated protein kinases (MAPK) that can be subdivided into three groups: the extracellular signal-regulated kinases (ERK), the jun N-terminal kinases (JNK), and the p38 MAPK. 50 BMPs, Notch, wingless/integrated (Wnt), and fibroblast growth factor (FGF) are involved in these different osteogenic signaling pathways. 51 Generally, osteoinductive factors by themselves are not sufficient and require supplementation by soluble factors or stimulation via external stimuli to reach the later stage of osteogenic differentiation (Fig. 1).
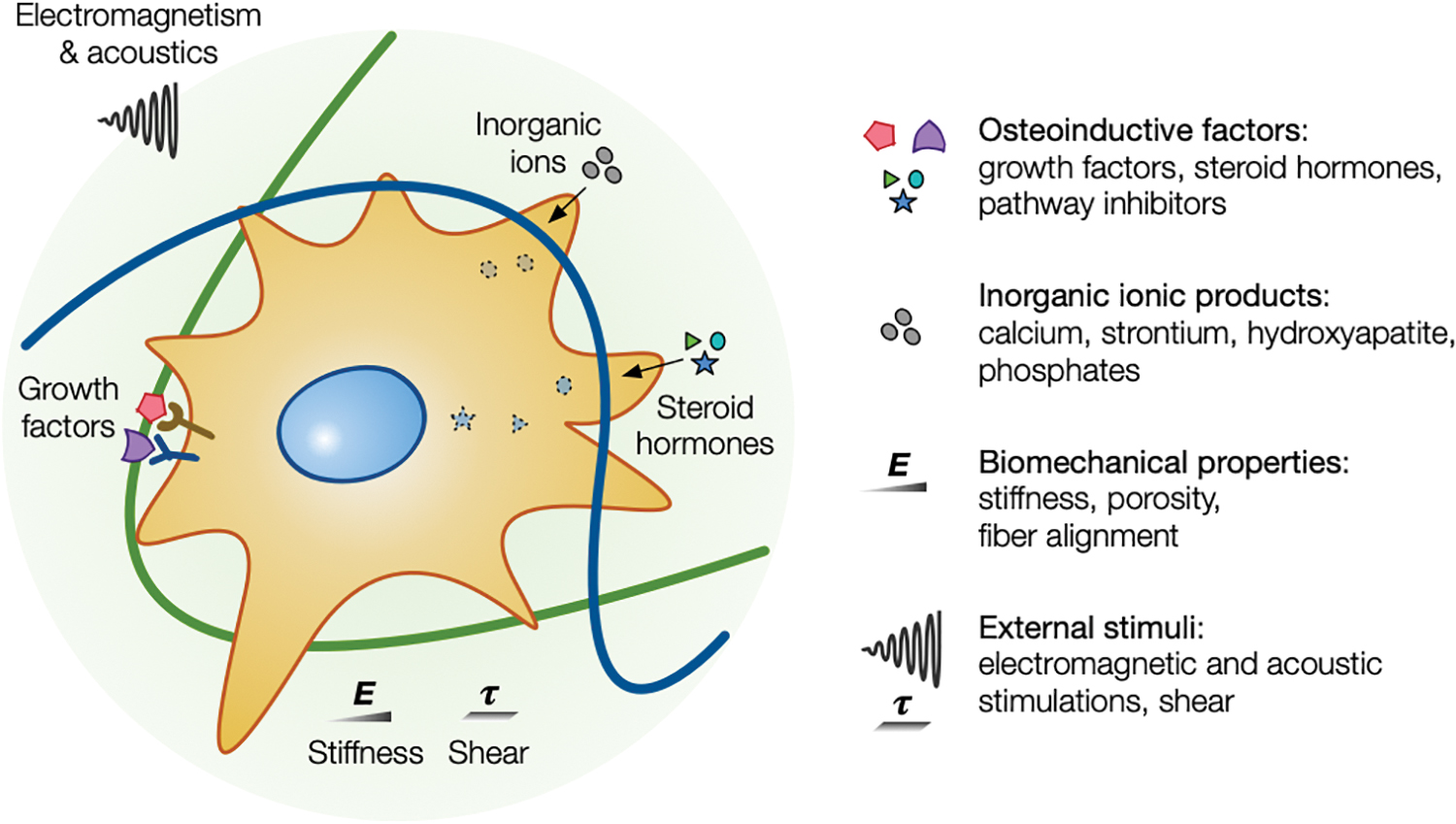
Schematic illustrating different physicochemical and biological cues that affect the osteogenic differentiation of ASCs. Various osteoinductive and supplementary factors and physical cues are known to affect the osteogenic differentiation of ASCs, including osteoinductive factors, inorganic ionic products, biomechanical properties of the cellular microenvironment, and external stimuli. ASC, adipose-derived stromal cell. Color images are available online.
