
Abstract
Nitric oxide (NO), which is synthesized by the enzyme NO synthase (NOS), is a versatile endogenous molecule with multiple biological effects on many tissues and organs. In dental pulp tissue, NO has been found to play multifaceted roles in regulating physiological activities, inflammation processes, and tissue repair events, such as cell proliferation, neuronal degeneration, angiogenesis, and odontoblastic differentiation. However, there is a deficiency of detailed discussion on the NO-mediated interactions between inflammation and reparative/regenerative responses in wounded dental pulp tissue, which is a central determinant of ultimate clinical outcomes. Thus, the purpose of this review is to outline the current molecular understanding on the roles of Janus-faced molecule NO in dental pulp physiology, inflammation, and reparative activities. Based on this knowledge, advanced physicochemical techniques designed to manipulate the therapeutic potential of NOS and NO production in endodontic regeneration procedures are further discussed.
Impact statement
The interaction between inflammation and reparative/regenerative responses is very important for regenerative endodontic procedures, which are biologically based approaches intended to replace damaged tissues. Inside dental pulp tissue, endogenous nitric oxide (NO) is generated mainly by immunocompetent cells and dental pulp cells and mediates not only inflammatory/immune activities but also signaling cascades that regulate tissue repair and reconstruction, indicating its involvement in both tissue destruction and regeneration. Thus, it is feasible that NO acts as one of the indicators and modulators in dental pulp repair or regeneration under physiological and pathological conditions.
Introduction
Dental pulp inflammation (pulpitis) results from the activation and interaction of host immune processes in response to pathogens and their toxins in the context of root canal and other oral infections or physiochemical stimuli. 1 These irritations activate resident immunocompetent cells and induce the recruitment of peripheral immunocompetent cells, including activated macrophages, neutrophils, dendritic cells, and lymphocytes, to the lesion region; these cells then release a variety of inflammatory mediators to kill microorganisms.2,3 If microbial stimuli are left untreated, they may cause progressive inflammatory reactions in the pulp that lead to pain and tissue necrosis; however, pulpitis resolution and tissue repair are possible if the harmful stimulus are eradicated before formation of pulp irreversible damage. 4
Depending on the state of the inflammation, different molecular factors, such as antimicrobial peptides, proteases, cytokines, inflammatory mediators, and growth factors, have been indicated to contribute to pulpal defense mechanisms.2,5 Dental pulp exists in a rigid chamber and is therefore in a low-compliance environment; infiltration of the pulp with overproduced proinflammatory mediators further stimulates mediator generation and accumulation, inducing a vicious cycle that potentially causes pulpal necrosis.3,6 Hence, control of inflammation is critical to conduct regenerative endodontic procedures. 7 The inflammatory mediators involved in pulpitis mainly include nitric oxide (NO), prostaglandin (PG), interleukins (IL; IL-1, IL-6, IL-10, IL-12), monocyte chemoattractant protein-1 (MCP-1/CCL2), interferon-γ (IFN-γ), tumor necrosis factor alpha (TNF-α), cyclooxygenase 2 (COX2), etc., and the foremost index of lesion development is NO, which is closely related to the production and action of many other mediators.3,8–10
NO is an important biological effector and intracellular messenger molecule with multiple physiological and pathological functions in various tissues, including oral tissue.11,12 Inside dental pulp tissue, the cell types that are responsible for NO production remain unclear; most researches indicated that NO may be synthesized by immunocompetent cells and dental pulp cells (DPCs)/odontoblast13,14 and mediates not only proinflammatory activities 13 but also signaling pathways that regulate cell growth and differentiation, 15 suggesting its involvement in both tissue destruction and wound healing. Although it is widely recognized that uncontrolled pulp inflammation inhibits reparative events, it also appears that low levels of inflammation are essential, at least initially, to favor the reparative process. 4 Therefore, it is plausible that NO serves as an indicator and modulator during dental pulp inflammation and repair or regeneration.
As dental pulp tissue exhibits both innate and adaptive immune responses that abolish and heal insults if most of the antigens are eradicated early enough16,17; recent investigations have concentrated on the development of novel immunogenic materials to modulate pulp immunity and maintain its vitality. 18 Through the use of several optimized agents, the regenerative endodontic approach aims to shape a microenvironment in which the host's immune defense system does not enhance tissue-injurious proinflammatory processes but to induce the natural regenerative potency of the dental pulp.7,18 Preexisting and/or newly formed odontoblasts from DPCs within the pulp are believed to be responsible for the innate immunity and mineralization of the reparative dentin matrix.19–21 Thus, this review article summarizes the physiological and pathological effects of the NO system on DPCs/odontoblasts and other cell lineages involved in immune responses inside dental pulp tissue and then discuss its potential role as a therapeutic target and tool to deal with dental pulp inflammation.
Brief Review of Nitric Oxide Synthesis and Signal Transduction
NO production mechanism
In mammals, NO is predominantly produced by genuine immune system cells and vascular endothelial cells at varied concentrations (nM–μM) from intracellular five-electron oxidation of
The nNOS form is abundantly expressed in the brain and peripheral nerves, while eNOS is present mainly in endothelial cells; however, nNOS and eNOS expression is not restricted to neurons and endothelial cells, respectively. iNOS has been identified in a wide range of cells, such as bone cells, glial cells, and peripheral blood cells.12,22,25 eNOS and nNOS, which are constitutively produced in low levels (and thus are both called cNOS), are typically activated by intracellular calcium ion (Ca2+) influx and subsequent binding to calmodulin to synthesize low levels (on the picomolar to nanomolar level; pM–nM) of short-acting NO (seconds to minutes), which in turn promotes calmodulin binding to cNOS. In contrast, iNOS has a higher affinity for calmodulin at very low intracellular Ca2+ levels (below 40 nM), which thus acts in a Ca2+-independent manner, and can be induced at high levels for prolonged periods and can produce copious amounts of NO up to micromolar (mM) levels for sustained periods of time (hours to days) in response to different mediators, including inflammatory incentives such as bacteria lipopolysaccharide (LPS) and cytokines (e.g., IL-1β, TNF-α, or IFN-γ). In turn, NO can further affect the synthesis of the following chemokines: IL-1, IL-6, IL-12, TNF-α, and PG.22,26 Similarly, NOS enzymes are responsible for endogenous NO synthesis; in turn, NO itself may negatively regulate NOS activity through feedback regulation.27,28
All isoforms of NOS synthesize endogenous NO by consuming
Biological functions of NO and relevant mechanisms
NO can mediate multiple cellular activities, such as apoptosis, differentiation, neurotransmission, immune responses, vascular tone regulation, platelet aggregation, and angiogenesis (Fig. 1). 23 Accordingly, the underlying chemical mechanisms of NO are complex processes that include numerous possible reactions exerting effects in two distinct categories: direct and indirect effects.

Examples of the biological effects of NO at different concentrations and the related signals. AKT, protein kinase B/PKB; cGMP, cyclic guanosine 3′,5′-monophosphate; EGFR, epidermal growth factor receptor; HIF-1α, hypoxia inducible factor-1α; HO-1, heme oxygenase-1; MAPK, mitogen-activated protein kinases; NF-κB, nuclear factor-kappa B; NO, nitric oxide; Nrf2, nuclear factor erythroid 2-related factor 2; PKG, protein kinase G; sGC, soluble guanylyl cyclase. Color images are available online.
