
Abstract
Collagen is a structural protein that is widely present in vertebrates, being usually distributed in tissues in the form of fibers. In living organisms, fibers are organized in different orientations in various tissues. As the structural base in connective tissue and load-bearing tissue, the orientation of collagen fibers plays an extremely important role in the mechanical properties and physiological and biochemical functions. The study on mechanics role in formation of oriented collagen fibers enables us to understand how discrete cells use limited molecular materials to create tissues with different structures, thereby promoting our understanding of the mechanism of tissue formation from scratch, from invisible to tangible. However, the current understanding of the mechanism of fiber orientation is still insufficient. In addition, existing fabrication methods of oriented fibers are varied and involve interdisciplinary study, and the achievements of each experiment are favorable to the construction and improvement of the fiber orientation theory. To this end, this review focuses on the preparation methods of oriented fibers and proposes a model explaining the formation process of oriented fibers in tendons based on the existing fiber theory.
Impact statement
As the structural base in connective tissue and load-bearing tissue, the orientation of collagen fibers plays an extremely important role in the mechanical properties and physiological and biochemical functions. However, the current understanding of the mechanism of fiber orientation is still insufficient, which is greatly responsible for the challenge of functional tissue repair and regeneration. Understanding the mechanism of fiber orientation can promote the successful application of fiber orientation scaffolds in tissue repair and regeneration, as well as providing an insight for the mechanism of tissue histomorphology.
Introduction
Collagen is a structural protein widely distributed in vertebrates, accounting for about 30% of the total protein of animals. It is common in tendon, bone, cartilage, skin, cornea, and other tissues. 1 Up to date, 29 types of collagen have been identified, of which type I, II, III, IV, and IX are more abundant in humans.2–4 At present, most of the experiments on oriented fibers are based on type I collagen. Collagen is usually distributed in tissues in the form of fibers, and whether it can present in tissues in the form of fibers highly depends on its molecular structure.1,5 In vivo, fibril-forming collagen can assemble into fibers, and the fibers are intertwined in a specific form in various tissues. For example, the fibers are arranged in parallel in the tendon, 6 concentric rings in the long bone, 7 and orthogonal grids in the cornea. 8 Collagen self-assembly is a gradual process,9,10 which is influenced by factors such as temperature, pH, ionic strength, and initial concentration.11–13 It is worth noting that the formation of the banding pattern (“D” period) of collagen fibers is due to the presence of potassium ions. 14 In addition, collagen fibers play an extremely indispensable role in the structure and function of tissues. At the cellular level, as one of the main components of the extracellular matrix, the realization of many functions of cells highly depends on the presence of fibers, such as cell adhesion, migration, proliferation, metabolism,15–20 and so on. At the organic level, collagen fibers are an important basis for tissue morphology,21–23 mechanical properties,6,24,25 and wound healing.26,27
As one of the main components of the extracellular matrix, collagen fibers become superior natural scaffolds due to their unique biological structure and biochemical function. Although the application of collagen fibers into scaffolds began in the 1960s,15,28 collagen fibers in scaffolds were not oriented in early tissue engineering experiments. With the deepening of research, researchers realized that the scaffold materials with aligned structures are crucial for cells in vitro, tissue repair, and tissue regeneration to provide a similar tissue structure.29,30 In 2011, Caliari pointed out that “…a range of studies have suggested that successful regeneration templates for natively aligned tissues such as peripheral nerves, the myocardium, and tendon must provide tissue specific aligned contact guidance cues that recapitulate aspects of the tissue anisotropy.” 31 In addition, damage and degeneration of connective tissue and load-bearing tissue are common clinical case, such as lesions and damages of intervertebral disc, 32 anterior cruciate ligament, 33 cartilage, 34 and cornea. 35 Such tissues have no blood supply, low cell volume, weak cellular metabolic activity, and poor self-repair ability.36–38 Therefore, in clinical treatment, tissue repair and regeneration face great challenges. The main treatments for repairing tissue defects are autograft, allogeneic transplantation, and scaffold repair. However, such repair methods have both advantages and disadvantages. First, autologous transplantation is the best approach to repair the defect tissue, but it can lead to lesions, degeneration of the donor tissue, and dysfunction caused by ectopic tissue.39,40 Second, allogeneic transplantation can cause immune rejection and face the problem of serious shortage of supply. 41 Third, the scaffold is a tissue substitute that requires good biocompatibility 42 (directing cell adhesion and proliferation) and degradable properties. 43 With the continuous development of tissue engineering, scaffolds have a great application prospect in drug transport,44,45 bone repair,46,47 tissue regeneration,48–50 etc. However, tissue engineering, which focuses on the regeneration and repair of damaged tissue, still faces the challenge of tissue function repair. 51 Nevertheless, it is worth noting that the advantages of oriented collagen fiber scaffolds in promoting cell migration,52,53 differentiation, 54 wound healing, 55 and the formation of vascular network 56 indicate the necessity of the application of oriented collagen fiber scaffolds in tissue repair, regeneration, and other aspects, especially in the repair and regeneration of bearing tissues with specific fiber orientation, such as discs, cartilage, ligaments, and fibers.
Nowadays, the scientific community has a deep understanding of the mechanism of collagen, structure of fiber, and collagen self-assembly, but there is still a lack of understanding of the role of mechanics in the mechanism of fiber orientation, which is also responsible for the difference between oriented fiber and natural fiber prepared by each method. Furthermore, the study of fiber orientation mechanism can allow us to understand the relationship between tissue structure and function, helping us to solve the problem of tissue function repair in the field of tissue engineering. This review will describe the methods for the preparation of oriented fibers and the existing fiber orientation theory. Moreover, a possible theoretical model of fiber orientation generation is proposed.