Inorganic ionic products such as calcium,52,53 strontium, 54 or hydroxyapatite55,56 can also enhance the extent of the osteogenic differentiation of ASCs. Besides, the physicochemical properties of the extracellular matrix such as stiffness, 57 porosity, 58 fiber alignment, 59 and chemical composition60,61 are also playing an important role. In addition, external stimuli such as shear, 62 electromagnetic,63,64 and acoustic65,66 stimulations can also impact osteogenesis. All these stimuli likely activate different osteogenic signaling pathways. 67
Osteoinductive factors for the osteogenic differentiation of ASCs
The most common method to induce osteogenic differentiation of ASCs is to culture them in an osteogenic medium (OM) that is made by supplementing the growth medium (GM) with different soluble components (Fig. 2), including different types of sera, inductive factors, and sources of ascorbates and phosphates that are needed for matrix deposition and mineralization. 68
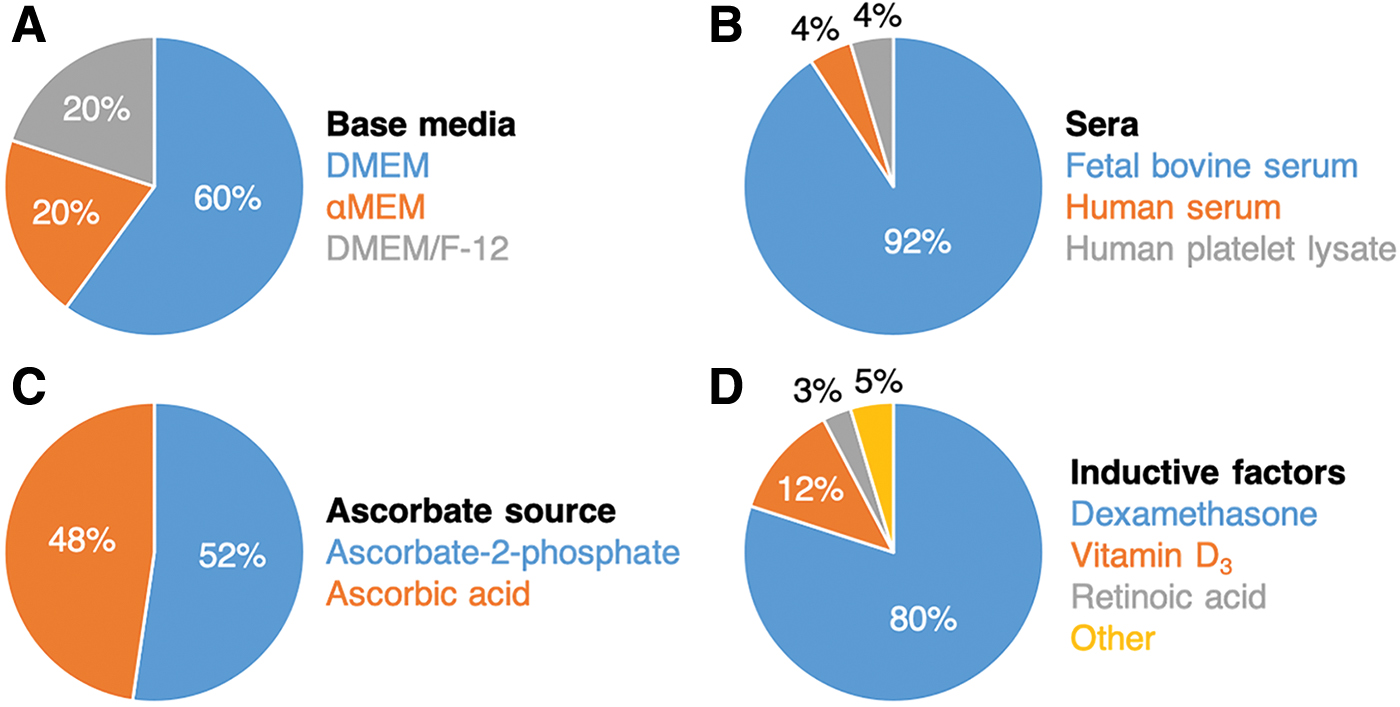
Summary of compositions of ASC OM. Main components of osteogenic media include the base medium
For ASCs, such media can be formulated using standard base media such as Dulbecco's modified Eagle's medium (DMEM), Minimum Essential Medium α (αMEM), or in combination with Ham's F-12 Nutrient Mix (DMEM/F-12). Among different available OM formulations, the most common one is based on DMEM with fetal bovine serum that is further supplemented with dexamethasone (DEX), a potent osteoinductive factor, and ascorbate-2-phosphate, and β-glycerophosphate (supplemented medium named hereafter SM).