The direct effects of NO, which happen at low steady-state levels of NO (nM), involve chemical reactions of NO with biological target molecules (e.g., direct reactions with transition metals in heme proteins to yield nitrosylated derivatives), which tend to be regulatory in nature but may be cytoprotective or even cytotoxic in some circumstances. 31 In contrast, the indirect effects, which arise at relatively high NO levels (mM), include reactions of NO with O2 or O2− to form reactive nitrogen species (RNS) such as peroxynitrite anion (ONOO−) and S-nitrosothiols, which subsequently interact with biological targets and tend to be cytotoxic and to impair the activity of target enzymes and proteins, including transcription factors and mitochondrial enzymes, producing long-term cellular effects. Furthermore, the indirect effects can be subdivided into nitrosative stress and oxidative stress.31,32
NO signaling transduction (NO/cGMP-dependent or -independent signaling)
NO signaling can be classified into classic cyclic guanosine 3′,5′-monophosphate (cGMP)-dependent or nonclassic cGMP-independent mechanisms. 25 The cGMP-dependent pathway is thought to be a mode of “long-range” signaling, as the signal travels a relatively long distance from the NO sources. In contrast, the cGMP-independent pathway is referred to as a “short-range” scheme, since the action occurs relatively close to the NO source and often within a certain subcellular location. 33 For instance, in cerebellar granular neurons, endogenous NO exerts its effects in a cGMP-dependent manner, while exogenous NO shows its effects in a cGMP-independent manner. 34
The cGMP-dependent pathway mediates the direct actions of NO; the most representative example involves eNOS-generated NO in vascular endothelial cells. Upon diffusion into the subendothelial space, NO undergoes nitrosylation and activates the heme enzyme-soluble guanylyl cyclase (sGC). 31 sGC is a heterodimer made up of two different alpha and beta subunits. In general, two alpha subunits (α1 and α2) and two beta subunits (β1 and β2) are recognized, but β2 has not been identified at the protein level and does not generate a catalytically active sGC upon coexpression with the other subunits, thus indicating that the β2 gene may be a pseudogene.25,35,36 Most tissues possess the heterodimers, α1β1 (NO-GC1) and α2β1 (NO-GC2), which have been shown to exhibit enzymatic activity and to be potentially expressed in almost all cell types except platelets and erythrocytes; however, the ratios of the isoforms in individual cell types are not known. 35 Under physiologically low NO concentrations, cNOS-generated NO reacts directly with the iron atom in the heme group of sGC to activate enzymatic conversion of guanosine 5′-triphosphate (GTP) into the intracellular second messenger cGMP. cGMP exerts its effects primarily by cGMP-dependent protein kinase (protein kinase G or PKG). 37 Upon facilitating the formation of cGMP, the NO-sensitive enzyme sGC acts as a receptor for NO within the NO-cGMP signaling cascade, triggering a variety of cellular responses. 25 Under pathological conditions (e.g., endothelial dysfunction), NO-dependent cGMP signaling is impaired; thus, novel NO-independent sGC stimulators could be useful as both pharmacological tools and therapeutics. 38
In addition to activating sGC to produce cGMP, NO has also been implicated to play critical roles in normal physiology as well as in pathological changes by the cGMP-independent signaling pathway (e.g., through S-nitrosylation of cysteines and nitration of tyrosines), which is mainly involved in the indirect effects of NO.25,27 The protein S-nitrosylation reaction (in which NO reacts with cysteine thiols to form S-nitrosylated proteins) is an important posttranslational modification and thus regulates protein structures and functions.33,39–41 Tyrosine nitration is another modification induced by NO-derived species. NO reacts with O2− in a diffusion-limited manner to form ONOO−, which may cause significant oxidative damage on various target biomolecules, such as lipids, sugars, DNA, and proteins.25,42
Functions of the NO System Inside Dental Pulp Tissue and the Involved Mechanisms
The dental pulp contains a rich microcirculatory system and features greater sensory nerve innervation than other oral tissues. Mature pulp tissue has a strong resemblance to embryonic connective tissue and is responsible for different functions according to its various components. 6 DPCs, odontoblasts (highly specialized cells arranged in a layer along the periphery), and mesenchymal cells are mainly responsible for the regeneration of dentin that has been damaged by dental caries or other insults, physically aiding in the removal of exogenous stimuli by producing extracellular matrix components and tertiary dentin on the pulp chamber surface. Immune cells, including macrophages, lymphocytes, dendritic cells, and leucocytes, immunologically protect the tooth from pathogens and foreign antigens. Blood vessels (endothelial cells) deliver nutrients to the tooth and remove waste products, and a neural network (neurons) protects against harmful stimuli, such as pain, through the apical foramen of the tooth. 43
Accordingly, relevant researches have focused on the role of NO in many physiological and pathological activities of dental pulp, including odontogenesis, angiogenesis, immune responses, neurotransmission, etc.3,8,14,44–50
Expression of NOS and NO/sGC/cGMP signaling molecules in dental pulp tissue
Under physiological conditions, nNOS has been identified in human pulpal nerve fibers and rat molar odontoblasts (Table 1).47,51 eNOS has been identified in both endothelial cells and odontoblasts within healthy human dental pulp 52 and rat molars.47,53 These findings suggest that cNOS are physiological sources of endogenous NO in odontoblasts. It is generally recognized that iNOS is not expressed in healthy pulp tissue 44 ; however, some researchers have detected weak iNOS protein expression in healthy pulp.54,55
Identification and Functions of Nitric Oxide System Inside Dental Pulp Tissue and Involved Mechanisms
Ca2+, calcium ion; cGMP, cyclic guanosine 3′,5′-monophosphate; DPCs, dental pulp cells; eNOS, endothelial NOS; HO-1, heme oxygenase-1; iNOS, inducible NOS; MAPK, mitogen-activated protein kinases; mRNA, messenger RNA; N.a., not analyzed; NF-κB, nuclear factor-kappa B; NMDA, N-methyl-
In inflamed dental pulp tissue, increased eNOS messenger RNA (mRNA) and protein levels with coexistent vasodilation were identified in human samples 44 ; however, a later study found a decreased expression of eNOS mRNA in rat odontoblasts and DPCs at the early phase (1–3 days) after tooth preparation. 53 Enhanced mRNA and protein levels of iNOS have also been identified in inflamed or hyperemic rat pulp tissue, which is mainly composed of macrophages, neutrophils,3,53 leukocytes, and odontoblasts,44,53 along with perivascular lymphocyte and erythrocyte infiltration. 54 Notably, it is generally accepted that iNOS protein is absent from most cells under resting conditions, but that the iNOS gene is regulated by a range of inflammatory mediators, such as bacterial LPS, double-stranded RNA, and cytokines (e.g., IL-1, TNF-α, and IFN-γ).22,56 For instance, in an LPS-induced experimental model of rat pulpitis, the gene expression of iNOS has been found to be dramatically increased 3 h and remain upregulated after 3 days despite a reduction by 9 h compared with that in untreated teeth, 57 suggesting an essential role of iNOS induction in high concentrations of NO inside the inflamed pulp tissue. In addition, several investigations have indicated that nNOS and eNOS are also under the influence of expression regulation and that iNOS is present even under resting status.58,59 Thus, it has become manifest that all three NOS isotypes are simultaneously expressed under both physiological and pathological states.