Fiber and Tissue Strength
The structure, mechanical properties, and orientation characteristics of collagen fibers, as the basic unit of connective tissue and load-bearing tissue, constitute the unique properties of various tissues. In terms of structure, the collagen molecule is a right helix structure composed of three left-helix polypeptide chains.1,5 The collagen molecules form microfibers in the form of a right helix, and the microfibers stack on each other to form a quasi-hexagonal molecular arrangement. 57 This distinct level of supercoiled structure is the key to the mechanical bearing capacity of collagen fibers, and the presence of a helical structure gives the fibers a higher tensile strength. In terms of mechanical properties, the single collagen I fiber has an elastic modulus of 0.1–0.36 GPa. 58 The detonated fibrils from bovine tendon 59 and the rat tail tendon 60 have elastic modulus of E = 5 GPa and E ≤ 11 GPa, respectively. To further understand the mechanical properties of collagen fibers, Lanti measured that the shear modulus of dehydrated fibers was 33 MPa, and the shear modulus of fibers in solvent decreased to 2.9 MPa under hydration. 61 It is worth mentioning that the mechanical properties of collagen (E = 6–7 GPa 62 ) are better compared with fiber. The reason is that the strength of the covalent bond between collagen is relatively low (the covalent bond can be broken by 2000 pN, 63 while the structure of collagen fractures under 19,000 pN 64 ). In addition, the mechanical properties of the fiber can be further improved by cross-linking. Under the action of cross-linking factors, specific cross-linking bonds can be formed between molecules and fibers (the force of breaking the cross-linking bond is about 400 pN 65 ), thus strengthening the fiber structure. However, the type of cross-linking (enzyme reaction, nonenzyme reaction 66 ), the degree of cross-linking, and the strength of cross-linking all influence the mechanical properties of the fiber, such as the rigidity and strength. 67 Moderate cross-linking is beneficial to the mechanical property of collagen fibers, but excessive cross-linking leads to collagen fibers becoming more fragile. 68 The structure and mechanical properties of the fiber make the tissue have a certain bearing capacity, but the specific mechanical property of the tissue is highly dependent on the orientation of the collagen fibers.54,69–72 In vivo, all kinds of tissues are subjected to mechanical stimulation in various forms and at various amplitudes. With the effect of mechanics, the diversity of fiber orientation satisfies the physiological requirements of tissues. For example, skin that protects organisms is usually subjected to low multidirectional mechanical loads, and interwoven fibers have greater adaptability to multidirectional loads. The tendon, acting as a force transmission medium between muscle and bone to achieve body movement, is mainly subjected to one-way, high-frequency, large-load mechanical stimulation, having the high-tensile strength. The key to superior mechanical properties of tendon is due to abundant highly oriented type I collagen fibers. Moreover, the mechanical response of fiber in tendons plays an important role in bearing capacity. Under the mechanical load, at first, the kinks in the fibers and molecules are stretched out, 73 and the kinks store a part of energy generated by the load in the form of elastic potential energy in a spring-like manner. Subsequently, as the tensile force gradually increases, the fibers and the molecules slide relative to each other, and the sliding between the fibers causes the energy to further dissipate. During this process, the sliding dissipates energy in a damper-like manner and keeps the structure relatively slowly changing under load.74,75 The occurrence of fiber reorganization causes the tendon stress–strain curve to be nonlinear, that is, under lower stress, the fiber undergoes large deformation.76–78 The linear region in the mechanical curve is the process in which the fiber is stretched. Finally, the fiber is stretched to break under load. Figure 1 shows the stress–strain curve of the tendon under static load. In tissue engineering, the mechanical properties of scaffolds are one of the key factors for the successful application for tissue repair, where the consistent mechanical properties of scaffolds compared to natural tissue usually provide better biocompatibility in terms of physiological functions. 40 At present, reconstituted collagen fibers in vitro are basically consistent with natural fibers in terms of structure and mechanical properties. 79 However, the degree of fiber orientation (parallelism between fibers) in collagen fiber scaffolds still has a large gap with the degree of fiber orientation in tendons and corneas. In addition, there is a significant correlation between the degree of fiber orientation in the tissue and its mechanical parameters such as elastic modulus,70,72,76 which may be one of the reasons why the mechanical properties of fiber-rich tissue are superior to collagen fiber scaffolds. An in-depth understanding of the mechanism of fiber orientation in tissues may provide a solution to the problem of fiber orientation improvement in biological scaffold. For this purpose, the next section will demonstrate the relevant fiber orientation theory.
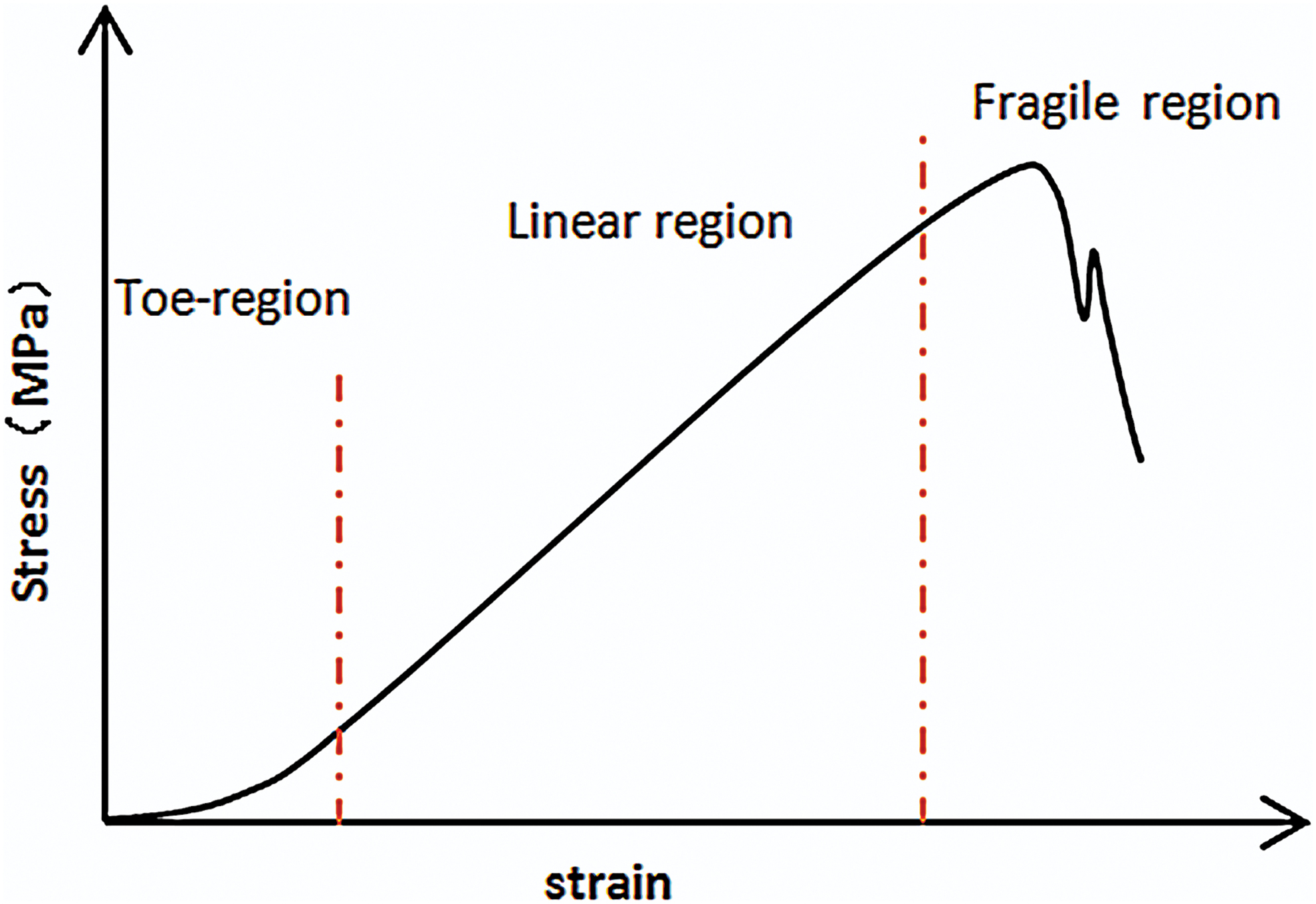
Schematic diagram of tendon stress–strain curve. Color images are available online.
Fiber Orientation Theory
In 1984, Birk and Trelstad found collagen fibers in the compartments of fibroblasts by observing chicken embryonic corneal sections 80 and then observed the same phenomenon in chicken embryo tendon. 81 They believed that microfibers are formed in the compartments, and the microfibers grow into a dense bundle by lateral fusion of compartments as demonstrated in Figure 2. In 2004, based on experimental observations of chicken embryonic and rat tail tendon, Canty proposed the theory of “fibriposistors”, that is, the fibers are secreted into the extracellular matrix through the pores formed by the fusion of the microtubule carriers for transporting microfibers and the cell protrusion. The orientation of the fibers during this process is consistent with the orientation of fibrilposistors. 82 In 2013, Kalson further expanded the theory, believing that the fibrilposistors exert an inward pulling force on the fibers and synergize with the tissue tension to make the fibers organized. 83 Figure 3 is a simplified diagram of the theoretical model of fibrilposistors. Subsequently, Paten showed that mechanics plays an important role in the mechanism of oriented fiber formation by proposing a model explaining the manner of tendon growth under tensile load. 84 In addition, other scholars believe that the formation of oriented fibers is based on the mechanism of collagen liquid crystal, 85 that is, the collagen monomer in high concentration solution self-assembled into oriented fibers under certain confined conditions. However, the liquid-crystal orientation theory cannot perfectly explain the formation of mature fibers. We believe the “fibrilposistors” theory can better explain the mechanism of fiber orientation, and we argue that the mechanical environment plays a pivotal role in fiber orientation. Thus, next section mainly demonstrates the mechanics-mediated methods to form oriented fibers.