Steroid hormones
Steroid hormones are the most prevalent soluble factors used for the osteogenic differentiation of ASCs. 69 This term regroups both naturally occurring and synthetic compounds, which share a common molecular structure of four hydrocarbon rings arranged in a specific manner. 70 Given their solubility in lipids, steroid hormones easily pass through the cellular membrane and bind to intracellular receptors located in the nucleus, cytosol, or plasma membrane. 71 They have wide-ranging functionalities and participate in cellular metabolism, development, and physiology by altering transcription levels of certain genes (known as genomic pathway) 72 or by interacting with a diverse set of targets like ion channels and membrane receptors (nongenomic pathway). 73 Three members of this family, namely DEX, vitamin D3 (VD3), and retinoic acid (RA) are potent osteoinductive factors in ASCs. 69
Dexamethasone
DEX is a soluble osteoinductive factor for ASCs, affecting a large variety of processes, including osteoblast proliferation, differentiation, and cell life span. 69 Initially reported by Zuk et al. for use in ASCs in view of its proven osteoinductive effect, 7 it was then shown to be osteoinductive in both in vitro74,75 and in vivo17,28 studies. DEX-differentiated ASCs were more efficient for bone regeneration than their nondifferentiated counterparts. 76
For a successful osteogenic differentiation, DEX must be administered continuously throughout the study, usually during medium changes operated every 2–3 days. 7 Several studies evidenced that DEX displayed greater osteoinductive capacity compared to other potent osteoinductive factors such as BMPs.77,78
It should be noted that the effect of DEX on ASCs is not exclusively osteoinductive as it is a crucial component in both adipogenic and chondrogenic differentiation media, although at different concentrations and in conjunction with other lineage-specific inductive factors. 24 Thus, its osteoinductive effect seems to be concentration-dependent. In most studies, DEX was added at 100 nM or below, 7 since several studies revealed that lower concentrations of 5 or 10 nM might lead to a faster and superior osteogenic differentiation.25,78 A higher concentration (1μM) led to the adipogenic differentiation of ASCs.4,7
To the best of our knowledge, there is no systematic study analyzing the effect of DEX concentration over a range of different medium formulations with various types and concentrations of sera and different concentrations of other supplements. Only one study investigated three differentiation medium formulations, in which DEX and L-ascorbate-2-phosphate (AP) concentrations were simultaneously varied. 78 Higher concentrations of DEX are normally used for adipogenic differentiation 79 and thereby inhibit osteogenic differentiation of ASCs. For instance, Liu et al. showed that blocking the osteogenesis-associated ERK pathway with a PD98059 chemical inhibitor in ASC cultures performed in OM supplemented with 100 nM DEX favored the adipogenic differentiation. 80 This indicates that a DEX concentration of 100 nM might be inducing both differentiation pathways and needs further induction for the commitment to the osteogenic lineage. One study even reported the detection of triglycerides in ASC lysates cultured in OM supplemented with 100 nM DEX. 60
Several different signaling pathways appear to be implicated in DEX-mediated osteogenic differentiation in ASCs, especially the MAPK signaling pathway. Inhibiting one of the proteins involved in this pathway greatly compromised osteogenic differentiation of ASCs in favor of their adipogenic differentiation. This is potentially due to the interruption of MAPK-dependent phosphorylation of PPARγ. 80 Activating the ERK pathway with a chemical activator AR-A014418 in ASCs cultured in OM supplemented with DEX led to the upregulation of RUNX2, ALP, and OCN, an increase of ALP activity, and matrix mineralization. 81 BMP signaling pathway is also involved since the suppression of noggin, an antagonist of BMPs, led to the upregulation of RUNX2, ALP, OSX, and OCN in ASCs cultured in OM supplemented with DEX. 82 The upregulation of SMAD1, a protein involved in BMP signaling pathway, further enhanced DEX-induced ASC osteogenic differentiation. 83 Several other studies have detected the endogenous expression of BMP-4 and BMP-6 in OM. 84 The Wnt signaling pathway is another potential contributor to DEX-induced osteogenic differentiation since its disruption via viral knockdown of ARL4C led to downregulation of RUNX2, ALP, OCN, and reduced matrix mineralization. 84 However, its activation via supplementation with WNT3A protein led to the increase in ALP activity but no significant upregulation of the osteogenic genes. 85
In constrast, other studies questionned the utility of DEX for achieving the osteogenic differentiation of ASCs.4,69 Malladi et al. found notably that 10 nM DEX by itself and in combination with either VD3 or RA consistently upregulated adipogenic PPARγ and downregulated OPN and OCN. 69 Others reported similar downregulation of OCN in ASCs cultured in OM supplemented with DEX compared to their nonsupplemented counterparts. 60 Interestingly, several reports have indicated that ASCs can undergo osteogenic differentiation when cultured in SM without the need for DEX.6,86,87
Vitamin D3
Like DEX, VD3 serves multiple vital cellular functions, the most notable being the maintenance of calcium and phosphorus levels in the cytosol. 58 Present in virtually every cell in the human body, it is implicated in the synthesis of hundreds of important proteins via binding to the vitamin D nuclear receptors. 88 In certain osteoblasts and hematopoietic cells, it inhibits cellular proliferation and induces cell differentiation. 69 Initially introduced by Zuk et al., who sought a more effective replacement for DEX, 5 VD3-treated ASCs were found to display significantly higher levels of ALP activity compared to DEX-treated groups and the required concentration of VD3 for osteogenic differentiation was 10 times lower than the DEX one (10 nM VD3 vs. 100 nM DEX).