As mentioned above, since NADPH is a common histochemical marker for NOS, many studies have histochemically analyzed NADPH-diaphorase (NADPH-d) to determine the localization of NOS in dental pulp. It has been reported that NADPH-d-positive/NOS-immunoreactive perivascular and solitary varicose axons are present in the dental pulp and gingiva of cats and dogs. 60 In healthy human pulp, strong NADPH-d activity has been identified in the odontoblastic layer and in DPCs, endothelial cells, and Schwann cells, while weak positivity has been detected in the other parts of the central area; in addition, NADPH-d staining density has been found to be more intense within carious teeth with chronically inflamed pulp than in teeth with normal pulp in the stromal area, as indicated by positive staining of inflammatory cells, angioblasts, fibroblast, collagen fibers, and endothelial cells in abundant, newly formed capillaries.8,61
Similar to the case in other tissues, the NO-sensitive enzyme/receptor sGC and its product cGMP have also been identified in dental pulp tissue.47,54 In healthy rat molar pulp tissue, the sGC α1 and α2 subunits are localized in odontoblasts and DPCs, while the β1 subunits and cGMP are localized in odontoblasts, odontoblast processes, and DPCs.47,54 Challenge of the rat molar dentin/pulp complex for up to 30 min using the NO donor (viz. synthetic NO-releasing compound) spermine-N-(2-aminoethyl)-N-(2-hydroxy-2-nitrosohydrazino)-1,2-ethylenediamine (NONOate) increases cGMP staining intensity in odontoblasts, while stimulation with the NOS inhibitor NG-nitro
All above findings are indicative of a plastic pattern of NOS localization and expression that occurs in an isoform- and a pulp status-dependent manner. Research has revealed that the induction of NOS and signaling by relevant molecules are mediated by various proinflammatory molecules, but few studies have explored the role of cGMP in regulating microcirculation in the dental pulp. Hence, the roles of NOS and the NO/sGC/cGMP pathway in dental pulp circulation need to be clarified in the future, as these roles may have great significance for pulp revascularization.
nNOS (NOS I)/NO and dental pulp innervation and nociception
nNOS, which is predominantly expressed in neurons, produces NO that can act as an anterograde and retrograde messenger at synapses involved in nociceptive processing. Moreover, nNOS is also identified in the vascular endothelium and has been considered as an important contributor to the maintenance of cardiovascular system homeostasis. 67 In addition to the involvement in physiological activities, recent studies have demonstrated that nNOS plays important roles in several mechanisms related to neuronal degeneration/plasticity and cardiovascular disorders.68,69
Dental pulp tissue contains both sensory and autonomic nerves to fulfill its defensive and vasomotor functions. 6 The trigeminal nerve endings, pulpal nerve fibers, and odontoblasts are together responsible for the sensory innervation of the pulp and are all nNOS positive.47,70,71 During human tooth germ development, nNOS is expressed in stellate reticulum cells, ameloblasts, and odontoblast clusters at much higher levels in the advanced stage than in the early stage. 71 A previous study reported that the growth of axons into molar tooth pulp parallels increases of NOS activity in rat undergoing inferior alveolar nerve transection, suggesting a favorable action of increased NOS activity in axotomized trigeminal ganglion neurons on the regeneration of injured neurons. 50 These findings raise the possibility that the neuronal functions of NOS and NO may depend on instant circumstances such as the localization of these molecules and the statuses of the cells.
Under inflammatory conditions, both the numbers of nNOS-positive neurons and subsequent production of NO are clearly elevated in Sprague-Dawley rats with experimentally induced chronic pulp inflammation compared with healthy rats,
72
and the increased production of NO may facilitate neurotransmitter release and enhance nociceptive transmission. Peripheral injury due to local pulp inflammation can induce central sensitization and neuroplastic changes in the relative amounts and activity of nNOS in trigeminal nociceptive neurons through activation of glutamate receptors and N-methyl-
Thus, we speculated that nNOS/NO may play an important role in dental pulp neural transmission. However, it is not involved in neurogenic inflammation that is believed to be mediated by neuropeptides released from sensory nerves at the site of inflammation, such as substance P and ROS. 6 Overall, the existing studies have not clarified the action of nNOS in dental pulp inflammation, and there is a lack of relevant studies on the function of nNOS in pulp microcirculation.
eNOS (NOS III)/NO and dental pulp microcirculation
eNOS is mainly detected in vascular endothelial cells and is the major isoform regulating vascular function. 67 Increases in intracellular Ca2+ due to vasodilator agonists, such as acetylcholine and bradykinin, induce activation of eNOS and enhanced synthesis of NO. NO liberated from the vascular endothelium can regulate vascular tone and blood flow through activating sGC/cGMP signal; it can also inhibit platelet aggregation and adhesion, leukocyte adhesion and migration, and vascular smooth muscle cell proliferation, thus preventing atherosclerosis.11,12 In addition, eNOS exerts an anti-inflammatory effect through promotion of NO and regulation of cGMP signaling and nuclear transcription factor (nuclear factor-kappa B [NF-κB]) pathway, thus inhibiting the expression of inflammatory genes. 76
The dental pulp microcirculatory vessels is characterized by a hierarchical structure, low compliance, firm and resilient tissue, and a lack of a true collateral blood supply.
6
As suggested above, endothelial cells, DPCs, and odontoblasts in healthy human dental pulp are all eNOS-positive cells, and evidence shows that NADPH-d reactivity in pulpal vessels supports the role of NO as a mediator of dental vascular homeostasis by NO/sGC/cGMP signaling.12,47,52,77 A previous investigation using anesthetized rats reported that local application of
Angiogenesis, another critical physiological event associated with wound healing and tissue regeneration, involves complex cellular activities correlated with cell proliferation and migration.87,88 The role of eNOS-produced endogenous NO in angiogenesis has been confirmed by the impaired angiogenetic capacity in eNOS-knockout mice in response to ischemia. 89 Moderate concentrations of eNOS-dependent NO production can significantly regulate endothelial proliferation and migration induced by other key proangiogenic factors, such as vascular endothelial growth factor (VEGF), bradykinin, vitamin D, etc., and therefore contribute to angiogenesis and wound healing.90–93 In dental pulp tissue, cNOS- and VEGF-mediated angiogenesis plays a key role during dental embryo morphogenesis. 71 Bradykinin has been found to exert dual effects on eNOS in the context of odontoblast activation and on protein kinase B (PKB/Akt)-dependent upregulation and extracellular signal-regulated kinase 1/2 (ERK1/2)-independent downregulation of eNOS. 94
Besides its effects on vascularization, eNOS likely exerts an anti-inflammatory effect in healthy pulp and during the early phases of inflammation in hyperemic pulp by restraining the aggregation/adhesion on the endothelial surface of platelet and leukocyte and by increasing tissue perfusion.8,47,95 Furthermore, eNOS is localized in rat and human odontoblasts and is a physiological source of NO in odontoblasts. NO can modulate nociceptive input in double directions: small amounts of peripherally generated NO are algesic, while large amounts of NO are analgesic.44,96,97 Thus, it is possible that eNOS is also involved in nociceptive transmission in dental pulp.