Growth model of the fiber bundles. With gaps on cellular membrane fusing laterally, fibers are gathered together to form a fiber bundle. Color images are available online.
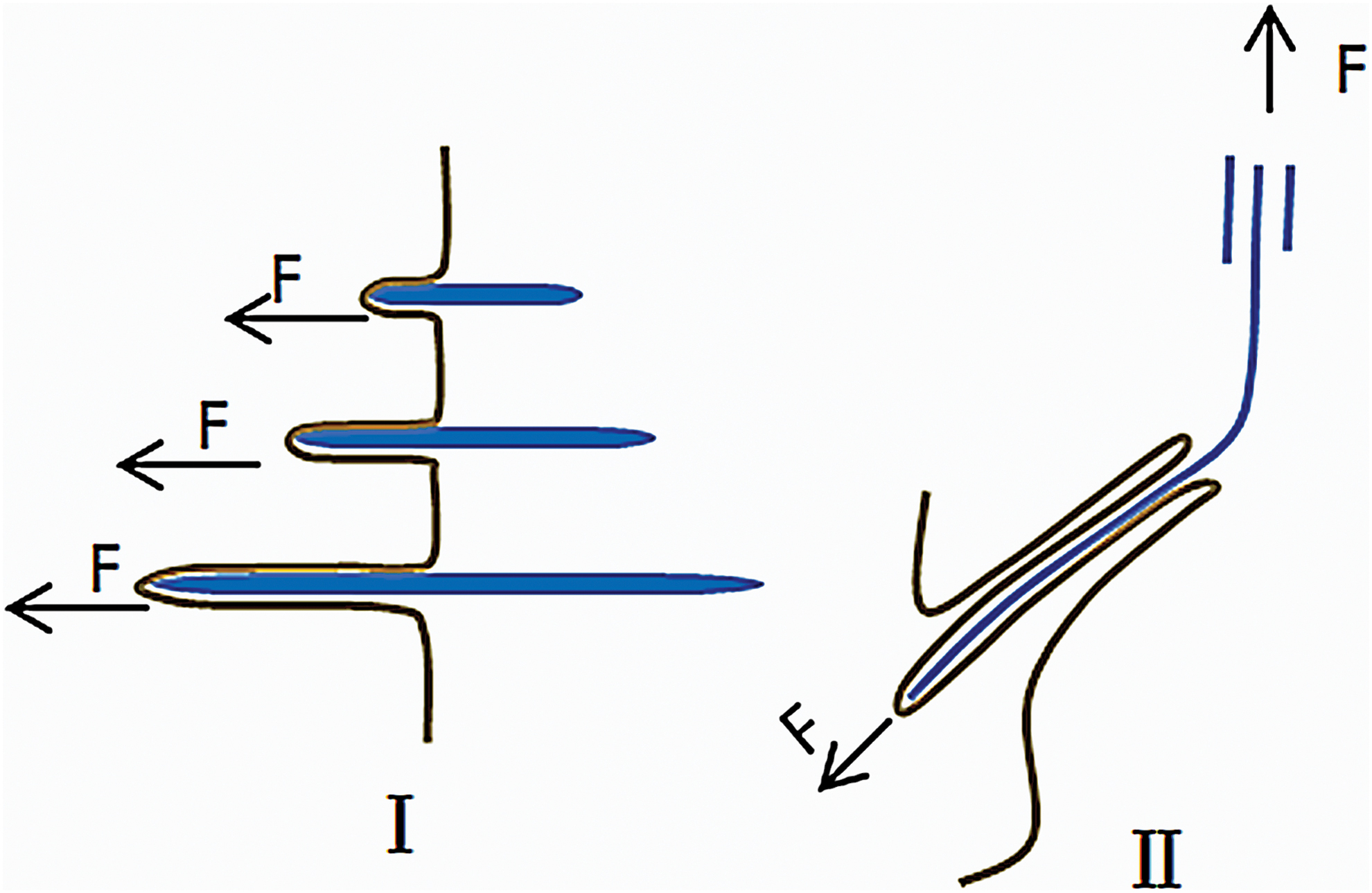
Fibripositor model. The single fiber is extended inward under the tension of fibripositor, which is pulled out of the fibripositor under the tension of extracellular tissue. Color images are available online.
Preparation of Oriented Fibers In Vitro
As an interdisciplinary research field, tissue engineering is committed to the repair and regeneration of tissue structure and function through the combination of cells, biomaterial scaffolds, regulatory factors, and growth environment. 40 Among them, biological scaffold is an important tool for tissue repair and regeneration, which can provide a favorable growth environment for cell culture in vitro and promote the growth and repair of damaged tissue in vivo. Collagen, as a natural material with superior performance, has been widely used in the preparation of scaffolds, and the oriented collagen fiber scaffolds have better repair effect. 55 In addition, the successful preparation of oriented collagen fibers may further improve the properties of artificial tissues86,87 in vitro. Since the 1950s, Bahr 88 discovered that collagen can self-assemble in vitro to form a structure similar to natural fibers. Various researchers have conducted extensive experimental studies on collagen assembly, fibrous structure, and the fiber orientation. In addition, there has been significant improvement in the preparation of oriented collagen fibers. The preparation methods of oriented fibers involve the fields of electricity, chemistry, mechanics, etc. This section mainly focuses on the field of mechanics in the preparation of oriented fiber methods. Mechanical environment is indispensable in the process of tissue growth, which has an important influence on the formation of tissue morphology. In vivo, fibers (tissue engineering implants) are subjected to mechanical stimulation such as tension, compression, and shear due to cell contraction and limb movement during the growth and cluster process, but qualitative research on the mechanical environment has still limited the study on the preparation of oriented fibers in vitro by mechanics allowing us to further understand the role of mechanical environment in the process of tissue repair and regeneration. Therefore, the related literatures of oriented fiber preparation are explained separately in terms of mediators (tensile, shear, and nonmechanical mediating factors).
Shear
According to the loading mode, the preparation method of shear-mediated fiber orientation is mainly divided into fluid shearing controlled by fluid flow and mechanical shearing produced by the relative motion of the acting surfaces.