Other comparative studies also pointed to their superior osteoinductive potential over DEX.80,89 In vitro studies suggest that VD3-induced osteogenic differentiation is dose-dependent 69 with an increase of ALP activity and matrix mineralization at 1 μM. 90 Due to its effectiveness, VD3 has been combined with DEX90,91 and BMP-284 to synergistically improve the extent of ASC osteogenic differentiation. Interestingly, in contrast to DEX, VD3 seems to induce the endogenous expression of BMP-2. This suggests that BMP signaling pathway is involved in VD3-meadiated osteogenic differentiation of ASCs. 89
Retinoic acid
Similar to DEX and VD3, main biological targets of RA are genes, whose expression is modulated by binding RA to two types of nuclear receptors, the retinoic acid and the retinoid X ones. 69 RA plays a critical role in embryological development and supports the functional viability of adult organs. 92 The use of retinoic acid for ASC osteogenic differentiation is much less common compared to DEX or VD3. Some studies demonstrated that RA enhances the osteogenic differentiation of ASCs in a dose-dependent manner, but the doses used were much higher compared to DEX and VD3, in the range of 1–5 μM versus standard 1–100 nM for DEX and VD3.69,93 Other studies found that RA can synergistically stimulate the osteogenic differentiation when coadministered with BMP-2,93,94 in part, due to its inhibitory effect on the adipogenic differentiation. 93
Bone Morphogenetic Proteins
Overview of bone morphogenetic proteins and their receptors
BMPs are an important group of growth factors belonging to the transforming growth factor β (TGF-β) superfamily, 95 with a diverse set of embryological and physiological functions. 96 BMPs have been shown to play key roles in overall tissue architecture and homeostasis 97 and are now considered as “body morphogenetic proteins.” 95 BMP signaling is mediated via BMP receptors of type I and type II.87,98 There is a total of four type I receptors: Activin receptor-like kinase I (ALK1), Activin receptor type I (ALK2), and BMP receptor type IA (ALK3) and IB (ALK6); and three type II receptors: BMP receptor type II (BMPR2) and activin receptors type IIA (ACTVR2A) and IIB (ACVR2B). Binding of BMPs to their receptors results in the activation of the SMAD-dependent and MAPK (ERK, p38, JNK) pathways. 84
Several studies have reported that ASCs may have a heterogeneous population of BMP receptors. 99 Generally, ASCs do express the majority of these receptors, 94 including ALK1, 100 ALK2, 101 ALK3, 102 ALK6, 99 and BMPR2. 103 Interestingly, osteogenic differentiation of ASCs can itself lead to their upregulation, as observed for ALK694,104 and BMPR2, 103 or downregulation, as observed for ALK1. It is worth mentioning that BMPs bind to different receptors with different affinities. 105 Thus, the BMP-response of ASCs might be lineage-specific. 94 Studies have shown that BMP signaling through ALK6 seems to preferentially promote osteogenesis, 99 while signaling through ALK3 rather promotes dipogenesis. 104
In view of their clinical approval,106–108 many studies have targeted BMP-2 and BMP-7 for the osteogenic differentiation of ASCs.77,78,109–111 However, it remains to be seen whether they are indeed the most osteogenic ones. A comparative study of the osteoinductive capacity of six different BMPs on ASCs found that BMP-6 and BMP-9 are the most efficient to enhance the ALP activity. 112 Moreover, other studies have shown poor osteoinductive performances for both BMP-298 and BMP-7. 78 It is presumed that BMP-2, BMP-4, BMP-6, BMP-7, and BMP-9 might be the most osteoinductive ones among the BMP family,113–115 although studies on their osteogenic effects on ASCs are still largely lacking. 84 Some studies have found that the timing of the administration also had an important influence with delayed administrations being more efficient in driving the differentiation. 90
Moreover, BMP heterodimers seem to be more potently osteoinductive compared to homodimers, with BMP-2/6, BMP-2/7, and BMP-4/7 promoting much higher ALP activity than their homodimer counterparts.20,111
Bone morphogenetic protein-2
BMP-2 is by far the most targeted BMP for the induction of osteogenic differentiation in ASCs.