In conclusion, eNOS located in endothelial cells of dental pulp may mediate local vasodilation, inflammation, angiogenesis, and nociception, but the exact functions of eNOS in odontoblasts are still unknown. It is assumed that eNOS-produced NO in odontoblasts regulates the vascular tone of adjacent vessels. Further studies should be carried out to clarify the role of eNOS in the crosstalk between endothelial cells and DPCs/odontoblasts during dental pulp neovascularization.
iNOS (NOS II)/NO and dental pulp inflammation
In comparison with cNOS, iNOS expression is absent in strictly resting cells; instead, it is induced by immunological irritation, such as proinflammatory cytokines and bacterial LPS, in a Ca2+-independent manner.79,98 iNOS-produced NO plays distinct roles in cell activities; for example, it stimulates apoptosis, inhibits mitochondrial respiration, regulates oxidative phosphorylation, and exhibits cytotoxicity in target cells. 22 iNOS-produced NO has also been implicated in the pathogenesis of a range of human diseases, such as arteriosclerosis, diabetes, rheumatoid arthritis, cancer initiation, aging, and acute and chronic inflammation.22,25 With regard to teeth, an example is the iNOS-mediated inflammation-induced bone resorption on the compression side during orthodontic tooth movement. 99
Roles of iNOS/NO in pathogen clearance of pulpitis
The innate immune system provides the first defense line against pulpitis, as it recognizes the pathogen-associated molecular patterns (PAMPs) that penetrate into the pulp tissue through dentinal tubules. 16 In experimental studies clarifying the mechanism of pulpitis, bacterial LPS has been the factor most frequently used to investigate cellular responses to external stimuli. LPS is one of the most potent activators of the human innate immune system. It can be recognized by Toll-like receptor (TLR)-4, and its binding to TLR-4 initiates innate immune responses mediated by TLR-4/MyD88–NF-κB–p53/p21 signaling. 100 Thus, LPS is usually topically administered to mimic pulp inflammation caused by cariogenic microorganisms to some degree; it can cause experimental pulpitis by inducing the production of NO and many other inflammatory cytokines.3,57,101–104 Notably, proinflammatory mediator production is likely to be activated directly by LPS and/or to be related to NO synthesis.3,57 In pulp tissue, immunocompetent cells, DPCs (fibroblasts), odontoblasts, and stem cells are all able to maintain immunomodulatory activity and recognize PAMPs through pattern recognition receptors. In particular, TLR and NO production may participate in the antimicrobial pulp response in an iNOS-dependent or -independent manner.19,21,100,105–107
iNOS synthesis and subsequent NO production can be served as part of the immunologic defense in human physiology and participate in the elimination of invading pathogens, including oral pathogens (e.g., cariogenic Streptococcus mutans).79,108 Previous studies have indicated a concentration-dependent antibacterial effect of NO on endodontic microorganisms and its primary role in the clearance of Enterococcus faecalis by macrophages.48,109 As suggested above, DPCs are able to produce various inflammatory mediators to regulate inflammatory responses inside pulp tissue. Compared with control DPCs, heat-killed E. faecalis-treated human DPCs show increased NO production as a consequence of increased rates of NO synthesis; the generation of NO underlies a key defense mechanism against E. faecalis, supported by the NO-exerted suppression on the growth of this microorganism in a reversible manner. 110
Odontoblasts located at the pulp/dentin interface provide a natural barrier against tooth-invading pathogens and show obvious immunoreactivity for 3-nitrotyrosine (a biomarker for NO-derived ONOO−) in inflamed pulp, indicating the involvement of NOS/NO system in these pathogen-induced pulp inflammatory and immune responses.19,46 In human DPCs, TLR-2- and TLR-4-neutralizing blocking antibodies upregulate iNOS and COX2 expression.19,106 TLR-2-activated human odontoblast-like cells can stimulate iNOS-mediated NO production in the presence of dentin-invading Gram-positive bacteria, which is strengthened by the augment of iNOS protein synthesis, NOS activity, and nNOS and eNOS gene expression in response to bacterial stimulation. 46 The involvement of iNOS in endodontic microorganisms clearance has also been strengthened, at least in part, by an human in vivo study of D'Arcangelo et al. who reported that the expression of iNOS mRNA and protein in inflamed pulp tissues capped with calcium hydroxide [Ca(OH)2] showed an obvious increase after 7 days (accompanied by clear vasodilation and macrophage and giant cell infiltration) but a decreased level after 28 days (disappeared vasodilation and macrophage and giant cell infiltration; the presence of neutrophils, lymphocytes, and DPCs and dentinogenic activity). 111 With regard to periapical lesions, immunohistochemical analyses have shown that iNOS-positive cells are widely distributed in various cell types (mainly neutrophils and macrophages) and are particularly prevalent around blood vessels,10,112 supporting the notion that indicated a key role of iNOS-generated NO in the infection-stimulated periapical bone loss. 9
Regulation of immune responses by iNOS/NO in dental pulp inflammation
iNOS and its related NO not only participate in pathogen clearance but also have immunoregulatory effects, such as connection of innate and adaptive immunity and inhibition of T cell activity.113,114 iNOS expressed by macrophages regulates the balance between the classically activated M1 and alternatively activated M2 phenotype. 113 M1 macrophages are stimulated by LPS, either alone or in association with Th1 cytokines (e.g., IFN-γ), and promote inflammation by producing large amounts of NO and proinflammatory cytokines, such as TNF-α, IL-1β, IL-6, IL-12, and IL-23; in contrast, M2 macrophages are activated by Th2 cytokines (e.g., IL-4 and IL-13) and inhibit inflammation by producing anti-inflammatory cytokines, such as IL-10, TGF-β, and arginase I.115,116 In general, copious quantities of NO synthesized by iNOS have detrimental effects on the immune system; for example, they inhibit I region-associated antigen (Ia) expression on macrophages, downregulate the antigen-presenting ability of dendritic cells, and suppress T-lymphocyte proliferation and leukocyte recruitment.22,72,98
Dental pulp inflammation is an adaptive immune process with changes in local blood flow, neuronal activity, and immunocompetent cell function.5,44,117 Numerous studies have indicated that iNOS-derived NO, COX2-derived prostaglandin E2 (PGE2), histamine, substance P, neuropeptides, IL-1β, IL-6, IL-8, and TNF-α are major inflammatory mediators that are involved in one or more of these processes in pulp inflammation,118–120
while NO may be involved in all steps of the initiation and/or progression of pulpitis.8,72,118 Some evidences for this statement may be gleaned from the following researches. First, it has been documented that iNOS is identified in human pulp tissue with suppurative pulpitis,44,111 in murine pulp exposed to LPS,
3
and in inflamed rat pulp after dental preparation/abrasion and demineralization.13,15 Moreover, the NO receptor sGC, as well as the three mammalian NOS isoforms (including iNOS), have been determined in rat odontoblasts.
47
However, the types of cells producing iNOS in the context of pulpitis have not yet been clarified. A previous study has reported that Ia-negative macrophages and neutrophils are responsible for the generation of NO in the early stage of pulpitis,
3
and DPCs and odontoblasts have also been reported to regulate NO-associated inflammatory responses.
110
Second, iNOS synthesizes large amounts of NO and promotes COX2 catalytic activity through the generation of ONOO−.