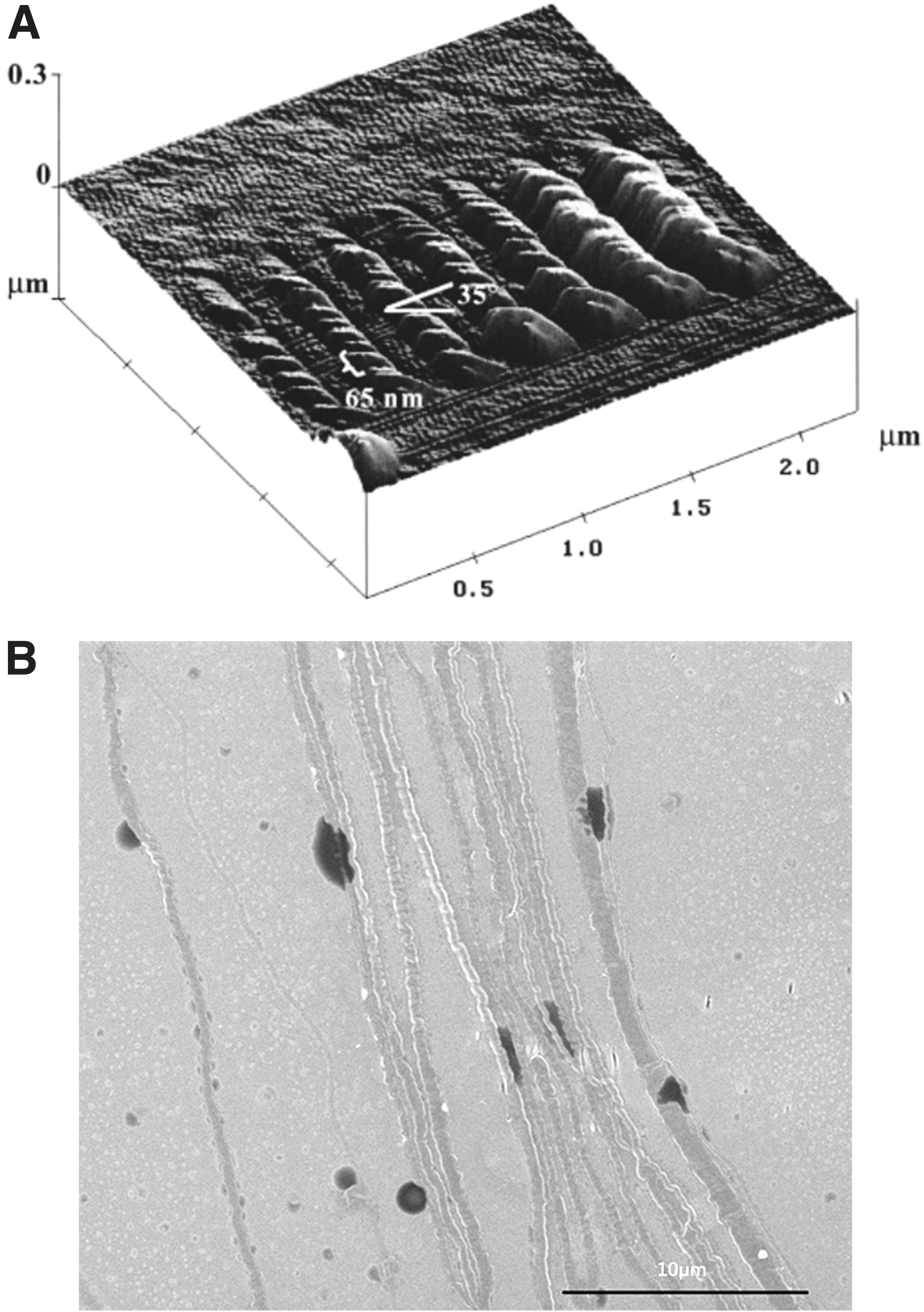
In 2008, Lanfer 89 used a fluid deposition method to prepare oriented fibers. In the experiment, with flow rates of 0.45, 4, and 11 mL/min (corresponding to shear rates of 8.3, 73.8, and 203.1 s−1, respectively), collagen solution flowed through a miniature rectangular tube (8 mm in length, 1 mm in width, 73.6 μm in height). At the flow rate of 11 mL/min, the orientation of fibers prepared is the best, and the degree of fiber orientation gradually decreased with the decrease of the flow rate. The increase of concentration of collagen solution (0.2, 0.4, 0.8 mg/mL) is also helpful to the increase of fiber orientation. In 2009, Saeidi 90 regulated collagen (3 mg/mL) self-assembly by fluid shearing. In the experiment, different shear rates (9, 20, 80, 500 s−1) produced by the fluid flow regulated the assembly process of collagen monomer on the glass sheet, and the prepared fibers were oriented along the direction of fluid flow. Under different shear rates, the axial growth rate of the fibers is different. The maximum growth rate is 0.100 ± 0.025 μm/s under the shear rate of 9 s−1. As the shear rate decreases, the length of fibers increases. However, the ends in the upstream of the fibers are reverse curved (hook shape). The reverse bending angle under the shear rate of 500 s−1 is 180°. Under the lower shear rate, the angle is ∼90°. Furthermore, the oriented fibers produced by this method have no banding pattern. Saeidi believed that the fiber orientation made by fluid flow is influenced by the deposition of precursor of monomers (collagen monomer adsorbing on the glass sheet) and the shear rate. The shear rate affects the distribution of the collagen monomer on the mat. Meanwhile, the mat provides a specific attachment position for the fiber by surface morphology and allows the fiber to grow in a specific direction. Subsequently, based on previous fluid shear experiments, Saeidi 91 adjusted the experimental scheme, producing orthogonally oriented fibers by the combination of fluid flow and a circular film. In the experiment, the collagen solution (2.6 mg/mL) flowed from a vertical conduit to a circular coverslip fixed on a rotating disk, which affected the fiber orientation together with the upper circular film affecting the fiber orientation. After depositing a layer of fibers, the coverslip is rotated 90°. Then the solution flowed again onto the coverslip to form orthogonally oriented fibers. Experiments showed that the degree of fiber orientation varies greatly at different flow rates and rotational speeds. At a rotational speed of 1000 rpm (400–600 s−1) and a flow rate of 1 mL/min, the degree of fiber orientation was the highest. Excessively high rotational speeds prevent the fibers from adhering to the substrate, while at lower rotational speeds, the deposited fibers are disorganized. In addition, the “hooks” still exist. It is believed that the film confinement defines the direction of fiber growth during assembly and promotes fiber distribution along the flow direction. However, the vibration of circular film can result in uneven shear force that affects fiber orientation. In addition, Su 92 used air flow to shear the collagen solution (1, 3 mg/mL) placed on the glass sheet. Experiments showed that the fibers are organized well at the wind speed of 0.5 m/s, and the degree of fiber orientation decreases with the increase of wind speed. He believed that the high wind speed causes the solution to evaporate too quickly, thereby shortening the shearing time and ultimately resulting in a decrease in the degree of fiber orientation. In 2011, Lai 93 used the gap in special syringe to shear a high concentration of collagen solution (30 mg/mL) to prepare fibers oriented along the axis of the syringe. In the experiment, the push rod of syringe was pushed at a speed of 0.124 cm/s (the corresponding shear rate was 221.11 s−1) to shear the preinfused collagen solution (0.1 mL), and then the putter was placed at 37°C to promote collagen self-assembly. He believed that shear and concentration together affect the fiber orientation, because the collagen solution will exhibit a cholesteric liquid crystal state when it reaches a certain concentration. In 2014, Hoogenkamp 94 used counter-rotating cone extrusion technology to prepare oriented collagen fibers. In the experiment, two coaxial tubes with opposite rotation directions sheared the collagen solution (4 mg/mL) between the gap of two tubes. Subsequently, the collagen solution adheres to an axially moving mandrel, forming a fibrous tubular film in which the fiber orientation gradually changes with the thickness of the film (2.6°/μm), that is, the fiber orientation near the outer tube surface area tends to the rotation direction of the outer tube. Close to the inner tube surface region, the fiber orientation tends to the rotation direction of the inner tube and fibers in the intermediate portion distributed along the tube axial direction. It is believed that this phenomenon is caused by uneven shear, that is, the shear action of the rotating surface is gradually attenuated with distance. Figure 4 is a simplified diagram of the technology of the counter rotation cone extrusion.