84
Despite initially promising results, especially with transfected ASCs for their endogenous expression,29,116 whether exogeneous BMP-2 is a viable osteoinductive factor for ASCs is still debated.
96
Published research on this topic can be grouped into two main categories (Table 2):
Studies showing the osteoinductive effect of BMP-2, although not for terminal commitment of ASCs to an osteogenic lineage; Studies showing its inefficacy and even inhibitory effects.
Summary of the Effects of Different Bone Morphogenetic Proteins on the Osteogenic Differentiation of Adipose-Derived Stem/Stromal Cells
More details on these effects are provided in respective sections.
ALP, alkaline phosphatase;
The apparent inconsistencies between these studies are likely due to the intrinsic variations in ASC characteristics, including the aforementioned differential expression of the BMP receptors, 99 differences in experimental designs, medium supplements, and their sources. 77 It is, however, evident that the BMP-2 signaling is a complex interplay between different actuators and inhibitors and needs further and more complete elucidation.
BMP-2 seems to have an osteoinductive effect on ASCs when it comes to the expression of osteogenic genes and ALP activity. In ASCs cultured in GM, 500 ng/mL of BMP-2 significantly increased ALP activity 112 and the expression of osteogenic genes ALP, OCN, and OPN. 117 When cultured in SM, both preliminary 15 min treatment with 10 ng/mL of BMP-2 and its continuous supplementation at 100 and 300 ng/mL throughout the duration of culture 118 were sufficient to increase the ALP activity.
At the same time, BMP-2 alone may be insufficient to drive the long-term osteogenic differentiation of ASCs, that is, matrix mineralization. For example, for ASCs cultured in SM, increasing BMP-2 concentrations of up to 200 ng/mL showed a dose-dependent increase of ALP activity, but failed to induce matrix mineralization. 93 In another study, the addition of 500 ng/mL of BMP-2 to GM was found to upregulate TGF-β1, RUNX2, and alpha-1 type I collagen (COL1A1) in ASCs, but led to neither a statistically significant increase of ALP activity nor matrix mineralization. 119 Similarly, BMP-2 at concentrations from 25 to 100 ng/mL induced a dose-dependent activation of SMAD signaling in ASCs cultured in GM, but failed to increase either the ALP activity or matrix mineralization in these conditions. 98
Matrix mineralization seems to require the supplementation with other osteoinductive factors such as RA or DEX. For instance, it was shown that the addition of RA in SM promoted matrix mineralization. 94 This result can be attributed to the synergistic suppression of adipogenic differentiation. 93 This was corroborated by other studies showing a dose-dependent increase of ALP activity and matrix mineralization in ASCs cultured in OM supplemented with either 10–100 ng/mL6,81,120 or 5–200 ng/mL of BMP-2. 111 The expression of RUNX2, ALP, and OCN was also upregulated, but not in a linear dose-dependent manner. Addition of BMP-2 at high concentration of 1 μg/mL also showed synergistic improvement of the matrix mineralization. 116
In contrast, the following cohort of studies suggests that BMP-2 has either no effect on such differentiation or may in fact actively inhibit it. A previously mentioned study by Zuk et al. showed the lack of SMAD phosphorylation and its nuclear translocation in ASCs treated with BMP-2. 6 In ASCs cultured in OM, matrix mineralization and the expression of RUNX2, OCN, OSN, OPN, and ALP were highly inconsistent depending on the source of the BMP-2. Surprisingly, both OM- and BMP-2-treated ASCs failed to express distal-less 3 (DLX3), distal-less 5 (DLX5), and OSX, three important target genes in the BMP signaling pathway.