121
In a study on LPS-induced experimental pulp inflammation in rat incisors, both the NOS-nonspecific inhibitor
Signaling pathways involved in iNOS expression and NO action during the process of pulp inflammation
Several markers participate in the underlying events mediated by iNOS/NO from downstream to upstream, such as heme oxygenase-1 (HO-1), nuclear factor erythroid 2-related factor 2 (Nrf2), NF-κB, mitogen-activated protein kinases (MAPKs), etc. HO-1, a rate-limiting enzyme that can inhibit immune reactions, is upregulated in the presence of NO, heme, heavy metals, cytokines, growth factors, LPS, etc., and can cooperate with NADPH to degrade heme into carbon monoxide (CO), free iron, and bilirubin; the latter two are converted to ferritin and bilirubin, respectively, which play key roles in NO-mediated cytoprotective and anti-inflammatory effects.57,123,124 In human DPCs, transcriptional activation of HO-1 is induced by Nrf2 in the cytosol, which is bound to a control protein known as Kelch-like ECH-associated protein 1 (Keap1) and interacts with the antioxidant response element (ARE). 125 Under normal conditions, Keap1 promotes the eventual degradation of Nrf2; however, under stress conditions, Nrf2 dissociates from Keap1, translocates to the nucleus, and binds to AREs, initiating activation of the transcription of cytoprotective genes, such as HO-1.123,126 The upstream signaling mechanism responsible for the transcriptional regulation of HO-1 is poorly defined at present. Many investigations have suggested that the most conceivable cascade activating Nrf2/HO-1 signaling is the MAPK cascade; different subfamilies of this cascade can phosphorylate specific target protein substrates, including Keap1/Nrf2, in various lineages, for example, RAW 264.7 cells (p38), 126 human chondrocytes (ERK), 127 and human DPCs (c-Jun N-terminal kinase; JNK). 125
In addition to activating the Nrf2/HO-1 pathway, the MAPK pathway (ERK) also affects the transcription factor NF-κB. 101 NF-κB regulates diverse aspects of innate and adaptive immunity and can trigger a downstream mechanism that affects the production of various cytokines, growth factors, adhesion molecules, chemokines, and immunoreceptors, especially NOS/NO in response to external stimuli, including microbial infection.128–130 Under unstimulated conditions, NF-κB exists in the cytosol bound to the inhibitory protein IκB, forming an NF-κB–IκB complex that is activated through phosphorylation of the inhibitory IκB subunit under the influence of IκB kinases in quiescent cells. Phosphorylation of IκB results in its ubiquitination and subsequent degradation by the adenosine triphosphate (ATP)-dependent proteolytic 26S proteasome; NF-κB is consequently released and then translocates to the nucleus. The activated NF-κB protein released from IκB translocates from the cytosol to the nucleus of the cell and binds to specific DNA sequences to initiate transcription. It thereby regulates the expression of many genes and proteins, mainly those related to inflammatory processes.131,132 For example, NO released from sodium nitroprusside (SNP) can increase IL-8 secretion and mRNA expression in a dose- and time-dependent manner in human DPCs by inducing the phosphorylation of p38 and ERK and the activation of NF-κB. 132 Moreover, NF-κB can also promote exogenous NO-induced odontogenic differentiation of rat DPCs, 49 and endogenous iNOS and HO-1-mediated dental papilla development. 130
Collectively, the data indicate that iNOS is expressed in inflamed DPCs and is involved in all steps of pulpitis. There appears to be reciprocal regulation between the expression of iNOS and the production of inflammatory factors wherein each process is regulated by the other, but the cell lineages synthesizing iNOS remain unclear. The MAPK pathway can regulate iNOS/NO-mediated inflammatory responses by activating downstream signaling molecules, such as HO-1/Nrf2 and NF-κB, eventually controlling the expression of target genes.
Crosstalk between NOS isoforms in dental pulp tissue
Low amounts of NO (presumably in the nM range) produced by cNOS are thought to primarily contribute to the regulation of physiological events; high NO levels (μM) induced by enhanced iNOS expression also play key roles in protective host immune defense, but are nonetheless associated with pathological conditions (e.g., infectious diseases and sepsis). 133 iNOS is a NO regulator that can be affected by NO donors, whereas cytokine-induced iNOS expression is mainly regulated by the transcription factor NF-κB.27,133 Under physiological conditions, low levels of NO (pM–nM) synthesized by cNOS inhibit NF-κB. Soon after a pathological stimulus (e.g., bacterial LPS and/or a variety of proinflammatory cytokines) arrives, cNOS is phosphorylated at a tyrosine residue, thus resulting in enzymatic suppression and decrease of physiological NO levels. Under these conditions, NF-κB is liberated from the inhibitory action of NO; it translocates into the nucleus and participates in the transcription of numerous genes, including iNOS.133–136 In summary, activation of NF-κB requires early declines in basal NO levels (nM range) that are dependent on signaling leading to the inhibition of cNOS, suggesting that the production of NO by cNOS is a suppressor for iNOS expression. Thus, crosstalk between cNOS and iNOS may exist in the context of inflammation. 137
Regarding dental pulp tissue, some information from previous studies supports the possibility of crosstalk between cNOS and iNOS. First, in inflamed or hyperemic pulp tissue, decreased expression of eNOS mRNA in odontoblasts and DPCs has been detected 53 ; in contrast, the iNOS gene and protein expressions of macrophages, neutrophils, and odontoblasts are significantly enhanced.3,53 These results are consistent with the concept that the shift of the inflammatory process into irreversible pulpitis in humans decreases eNOS levels but increases iNOS expression, which may be explained by the negative feedback that is provoked by the large amounts of NO synthesized by the significantly upregulated iNOS.44,138 Hence, we speculate that eNOS and iNOS are inversely regulated in pulp tissue. Moreover, as described above, low amounts of NO (nM) play a protective role by preventing accidental activation of NF-κB, while greater amounts (μM–mM) can induce iNOS expression and NF-κB activation. The NO donor NOC-18 at a concentration of 10 μM has been found to stimulate iNOS expression and NO production, promote odontogenic differentiation through the TNF-α/NF-κB axis, and induce tertiary dentin formation in vivo in rat DPCs, 49 and 1 mM of SNP can stimulate IL-8 production through activation of NF-κB and MAPK. 132 Second, different NOS isoforms have different ligand efficacies. In healthy dental pulp of Wistar rats, stimulation of muscarinic acetylcholine receptors (mAChRs) with the orthosteric agonist pilocarpine (10−9 M) exerts no clear effect on cNOS expression, but pilocarpine at 10−7 M can trigger release of PGE2 and NO through activation of nNOS and eNOS.139,140 In contrast, in LPS-treated rat pulp, a low concentration of pilocarpine (10−9 M) can inhibit iNOS mRNA and NOS activity in vivo and in vitro138,140,141 without exerting an obvious effect on cNOS expression, while a high concentration (10−7 M) produces a positive effect correlated with elevated cNOS gene levels without affecting iNOS expression. 140 Thus, cNOS and iNOS might be inversely regulated in cells or tissues depending on cell status.
Similarly, numerous studies have suggested the existence of crosstalk between nNOS and eNOS. First, their anatomical distributions may provide some clues: (1) nNOS is mainly expressed in parasympathetic vasodilator neurons; (2) and that is also present in nonneuronal cells; (3) eNOS-NO-cGMP signaling molecules has been involved in neurotransmission.47,97 Some typical studies are as following: in an experimental migraine model with stimulation of the rat trigeminal ganglion, most nNOS-immunoreactive nerve fibers were found to be associated with branches of the anterior meningeal artery 70 ; another study has indicated the existence of NADPH-d-positive/NOS-immunoreactive perivascular and solitary varicose axons in the dental pulp and gingiva of cats and dogs that may regulate local blood flow 60 ; in human dental pulp, parasympathetic innervations are distributed around small blood vessels. 142 Second, eNOS can produce NO to maintain the health of oral tissue circulation, not only under resting states but also under conditions subjected to physicochemical stimuli, which may be related to the pathophysiology of inflammatory dental diseases, such as pulpitis and gingivitis. 12 Further research on the correlations between the cellular and subcellular localizations of nNOS, eNOS, sGC, and cGMP and alterations in cGMP staining after challenge with an exogenous NO delivery compound might elucidate the involvement of different NO-sGC-cGMP signaling molecules in neurotransmission modulation, vascular regulation, and dentinogenesis in the dentin/pulp complex, and their crosstalk in these processes. 47
A more recent study reported that the knockout of nNOS in mice can increase the eNOS expression in DPCs and endothelial cells inside the dental pulp, and further induce positive expression of iNOS in odontoblasts that was negative in the control mice. 14 The above findings indicate that crosstalk may exist among three NOS isoforms in dental pulp tissue. However, compared with corresponding evidence for other tissues and cells, the available evidence supporting this notion for dental pulp is obviously unconvincing; therefore, additional high-quality studies should be conducted in the future.