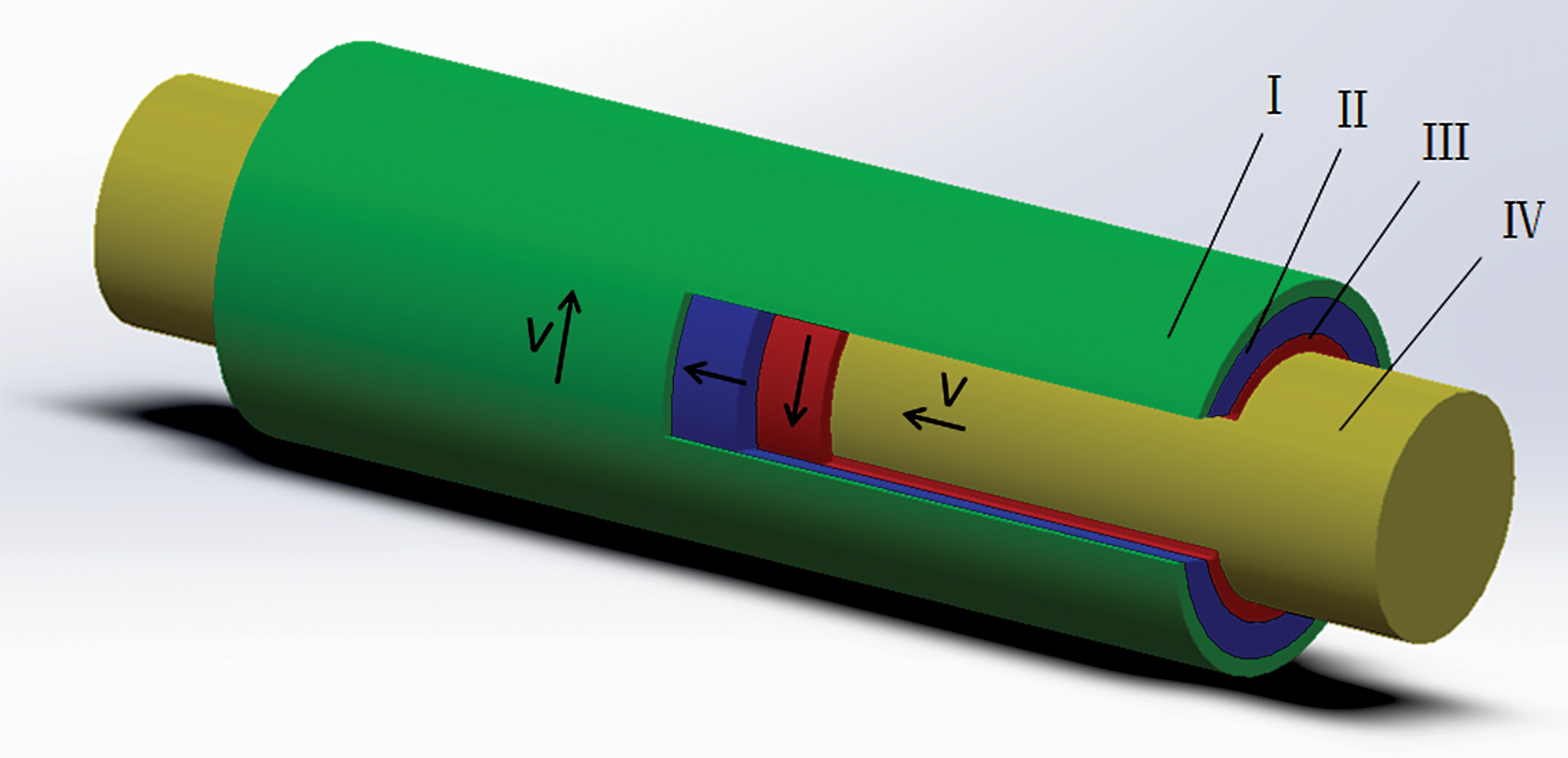
Schematic diagram of the technology of the reverse rotation cone extrusion. I. External cone. II. Collagen solution. III. Inner cone. IV. Mandrel. Color images are available online.
In the above experiments, the oriented fibers prepared under fluid shear were distributed along the fluid flow direction; however, there are some defects on the oriented fibers. For example, single fiber is unevenly distributed with low straightness and accumulation occurred. Figure 5 is the scanning electron microscopy (SEM) image91,93 of the fibers under shear. The higher the degree of fiber orientation prepared by mechanical shear, the less the accumulation phenomenon. However, before exploring the reasons, we need to understand the related concepts of fluid mechanics. Viscosity is the inherent physical property of fluid. Figure 6 is an example to explain the viscosity of fluid, with two plates placed in parallel and gap filled with static fluid. When the lower plate is held stationary and the upper plate is moved at a constant speed, the water layer in contact with the upper plate moves at the same speed with the plate and drives the movement of the next layer. As the distance from the moving plate gradually increases, the speed of movement of each water layer decreases layer by layer to zero. The reason why the plate movement can drive the water layers is that there is internal friction (shear force) between the layers named viscosity phenomenon. In Newton's internal friction law, the shear stress is proportional to the velocity gradient:

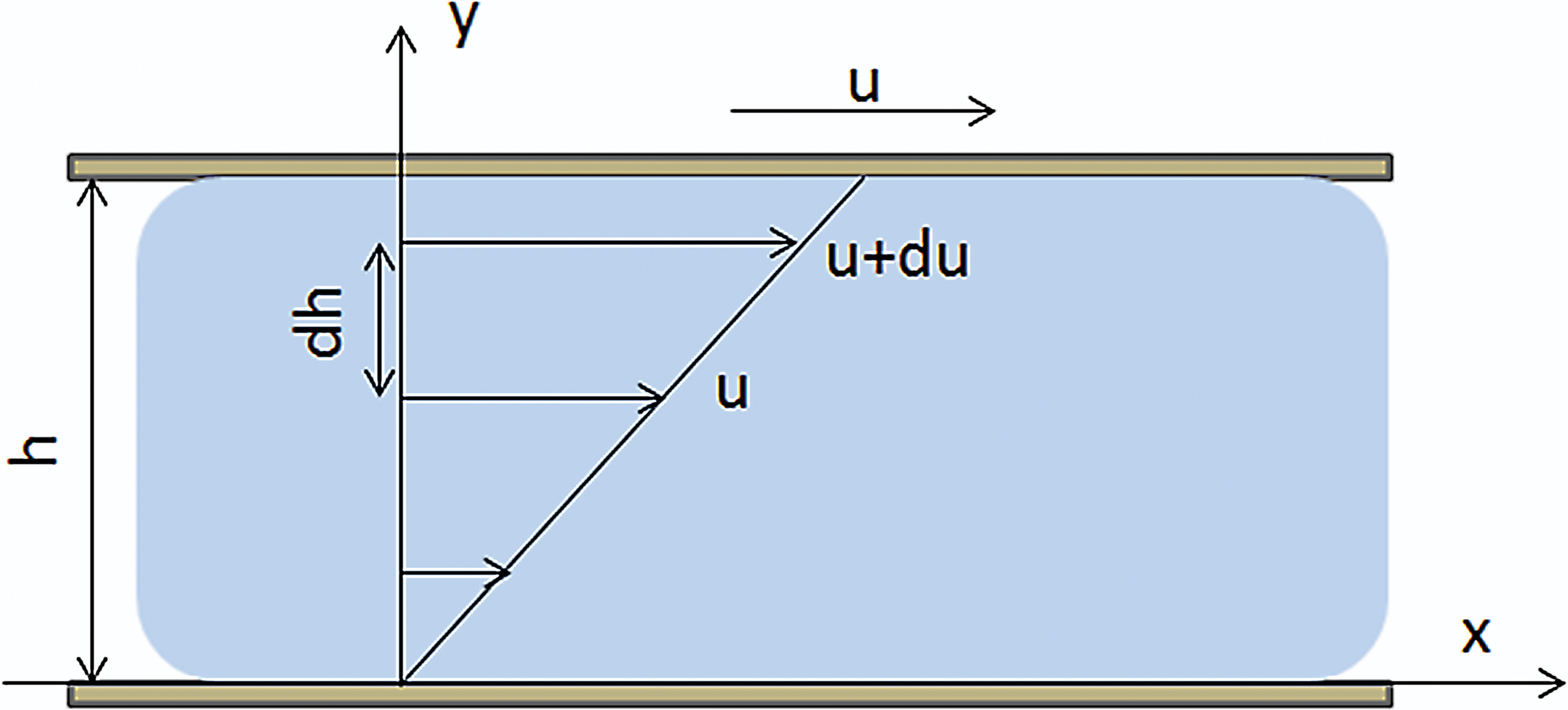
Viscous phenomenon. Color images are available online.