Lee et al., suggested that BMP-2 actively inhibits the osteogenic differentiation of ASCs by inducing their adipogenic differentiation instead. 121 Even though these ASCs tested positively for the presence of BMP receptors and BMP-responsive genes like DLX3 and inhibitor of DNA-binding 2 (ID2), the addition of BMP-2 at 50 and 100 ng/mL in both GM and OM led to the formation of lipid droplets and the upregulation of adipogenic genes C/EBP-α and PPARγ. Matrix mineralization was inhibited in six out of the eight different ASC studied lots. The induction of adipogenic differentiation in BMP-2-supplemented OM was corroborated by another study showing increased expression of adipogenic protein 2 (AP2), 98 this effect being highly donor-dependent. A similar absence of BMP-2 osteoinductivity was reported in the case of ASCs treated with 50 ng/mL of BMP-2 during 2 weeks in both GM and OM. 77 Hutton et al. found that the supplementation of OM with 20 ng/mL of BMP-2 upregulated the expression of OCN but did not result in greater matrix mineralization compared to OM alone. 87
Bone morphogenetic protein-4
There are only a few studies concerning the use of BMP-4 for the osteogenic differentiation of ASCs. At very low concentrations of 0.01–0.1 ng/mL, BMP-4 was shown to aid in the survival of ASCs and the maintenance of their stemness, 122 while at 100 ng/mL, it increased both ALP activity and matrix mineralization. 123 Brennan et al. found that adding 50 ng/mL of BMP-4 in SM caused upregulated expressions of RUNX2, ALP, OSX, DLX5, and OSC in vitro, 19 but these primed ASCs showed minor improvement of the osteoforming ability in vivo.
It was also reported that 10 ng/mL of BMP-4 can significantly increase the ALP activity, although its increasing concentrations of 50 and 100 ng/mL seemed to drastically reduce it. 112 Only few studies deal with BMP-4-transduced ASCs for their osteogenic differentiation, 124 but the lack of other studies with BMP-4 belies the fact that its function in ASCs is likely not exclusively osteogenic. For example, low concentrations of BMP-4 of about 0.01–0.1 ng/mL seem to increase cell number and prevent apoptosis during ASC culture without adverse effects on either of their osteogenic or adipogenic capacities. 122 Others found that BMP-4 might induce adipogenesis in ASCs 125 or even their differentiation into smooth muscle cells. 126
Bone morphogenetic protein-6
BMP-6 appears to induce both osteogenic and chondrogenic differentiation of ASCs. The ALP activity increased for ASCs cultured in GM with increasing concentrations of BMP-6 from 10 to 500 ng/mL, even though no linear dose-dependence was apparent. 112 Another study using BMP-6 at concentrations between 0 and 200 ng/mL in SM found osteogenic or chondrogenic differentiation depending on the dose and culture conditions 127 : there was a dose-dependent increase of osteogenic genes (RUNX2 and OCN), ALP activity, and matrix mineralization for monolayer ASC cultures, while BMP-6 pellet cultures resulted in chondrogenic genes (sex-determining region Y)-box 9 (SOX9) and aggrecan (AGC) upregulation and proteoglycan accumulation. Zhang et al. showed that BMP-6 along with vascular endothelial growth factor (VEGF) may synergistically enhance the osteogenic differentiation of ASCs cultured in OM as opposed to BMP-6 alone. 100 The same group further investigated the molecular mechanisms of the VEGF/BMP-6 crosstalk. 128 This study showed the upregulation of RUNX2, OSX, and DLX5 when OM was supplemented with 1 ng/mL of BMP-6 without difference in matrix mineralization compared to OM alone. In constrast, a study by Kyllönen et al. used 100 ng/mL of BMP-6 in combination with OM and found that OM alone resulted in stronger ALP activity and higher matrix mineralization than in its combination with BMP-6. 78 Similar results were found when GM was supplemented with BMP-6.
Bone morphogenetic protein-7
The use of BMP-7 for osteogenic differentiation of ASCs is less common compared to other members of the BMP family as BMP-7 seems to be an important inductive factor for their chondrogenic differentiation. 129 A study by Tirkkonen et al. found that the addition of 100 ng/mL of BMP-7 to GM resulted in significant reduction of cell proliferation and ALP activity, compared to both GM and OM without the BMP-7 supplementation. 77 In OM, the addition of BMP-7 did not result in either significant increase or decrease of ALP activity. Similarly, the addition of 100 ng/mL of BMP-7 in OM, even in combination with BMP-6 and VEGF, lowered both ALP activity and matrix mineralization compared to OM alone. 78 Adding BMP-7 with BMP-6 and VEGF also failed to induce any significant effect on the ALP activity and matrix mineralization. A study by Rivera et al. showed the decrease of the ALP activity of ASCs cultured in GM for all concentrations studied, from 10 up to 500 ng/mL. 112 On the contrary, Zhang et al. reported that the addition of BMP-7 in OM induced a dose-dependent increase of ALP activity as well as the expression of RUNX2, OPN, COL1A1, and OSC within the concentration range from 5 to 200 ng/mL, compared to OM. 111 Furthermore, the addition of 200 ng/mL of BMP-7 in OM resulted in significantly enhanced matrix mineralization. However, in GM, BMP-7 seemed to have no effect. Interestingly, the authors found similar effects for BMP-2/7 heterodimer that promoted the osteogenic differentiation of ASCs via the Wnt signaling pathway and not through the canonical BMP signaling pathway.