In brief, healthy and inflamed dental pulp tissues have different NOS expression levels and localizations.44,52,54 In healthy pulp, cNOS (eNOS and nNOS) is constitutively expressed in a Ca2+-dependent manner such that it can rapidly produce low levels of NO (pM–nM range), which play protective roles by maintaining pulpal homeostasis and regulating other physiological events. iNOS, an inducible Ca2+-independent isoform, is mainly expressed in inflamed pulp tissue as a result of external stimulation and synthesizes high levels of NO (μM–mM range) for sustained periods of time. These large amounts of NO are involved in inflammatory processes, including pulp defense and repair/regeneration. Evidently, crosstalk between NOS isoforms exists but is far from being understood and needs to be verified by extensive research.
The NO System as a Potential Therapeutic Target for Dental Pulp Repair and Regeneration
The control of pulp inflammation and the formation of reparative dentine (odontogenesis) and new blood vessels (angiogenesis) are vital determinants of whether good clinical outcomes are achieved in regenerative endodontic approaches.7,143 Many investigations have indicated the key roles of NO production due to cytokine-induced expression of iNOS in these processes: it acts as a proinflammatory or anti-inflammatory and odontoblastic differentiation mediator depending on its site and level of production in dental pulp tissue.9,49,144,145 Thus, promotion or suppression of the production or action of NO through specific approaches may be a strategy to control inflammation and promote tertiary dentin formation to maintain pulp vitality or prevent possible external root resorption and periapical bone loss (Fig. 2).48,49
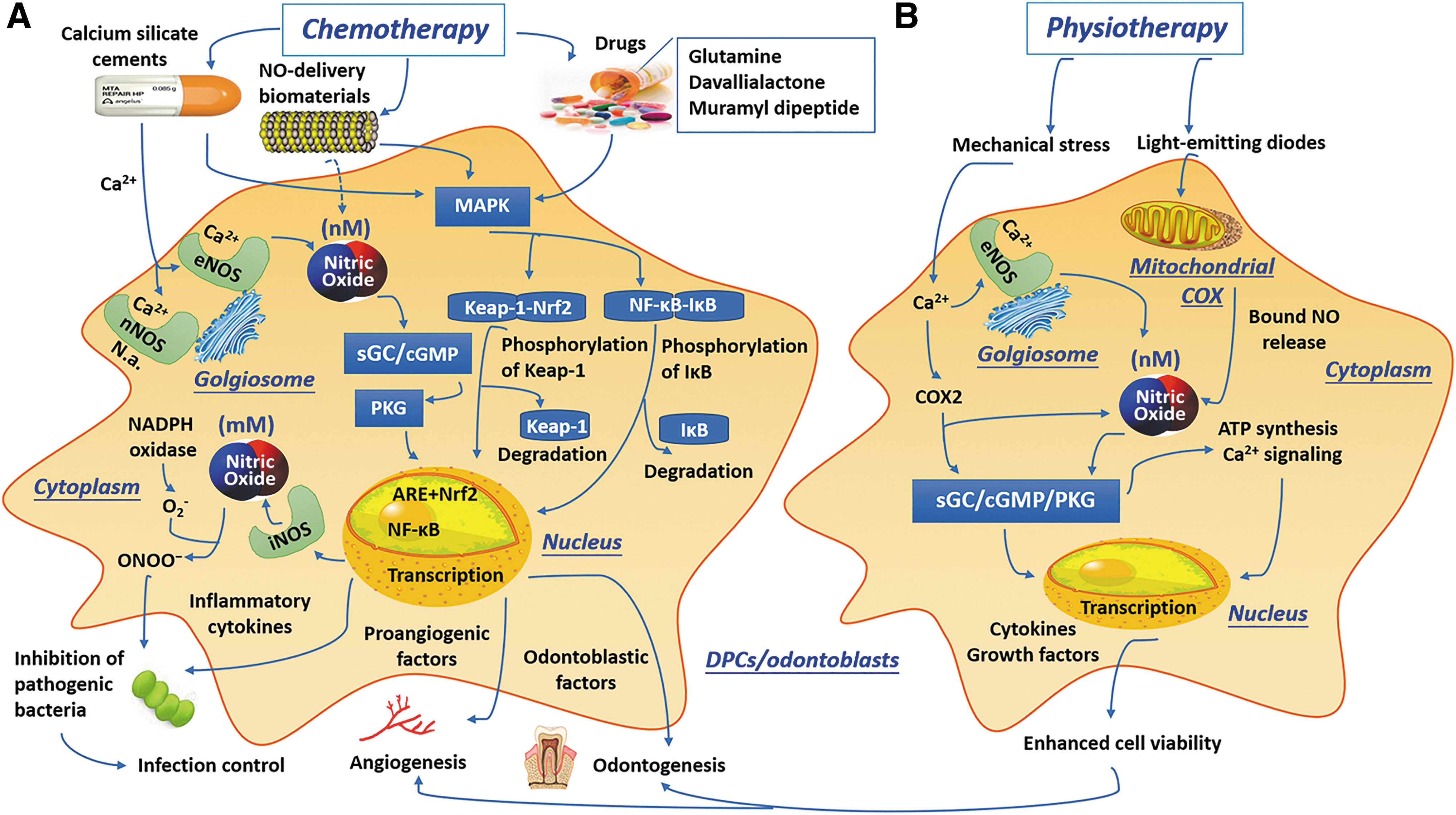
Schematic illustration of the clinical applications of NOS/NO system in dental pulp repair or regeneration and the involved signaling pathways.
Targeting NO signaling with NO donors, NO scavengers, and NOS inhibitors from NO-delivering biomaterials
With the progress in understanding the critical role of NO in tissue repair and regeneration under physiological and pathological conditions, such as cardiovascular, anti-inflammatory, antibacterial, antiparasitic, and anticancer processes, research focusing on NO-based therapies has rapidly increased. 146 Most studies on the biological activity of NO have used various hybrid drug compounds, such as NO donors, NO scavengers, and NOS inhibitors, to elevate or suppress NO levels or activity in the extracellular microenvironment to manipulate cell behaviors.