Tension
In the tensile-mediated experiments, the force can act on both collagen monomer and fiber. The effect of the force on the collagen monomers is mainly to promote the directional distribution of the monomers, and the effect of the force on the fibers is to improve the straightness of the single fiber. In 2001, Wilson 95 regulated the self-assembly of collagen monomers by the dip-pen nanolithography. In the experiments, with the thiolated collagen solution adhering on a probe of atomic force microscope, he drew a line (about 2 μm/s) in a specific direction on the gold substrate. The thiolation of the collagen allows it to adhere firmly onto the substrate. During the process, the monomers away from the edge of the probe adhere to the substrate. As the probe moves, the solution attached to the gold surface pulls the collagen monomers on the probe by cohesion and Coulomb force. Thereby, the collagen monomers assemble into oriented fibers. In 2016, based on the theory of flow-induced crystallization, Paten 84 used a glass needle to pull the collagen solution in one direction to fabricate oriented fibers. In the experiments, the micro glass needle was immersed in a collagen droplet with high concentration first. Then the microneedle was slowly pulled out of the droplet (the strain rate was <1 s−1). During the pulling process, a continuous liquid column where collagen monomers assemble into oriented fibers is formed between the microneedle and the droplet. The fibers are arranged in the direction of the force as a whole; however, the fiber orientation distribution is uneven. The fibers at the boundary of the liquid column are arranged in the direction of force, while the inside of the liquid column consists of short fibers with no orientation. The authors believed that the change of fiber orientation from the outside to the inside of the liquid column is the result of complex hydrodynamic action, explaining the difference in fiber orientation at various locations within the liquid column by separating the liquid column and droplet into five regions: tension zone, shear zone, transition zone, circulation zone, and disorder zone. In our laboratory, collagen solution droplets (6 mg/mL) on the cover slip were pulled by a glass sheet (5 mm in width), and the prepared fibers were arranged along the pulling direction as shown in Figure 8B (the pulling direction is parallel to the coverslip, and the velocity is 25 μm/s). In 2008, Voge 96 chose hydrogel as a tensile carrier for cyclic uniaxial stretching stimulation of collagen fibers with a tensile strain of 8%. After stretching, the fibers are arranged along the direction of tension. Subsequently, Vader 97 pulled (12.5 μm/s) the gelatinous fiber solution with a diameter of 20 mm and a height of 2 mm by uniaxial tension. Experiments indicate that as the tensile strain increases, the degree of fiber orientation increases continuously. When the tensile strain is <5%, the degree of fiber orientation does not change significantly. In addition, the orientation of fibers along the line of force is particularly obvious. However, as the distance from the line of force increases, the degree of fiber orientation is gradually reduced.
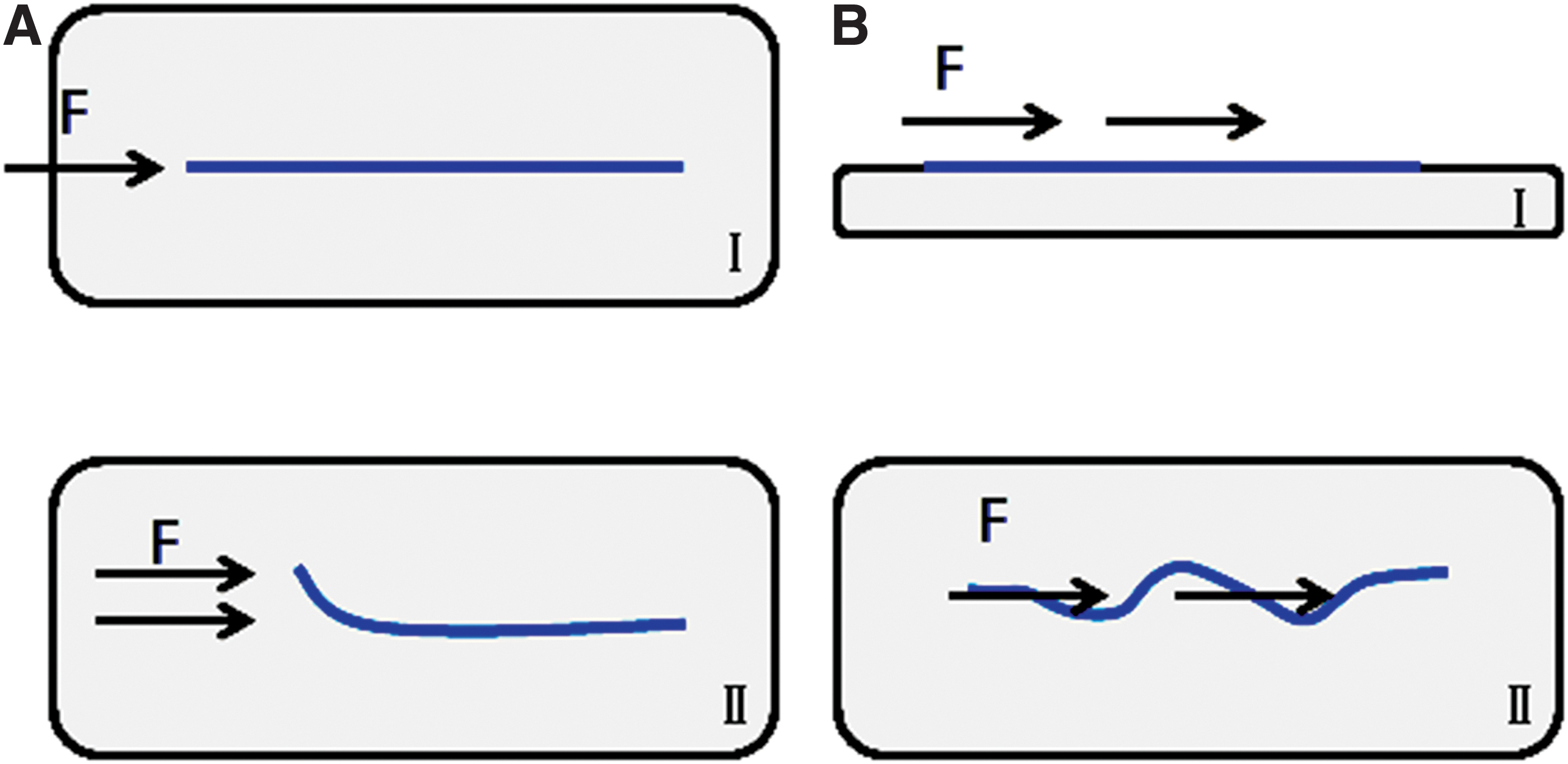
Oriented fibers under tension.