Bone morphogenetic protein-9
The interest in the use of BMP-9 as an osteoinductive factor for ASCs is rather recent. 130 BMP-9 is considered to be one of the most osteogenic BMPs 92 and has been shown to induce strong osteogenic differentiation both in vitro and in vivo when endogeneously expressed in ASCs. 130 A comparative study of different BMPs on ALP activity evidenced that BMP-9 was the most effective at concentrations higher than 100 ng/mL. The concentrations below 100 ng/mL did not affect ALP activity and no apparent dose dependency was observed. 98 Yuan et al. showed that increasing concentrations of BMP-9 from 2.5 up to 10 μg/mL upregulated the expression of both ALP and RUNX2 in a dose-dependent manner. 131 The study also identified the synergistic cooperation between BMP-9 and focal adhesion kinases, which promoted the osteogenic differentiation of ASCs via the Wnt signaling pathway and not through the canonical BMP signaling pathway.
Matrix mineralization was similarly enhanced when ASCs were exposed to BMP-9. BMP-9 interacts mainly via ALK1132 and, in conjunction with ALK2, mediates its osteogenic signaling. 92 Endoglin, a type I membrane glycoprotein also known as CD105, was recently identified as partner in BMP-9 signaling 132 and involved in enhanced matrix mineralization. Interestingly, BMP-9 may induce the differentiation in brown adipocytes thus having antiobesity capacity. 132
Other Growth Factors
While BMPs are the most potent osteogenic growth factors, other growth factors such as FGF, platelet-derived growth factor (PDGF), and VEGF may participate in the osteogenic differentiation of ASCs. 20 Their effect on the osteogenic differentiation of ASCs is summarized in Table 3.
Summary of the Effects of Platelet-Derived Growth Factor, Vascular Endothelial Growth Factor and Fibroblast Growth Factor on the Osteogenic Differentiation of Adipose-Derived Stem/Stromal Cells
More details are given in the main text.
FGF, fibroblast growth factor; PDGF, platelet-derived growth factor; VEGF, vascular endothelial growth factor.
Fibroblast growth factor
FGF plays a role in several important cellular functions such as mitogenesis and angiogenesis. 20 It was found to be involved in all stages of endochondral bone formation and skeletal development. It is sometimes supplemented in culture media for ASCs, both in osteogenic 133 and neurogenic 7 formulations. Furthermore, its upregulation in spheroid cultures of ASCs compared to ASC monolayer was shown to be critical for in vitro angiogenesis 134 ; however, studies with murine ASCs evidenced that FGF actively inhibits the osteogenic differentiation of ASCs cultured in RA-supplemented SM. 135 This inhibition was dose-dependent between 0.2 and 20 ng/mL regarding ALP activity and matrix mineralization. The effect was not found for hASCs, for which FGF showed no inhibitory effect on matrix mineralization.