NO donors (viz. NO-releasing compounds) preserve NO in their molecular structures and evoke biological activity after decomposing. Some examples include organic nitrates, SNP, furoxans, and compounds containing the N(O)NO functional group (e.g., the diethylamine-NO complex or spermine-NO complex). 147 In numerous experimental studies, different NO donors have been adopted to monitor the biological effects of exogenous NO on DPCs, which have shown promotive effects on odontoblastic differentiation and subsequent reparative dentin formation in varying degrees (Table 2).15,49,53 For example, an in vivo analysis has revealed that a NO-releasing biomimetic nanomatrix gel incorporating antibiotics (ciprofloxacin and metronidazole) can promote tooth revascularization with apexification in healthy beagles. 48 Hence, NO donors may serve as a promising adjuvant in regenerative endodontics to promote the angiogenic and odontogenic events. However, only a few studies have explored the participation of exogenous NO in the cellular activities of DPCs, and the results are still controversial.15,49,124,148,149 The amounts of NO, cell type, and the states of cells may be pivotal determinants of the biological actions of NO. For example, low concentrations of NO tend to maintain the cell vitality although enhancing the expression of cytoprotective proteins such as HO-1; in these cases, NO is used as a signaling messenger. In contrast, high concentrations of NO appear to immediately cause cell damage before the production of cytoprotective proteins. 124
Cellular Responses to Exogenous Nitric Oxide and Involved Signal Pathways in Dental Pulp Cells/Odontoblasts
ALP, alkaline phosphatase; AMPK, adenosine monophosphate-activated protein kinase; Annexin V/PI, annexin V/propidium iodide double staining assay; Bax, bcl-2-like protein 4; Bcl-2, B cell lymphoma 2; BGLAP, bone gamma-carboxyglutamic acid-containinig protein; BrdU, bromodeoxyuridine; BSP, bone sialoprotein; COL1A1, type1 collagen; DMP1, dental matrix protein 1; DSPP, dentin sialophosphoprotein FCM, flow cytometry; HC, histochemistry; mTOR, mammalian target of rapamycin; MTS, 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium; MTT, 3,4,5-dimethylthiazol-2-yl-2,5-diphenyl tetrazolium bromide; qRT-PCR, quantitative real-time polymerase chain reaction; ROS, reactive oxygen species; RUNX2, runt-related transcription factor 2; SNAP, S-nitroso-N-acetyl-
Controlling bacterial progression and subsequent inflammatory reaction is a well-known prerequisite for pulp repair and regeneration. 7 NO is a highly potent radical with good antimicrobial property generated by many cell types, which acts as an inhibiter or killer on a variety of microorganisms under inflammatory conditions, such as E. faecalis and Treponema denticola. 48 Its antibacterial property is primarily derived from interactions with O2− that form highly cytotoxic ONOO− and S-nitrosylation of thiol residues that possesses strong oxidizing property and cytotoxicity. Thus far, macrophages and neutrophils have been the host cells most thoroughly investigated in terms of iNOS localization, and these cells possess antibacterial effect against aerobic and anaerobic bacteria through an iNOS-derived NO mechanism. 98 Likewise, human odontoblasts have also been reported to produce endogenous NO by synthesizing large amounts of iNOS, thus exhibiting antibacterial activity against caries-related microorganisms, for example, S. mutans, through TLR-2 activation. 46 In addition, recent studies reported that exogenous NO from a NO-releasing compound, such as pluronic F68-branched polyethylenimine-NONOates (F68-BPEI-NO) and NO-releasing silica nanoparticles, showed an adequate antimicrobial effect against specific periodontal pathogens with minimal cytotoxicity to human gingival fibroblasts.108,150
In contrast to NO donors, synthetic NO scavengers, such as 3-methyl-1-phenyl-2-pyrazolin-5-one (Edaravone) and 2-phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (PTIOs), can directly counteract NO through direct radical/radical coupling interactions with NO and may play beneficial roles in therapeutic strategies for oxidative stress-related diseases. 29 A recent study reported that PTIOs can negatively regulate the odontoblast differentiation of rat DPCs induced by the NO donor NOC-18. 49 However, similar studies on the anti-inflammatory, proangiogenic, and antibacterial roles of NO scavengers in dental pulp are rare and need to be conducted in the future.
NOS inhibitors are other important agents that can potentially be used to modulate some pathological disorders (e.g., mood disorders, ischemic stroke, arthritis, pancreatitis) by selectively targeting all NOS isoforms.58,146 Nonselective NOS inhibitors, such as
As previously discussed, NO can cause strong relaxation of the smooth muscle of blood vessels and can enhance vascular permeability to promote healing in inflamed pulp; however, if overproduction of NO continues for a long time, these phenomena might become harmful to the low-compliance pulp tissue. Thus, administration of an NOS- or iNOS-specific inhibitor may bring benefit on pulp healing or regeneration.3,72 Notably, NO may be induced by different pathways independent of endothelial cells, and NOS inhibitors also exert multiple nonspecific effects, including antagonism of mAChRs, generation of O2−, suppression of cytochrome reduction, and restraint of endothelium-independent relaxation induced by cyclic adenosine monophosphate (cAMP) or amiloride. 58 In addition, as mentioned above, iNOS-related NO can modulate the M1/M2 macrophage ratio 113 ; the reductions of this ratio may diminish the inflammatory activities and accelerate tissue repair process in the tooth suffering root resorption. 152 Thus, we may speculate that employing NO donors or NOS inhibitors may be a plausible therapeutic strategy to deal with orthodontic root resorption.
In summary, NO-based therapies using different types of NO donors, scavengers, and NOS inhibitors have shown some potential for clinical applications. However, few of them have been translated into practice because of drawbacks such as burst release, low payloads, and untargeted delivery. Currently, various biomaterials that have the potential to load and deliver NO in a sustained and/or controlled manner have been investigated that comprise hydrogels, nanoparticles and microspheres, nanoporous membrane, electrospun mats, and surface coatings; the combination of NO donors and these matrices may provide an occasion not only to control the release profiles of the donors but also to govern their cellular performances.24,32,153
Stimulation of NO signals through mineral trioxide aggregate-like cements
Biomaterials are foreign to the host, and the immune interactions between materials and tissues are very important for their biocompatibility evaluation and application. 16 Thus, copious basic and clinical evaluations of the immune response and odontoblastic differentiation of DPCs in response to different endodontic bioactive materials have been conducted.
As mentioned above, increased NO and PGE2 production through upregulation of iNOS and COX2 is a major inflammatory response in dental pulp.154,155 The NOS/NO system is the major regulator of many cellular functions, including survival, migration, differentiation, apoptosis, etc.; this system participates in various physiological actions, such as vasodilation, angiogenesis, nociception, antithrombosis, and scavenging of ONOO− (anti-inflammatory processes).11,156 IL-1 is a typical multifunctional cytokine involved in immune responses, hematopoiesis, and inflammation with regard to both innate and adaptive immunity 157 ; it is also a predominant cytokine in pulpal inflammation. 120 The most salient property of IL-1 (mainly IL-1α and IL-1β) is its ability to activate iNOS and COX2, which accounts for the production of NO and PGE2. 158 HO-1 possesses a cytoprotective effect against oxidative damage and inflammatory responses and thus may contribute to good clinical outcomes through a sequence of healing events.123,159 Hence, iNOS/NO, PGE2, IL-1 and HO-1 have frequently been selected as pivotal markers for evaluation of the biological characteristics/immunocompatibility of endodontic bioactive cements.