In the above experiments, the oriented fibers formed by tension are also uneven in the degree of orientation. Figure 8 is the atomic force microscopy (AFM) 95 and SEM images of the fibers under tension. Regarding the problem of unevenness in fiber orientation under tension, the difference in structure between collagen monomer and collagen fiber may provide a reasonable explanation. We consider collagen monomer and fiber as rigid body rods and ropes, respectively, and the response of the rods and ropes to the tension is different. The various forces acting at different positions of the rods (different points of force) can be decomposed into major forces in several directions according to the principle of force composition. Thereby they can produce a unified mechanical effect on the rods. However, the forces acting on various positions of the rope cannot be composed and decomposed, and the effects of the forces are always inconsistent. Therefore, the straightness of the fibers under the pulling force is low. Figure 9 is a simplified diagram of the response of collagen monomer and fiber under tension. Although the collagen monomers can be arranged in specific direction under tension, the distribution of the collagen monomers is susceptible to the action of the fluid, thereby affecting the degree of fiber orientation. In addition, we believe that the key to the formation of oriented fibers under tension is low strain rate. During the stretching process with low strain rate, the distal end of collagen monomer binds to other collagen monomers, growing in orientation under internal friction. However, high strain rate produces great resistance and destroys the combination of monomers.

Nonmechanical Factors
With the development of tissue engineering, researchers in various fields have invested a lot of effort in the preparation of oriented fibers and have designed a variety of methods to prepare oriented fibers. Among them, the electrospinning technology98,99 has shown great advantages in the preparation of oriented fibers and has been widely used. In the preparation, the collagen monomers in the collagen solution break through the surface tension of the solution under the electric field and eject from the nozzle of the container, forming a continuous jet stream. Then the solvent rapidly volatilizes to form fibers, and the fibers are collected by a mandrel. It is worth noting that the elastic modulus of the fibers prepared by this method is 52.3 MPa. 98 We believe that the degree of fiber orientation and cross-link is responsible for the lower elastic modulus compared to that prepared by counter-rotating cone extrusion technology. In addition, there are other methods to prepare oriented fibers, such as magnetic-field fabrication,100–102 that is, based on the diamagnetic specificity of collagen under the magnetic field, collagen assembles into oriented fibers perpendicular to the direction of magnetic field, electrochemical fabrication 103 (under solution electrolysis, collagen assembles into fibers perpendicular to the direction of electric field. The ions in the solution need to be removed.), contact guiding53,104,105 (Based on the adsorption capacity of collagen on the surface of substrate, oriented fibers were prepared), wet spinning, 106 and liquid-crystal mechanism preparation. 107 It is worth mentioning that, it has gradually been applied in the preparation of oriented fibers,108–110 with the development of 3D printing technology; however, the degree of fiber orientation needs to be further improved. Compared with mechanical method, nonmechanical method, such as electrospinning, magnetic field, and so on, can set all kinds of parameters accurately and conveniently, to control the change of fiber orientation. However, those methods have a little guidance for the study of the mechanism of oriented fiber formation and tissue repair in vivo.
The Challenges in Methods
Collagen self-assembly is an entropy-driven process caused by the loss of water between monomers, 10 and the self-assembly process is mainly divided into two stages: nucleation and growth. Based on electrostatic interaction and hydrophobic interaction, the C-terminal peptides of collagen monomers bind to specific binding sites of other collagen monomers to form microfibers, 111 and the change of ionic strength and pH value can affect the interaction between molecules. 105 In addition, the goal of the mechanics-mediated fiber orientation experiments is not to recombine the collagen monomer by overcoming of the interaction between the monomers, but to impose additional external force on the interacting collagen monomer on the basis of the intermolecular interaction, which can cause the monomers to bond along the direction of force.
However, there is a general problem in various mechanics-mediated methods, that is, the force does not affect the collagen monomers and the fibers evenly. In the fluid shearing experiment, the “hooks” and the stack are an obvious indication that the shear force acting on collagen monomer is not even. The reason for the absence of “D” period may be due to fluid flush causing the potassium ion to be lower than the critical concentration. 14 Mechanical shearing solves the problem of uneven force action, so that the fiber orientation degree is greatly improved. In the microneedle drawing method, there is a difference in fiber orientation in the fiber bundle. The reason for the difference may be due to uneven force inside and outside the fiber bundle. The fiber orientation on the outside is mainly affected by tension, while the fiber orientation on the inside of the fiber bundle is affected by hydrodynamics. Although Voge used a porous hydrogel as a stretching medium to circularly stretch collagen fibers, the degree of fiber orientation after stretching improves to a certain degree due to the isotropic structure of hydrogel. During the stretching process, there are various forces acting on the fibers, and the acting points of force are not limited to the two ends, leading to the curved morphology of fibers after tension. However, the dip-pen nanolithography technology solves the problem by preparing oriented fibers from nanometer scale. In the experiments, the nanoscale probe of atomic force microscope precisely controls the position of each collagen monomer, so that the unevenness of force is controlled much better. In contrast, in the magnetic field and electrochemical methods, the degree of fiber orientation prepared is relatively high and relatively uniform. The reason is that the action of the magnetic field and the electric field is directly applied to each collagen monomer by the diamagnetic and electrical properties of the collagen monomers.
In general, fibers prepared by electric fields, magnetic fields, and electrochemistry have a higher degree of fiber orientation, and the orientation of fibers regulated by mechanics is relatively low. However, experiments indicate that the mechanics-mediated preparation method has a large potential to prepare highly organized fibers, since the fibers prepared by the mechanical force are arranged closely together, and the straightness of the single fibers is relatively high. In addition, the effect of uneven loading may be better solved in the tissue—cells exert forces on the fibers at the micro-nano dimension scale. This also shows the important role of mechanics in the fiber orientation in vivo.
Discussion
The factors affecting the structure of tissues are mainly genes and epigenetic environment, while the mechanical environment is the main epigenetic environment factor. 21 The mechanical environment also plays an indispensable role in the process of formation of oriented fibers in tissues. The effect of animal joint load on the structure of fiber network 112 and the improvement of fiber orientation of the tendon 113 better illustrate the effect of the mechanical field on fiber orientation. However, the critical role of mechanics has been ignored in the existing fiber orientation theory and model. There are some clues in the relevant biomechanical and mechanical biology experimental research, which may help us further understand the role of mechanics in fiber orientation formation.