Platelet-derived growth factor
PDGF is involved in a variety of cellular functions, most notably in mitogenesis, chemotaxis, 20 and angiogenesis 136 to form stable and mature vascular networks. 130 Like BMP-2 and BMP-7, the therapeutic use of PDGF-BB is regulatory approved, notably to treat ankle fusion and distal radius fractures. 3 The osteoinductivity of PDGF in ASCs cultured in SM was confirmed by dose-dependent increase of matrix mineralization at relatively low concentrations of 2 and 20 ng/mL. 87 At 20 ng/mL, it upregulated OCN to the same extent as BMP-2, but resulted in a 5.6-fold greater matrix mineralization. Despite these effects, PDGF does not seem to be essential for the osteogenic differentiation of ASCs as the administration of its inhibitor, AG1295, and did not fully inhibit matrix mineralization. The promotive effect of PDGF was further confirmed by Hung et al. who found that the addition of PDGF (20 ng/mL) in SM markedly enhanced matrix mineralization and upregulated RUNX2 and OCN. 86
Vascular endothelial growth factor
VEGF is involved in vasculogenesis 137 and is of particular interest in bone tissue engineering since it is present in native bone matrices and allows for the development of vascularized, thus viable, bone tissues. 138 Whether VEGF has an osteogenic capacity in ASCs is still debated. VEGF was found to induce a dose-dependent increase of the ALP activity and matrix mineralization when supplemented in SM at 20 and 200 ng/mL. 139 RUNX2 and OSC as well as angiogenic platelet endothelial cell adhesion molecule-1 and vascular endothelial cadherin (VEC) genes were also upregulated. These in vitro results were confirmed in vivo since ASCs treated with VEGF showed a significantly higher capacity to form bone than separate delivery of ASCs or VEGF. Even at low concentrations of 0.1, 1, and 10 ng/mL, VEGF seemed to dose dependently increase the matrix mineralization when supplemented in OM. 100 The expression of RUNX2, ALP, OSC, and OST was also upregulated when OM was supplemented with VEGF, although no clear dose dependency was observed. This positive effect on the osteogenic differentiation was further improved when VEGF was combined with BMP-6 at 1 ng/mL, presumably due to the synergistic cross talk between VEGF and BMP-6 signaling pathways. Another study also showed that the addition of 10 ng/mL of VEGF in OM significantly promoted upregulation of COL1A2, OST, RUNX2, and DXL5 and matrix mineralization compared to both GM and OM. 128 The authors also found that the cross talk between VEGF and BMP-6 activated the p38MAPK pathway and inhibited the activation of protein kinase B (AKT), but did not affect the phosphorylation of neither SMAD nor ERK. Valenzuela et al. 140 cocultured ASCs with osteoprogenitor and human umbilical vascular endothelial cells (HUVECs) at varying degrees of differentiation. They found that ASCs cocultured with HUVECs had the highest bone-forming capacity of all the combinations tested, which corroborates the above cited in vivo study that ASCs treated with VEGF showed a significantly higher capacity to form bone. 100 This suggests that the formation of a vascular network and its associated secreted factors (e.g., VEGF) may help to push ASCs along the osteoblastic differentiation pathway, recapitulating natural osteogenesis in vivo.
In constrast, a study by Kyllönen et al. using VEGF at 20 ng/mL along with 100 ng/mL of BMP-6 and BMP-7 showed that it did not improve ALP activity nor matrix mineralization compared to GM. 78 It remained ineffective when supplemented in OM as well. Overall, OM alone was superior to any growth factor supplementation in inducing the osteogenic differentiation of ASCs. Besides, supplementing VEGF into GM and OM did not result in marked improvement of the osteogenic differentiation as the levels of ALP activity, and matrix deposition remained unchanged in both media. 77
Conclusions
ASCs emerged as an alternative source of stem cells for therapy, instead of mesenchymal stromal cells. They are notably used for bone regeneration as evidenced by different studies performed at the preclinical level and in some clinical trials. Although the results of these clinical trials are poorly accessible because they have not yet been published, it is critical now to better control the osteogenic differentiation of ASCs to extend their use in bone tissue engineering applications. In this context, this review provided an overview on the influence of soluble factors such as steroid hormones and growth factors on the osteogenic differentiation of ASCs. If used solely or in a proper combination, and in the good range of concentration, these soluble factors allow promoting the osteogenic differentiation of ASCs and consequently the formation of bone tissue. In particular, BMPs appear to play an important role on adipose cell differentiation, driving them to bone, cartilage, or fat depending on the BMP type and concentration used. The steroid hormones can also be used to guide ASCs differentiation, DEX being the most often used one and VD3 being the most efficient, both having dose-dependent effects.
However, the harvesting and culture conditions as well as the initial characteristics of the stem cells are also important parameters, which determine the cellular behavior. Therefore, in the future, it would be important to provide a standard for the characterization and the culture of ASCs to be able to directly compare the results obtained from different studies. Such an approach is critical to extend the therapeutic use of ASCs for bone regeneration.
Footnotes
Disclosure Statement
There is no competing financial interest.
Funding Information
The work was supported by the Marie Skłodowska-Curie Innovative Training Network program (ITN-EJD) of the EU (European Joint Doctorate in Functional Materials Research; grant no: 641640). K. Glinel is a senior research associate of the F.R.S.–FNRS. C. Picart is a senior member of the Institut Universitaire de France, whose financial support is acknowledged. This work was funded by Fondation Recherche Médicale (DEQ20170336746), by the ANR (CO-DECIDE, grant ANR-17-CE13–022) and by the European Research Council (ERC POC BioactiveCoating, GA692924).