In 1980, Harris 114 discovered that fibroblasts placed on a silicone wrinkled film by applying a shear force (0.001 dyne/μm) to the film. Subsequently, Harris 115 found that cells placed on the collagen gel arranged fibers in the direction of the tension generated by the cellular contraction, and the fiber bundles were formed between the cells. As an environment detector of cells, filopodia play a central role in cell migration.116,117 Related literatures indicate that filopodia can pull granulum around the cells.118,119 Kress 120 believed that the filopodia pull the particles in a stepwise manner, and the pulling speed is related to the traction force. The measured maximum speed is 600 nm/s, and the maximum traction force is 19 pN. It is worth noting that collagen monomers can exhibit relevant mechanical curves under the action of 14 pN. 64 We believe that the cells indirectly control the fiber orientation by pulling the collagen monomers through the filopodia, and the filopodia have the ability to pull the microfibers away from fibripositors. In addition, Paten also pointed out the important role of filopodia in fiber orientation. Regarding the mechanism by which fibers gather into bundles, the growth process of fibers observed by Birk gives us some inspiration that the individual fiber in each fibripositor is pulled together to form small size fiber bundles. Then, bundles further aggregate. Based on the above analysis and the results of experiments on oriented fibers, we propose a model of the formation and growth of oriented fibers. Figure 10 shows the model of fiber formation and growth in tendons. In the model, initially, collagen nucleates on the cell membrane and assembles into microfibers as the amount of collagen monomer is increased.10,121 Subsequently, fibripositors grow inward, applying an inward pulling force to the fiber. 83 During the growth of the fibripositors, the peristaltic movement of the filopodia pushes the fibers away from the cells that secrete collagen. In contrast, filopodia of adjacent cell exert an outward pulling force on the microfibers. The pulling force applied to the microfibers by filopodia is stronger than that produced by fibripositors, which causes the fibers to move out of fibripositor at a slow rate. Since the fibers move at a slow speed, fibers have a relatively sufficient time to grow inwardly, and the fibers discharged away from the fibripositors are arranged nearly in parallel during the process. When the distal end of the fibers contacts with adjacent cells, the cellular contraction can generate sufficient force to cause the fibers to aggregate into bundles. During this process, the decorin secreted by cells inhibits the occurrence of cross-linking, 122 allowing the fibers to slide past each other to gather tightly. Subsequently, tension generated by cell contraction causes the bundles to continue to move, at which time the cells can completely wrap the small-sized fiber bundles, eliminating the effect of the decorin. Thereby, the cross-linking substance secreted by the cells can cause cross-linking of the fibers. Due to the hysteresis of cross-linking, the wrinkles caused by the contraction of cells can be repaired, to some extent, under tension, but the kinks are formed on the fibers due to incomplete repair of the wrinkles. 123 Meanwhile, short fiber bundles on the same axis are merged end to end to form a continuous long fiber bundle. During the growth of the fiber bundles, the filopodia of cells secreting collagen gradually apply tension to the fibers, and the resistance encountered by the movement of the fiber bundles gradually increases. When continuous fibers are formed between the bone and the muscle, the force generated by the contraction of the cells cannot pull the fibers to move. At this moment, with the cells unable to move the fiber bundle, cells migrate to further optimize the structure of the fibers by working with the tension generated by the muscle and the bone. Finally, the tendon structure is formed.
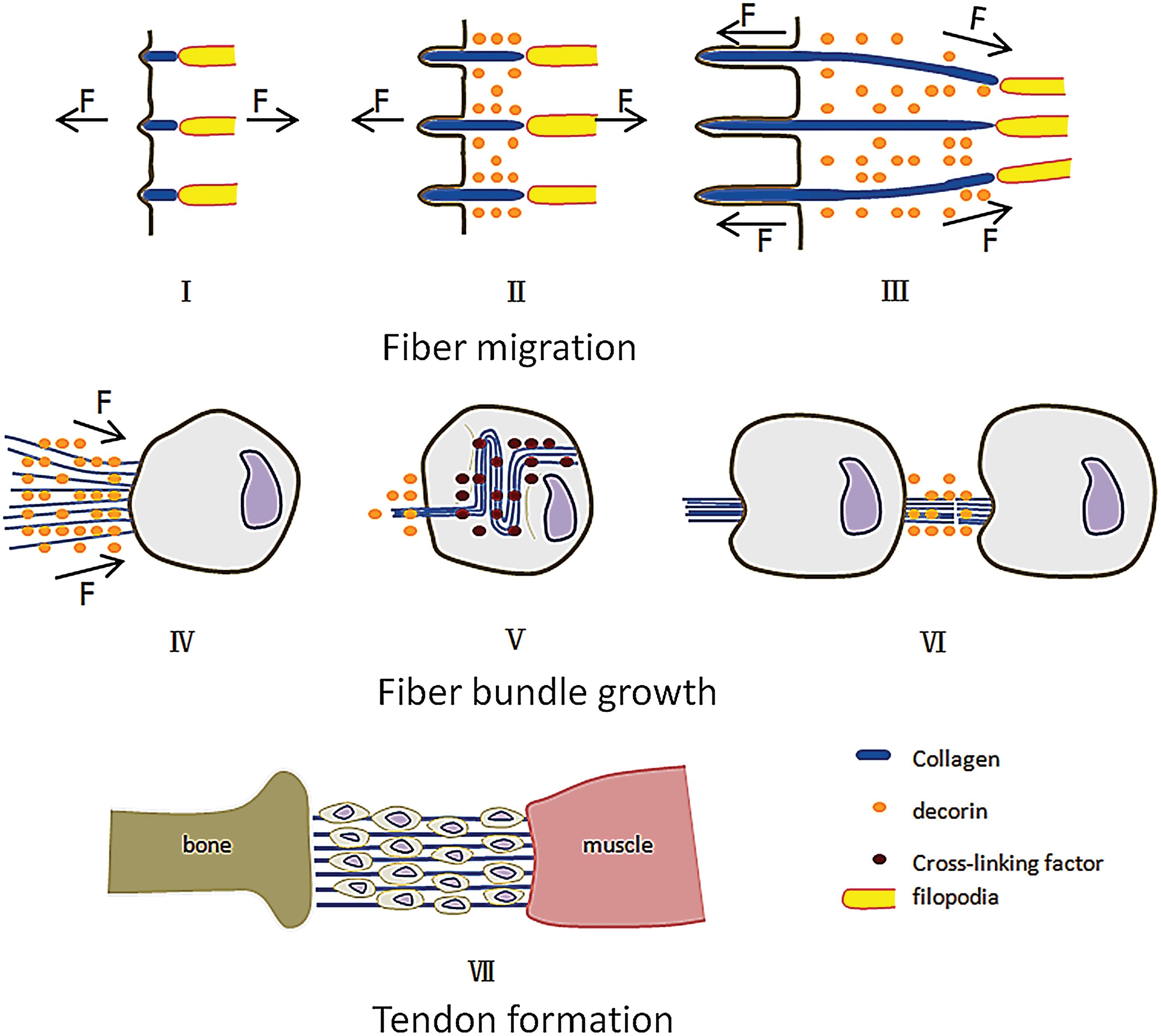
Tendon growth model. I. Collagen assembles on the cell membrane to form microfibers. II. The fibripositors internalize and extend. Tension generated by filopodia and fibripositors promotes the axial growth of microfibers. Decorin inhibits the lateral fusion of the fibers. III. During the growth process, the fibers are slowly pulled out by tension, and the fibers are gradually gathered together by tension. IV. The fibers are further gathered by cellular contraction. V. Cells secrete cross-linking factors to promote cross-linking of the fibers. VI. Fiber bundles are integrated at the beginning and end. VII. The tendon is formed. Color images are available online.
Although our model can better explain the formation of oriented fibers in tendon, there is still a lack of explanation on how cells produce different axial forces in the formation of corneal fiber orientation. However, the experiment taken by Wilks 124 may provide clues as to whether the distribution of cells can influence the direction of force. In addition, the model cannot explain the problem of initial cell migration, but some scholars have shown that the mechanical environment can affect cell distribution. 125 Based on our model, we argue that controlling cellular distribution to mediate fiber orientation will be the future research direction.
Summary
At present, the mechanical preparation of oriented collagen fibers has many deficiencies; however, it has a great potential to prepare highly oriented fibers. In terms of fiber orientation mechanism, we still have some shortcomings in the study of the role of mechanics in fiber formation. The study of mechanical factors is still limited to qualitative understanding, in terms of the mechanical model proposed in this article. From this point of view, future research should focus more on quantitative analysis of the role of mechanics in the formation of oriented fibers. In addition, cells have an indispensable effect on the formation of oriented fibers, which are also sensitive to the mechanical field. Future research on the mechanical response of cells may give us a more comprehensive understanding on the role of mechanics in the formation of oriented fibers and tissue morphogenesis.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
The project was partly supported by the National Natural Science Foundation of China (11432016, 11672208, 31700812) and Natural Science Foundation of Tianjin City (18JCZDJC36100, 18JCYBJC95200, 17JCYBJC41100).