Mineral trioxide aggregate (MTA), mainly comprising calcium and silicate elements, has been developed in gray (GMTA) and white (WMTA) forms and is widely used as an bioactive endodontic material, including in various vital pulp therapies. 160 Human DPCs cultured on a calcium silicate cement substrate have been found to exhibit significant upregulation of proangiogenic factors (von Willebrand factor and angiopoietin-1 [vWF and Ang-1, respectively]) and to exhibit downstream eNOS protein expression and NO release. 161 Minamikawa et al. 121 found that MTA (ProRootMTA; Dentsply/Tulsa, Tulsa, OK) can increase iNOS and COX2 gene expression through activation of NF-κB signaling in rat DPCs, thereby stimulating the phosphorylation and degradation of IκB as well as the release of PGE2. Similar results obtained with human DPCs have indicated that MTA can increase COX2 expression. 162
Although calcium silicate ceramics are attracting increasing interest in the bone and dental fields, they still have some drawbacks, such as the inherent brittleness of ceramics, which prevents their mechanical properties from simulating those of cortical bone; their extended setting time; and their sandy consistency, which makes the materials difficult to handle.163,164 Hence, many strategies have been adopted to overcome these drawbacks and improve the biological performances of calcium silicate cements. For example, calcium silicate/gelatin composites with increased initial mechanical strength and biological functions that can effectively inhibit iNOS and IL-1 expression but activate IL-10 expression in human DPCs have been developed. 165 In endodontic practice, various calcium silicate-based materials (MTA-like materials), such as Portland cement (Brasseler USA, Savannah, GA), BioAggregate (Innovative Bioceramix, Vancouver, Canada), Biodentine (Septodont, Saint Maur des Fosses, France), and iRoot BP (Innovative Bioceramix), have been developed for use; the manufacturers claim these products have characteristics similar to those of MTA without exhibiting its shortcomings. 166
Bismuth oxide (Bi2O3; a radiopacifier)-containing Portland cement (BPC) is able to elevate nitrite (a marker of NO production) concentrations and iNOS mRNA expression that associates with HO-1 expression in DPCs. Interestingly, BPC-induced HO-1 expression in DPCs was found to attenuate its own cytotoxic effects. 167 Several novel calcium silicate cements (e.g., Angelus-MTA; Angelus, Londrina, Brazil; Ortho-MTA; BioMTA, Seoul, Korea) have been developed that use zirconium oxide instead of bismuth oxide to alleviate tooth discoloration from MTA; these all reduce the expression levels of iNOS mRNA, COX2 protein, proinflammatory mediator (such as NO, PGE2, TNF-α, IL-1β, IL-6, IL-8, and matrix metalloproteinase [MMP; MMP-1, MMP-2, MMP-8, and MMP-9]) mRNA, and intracellular ROS through ROS-dependent MAPK and Nrf2 activation and subsequent HO-1 expression. 168 However, one study observed higher IL-1 and iNOS levels in human DPCs cultured on two cements, that is, radiopaque dicalcium silicate cement and WMTA, than in those cultured on a tissue culture plate, possibly because of the initial high pH values, the heat produced by material setting reaction, and the generation of proinflammatory cytokines, such as IL-1 and IL-6, during the procedure.154,169
Although they exhibit various biological effects, nearly all MTA-like cements are highly biocompatible under odontogenic and angiogenic conditions, and we are still unable to recommend one specific material over the others. 170 MTA-like materials exert anti-/proinflammatory effects through efficiently regulating the production of NOS mRNA and protein (eNOS and iNOS), COX2 protein, NO and PGE2, and macrophage polarization that may contribute to their excellent clinical performance, at least in part.111,161,168,170–173 However, it is unclear which NOS isoforms and signaling pathways are related to this process. More detailed data related to the effects of various MTA-like materials (viz., calcium silicate cements) on DPCs can be gleaned from a review by Emara et al. 170
Other potential drugs that affect NO signals
Sufficient control of infection and survival of stem/progenitor cells are two primary goals of all regenerative endodontic procedures, but traditional agents such as sodium hypochlorite, Ca(OH)2, and antibiotic pastes have some drawbacks and cannot reliably achieve these two goals simultaneously. 7 For instance, the high pH of Ca(OH)2 makes it potentially cytotoxic and causes it to tend to dissolve soft tissues and impair macrophages phagocytosis of E. faecalis, thereby resulting in chronic inflammation and cell necrosis. 174 Thus, various newer agents or drugs that affects NO signals and have cytoprotective and anti-inflammatory effects may be potentially adopted in pulp regeneration therapy.
MMPs are known to degrade the extracellular matrix, thus affecting cell apoptosis, migration, differentiation, and inflammatory processes. MMP-3 has been reported to downregulate LPS-induced NO synthesis, inactivate or counteract proinflammatory mediators, and promote clearance of inflammatory cells; its application to injured pulp tissue can induce angiogenesis and wound healing,57,175 which suggests the potential utility of MMP-3 in the control and elimination of pulpal inflammation.
Amino acids are well documented to interact with NOS/NO in the context of inflammation and to participate in the modulation of immune activities. 134 Glutamine is a nonessential amino acid that is synthesized by organisms as needed. Defined levels of exogenous glutamine (10 mM) have been shown to modulate immunosuppressive properties of bone marrow mesenchymal stem cells (BMSCs). 176 Thus, relevant studies have been conducted to determine its role in pulp tissue activity and have reported that it can promote growth, migration, and differentiation in human DPCs through the BMP-2 and MAPK pathways. 177 In addition, glutamine can reduce LPS-induced iNOS and COX2 protein expression as well as the release of NO, PGE2, and other cytokines (viz. IL-1β, TNF-α, and IL-8) in a dose-dependent fashion, thereby exerting an anti-inflammatory effect by blocking the MAPK (p38, ERK, and JNK) pathways, with activation of MAPK phosphatase 1 (MKP-1) and subsequent inhibition of NF-κB translocation, in LPS-treated human DPCs. 103
Some natural compounds (e.g., davallialactone, sappanchalone, and pachymic acid) extracted from plants and fungi exhibit anti-inflammatory effects by stimulating NO signals and are now emerging as significant agents for pulpitis treatment. Davallialactone shows anti-inflammatory and antioxidant effects that decreases the level of inflammatory molecules such as iNOS and COX2 in LPS-induced human DPCs through inactivation of ERK1/2 followed by blockade of NF-κB translocation from the cytosol through the nuclear membrane. 128 Sappanchalone is a bioactive flavonoid isolated from Caesalpinia sappan L. that exhibits many biological activities, including neuroprotective and anti-inflammatory activities. In human DPCs, 5–40 μM sappanchalone can increase the protein expression of HO-1 and HO activity in a concentration-dependent manner through activation of JNK and nuclear translocation of Nrf2, which results in the suppression of LPS-induced inflammatory mediators, decreasing iNOS and COX2 protein expression, NO and PGE2 production, and IL-1β, IL-6, IL-12, and TNF-α levels. 125 Epigallocatechin gallate, a major green tea polyphenol, has various pharmacological effects on cell vitality. It has also been reported to serve as a scavenger of ROS and regulator of B cell lymphoma 2 (Bcl-2) family, and thus exert a protective effect against NO-induced apoptosis in human DPCs. 178 A more recent study suggested that Schisandrin C, a natural compound extracted from Schisandra chinensis, can suppress the LPS-induced formation of NO and other inflammatory mediators (IL-1β, TNF-α, ICAM-1, VCAM-1, MMP-2/9, and ROS) in human DPCs and thus has the potential to be used as an anti-inflammatory agent in the context of pulpitis to reduce inflammation and oxidation through downregulation of the MAPK (ERK1/2 and JNK) and NF-κB pathways and to promote mitochondrial biogenesis through upregulation of p-Akt- and Nrf-2-mediated HO-1 expression. 179 Therefore, the favorable anti-inflammatory abilities of these natural compounds might enable their use in treatments for multiple oral diseases such as pulpitis and periodontitis.
In addition, some specific drugs used for other diseases, such as bone tumors, immune diseases, and metabolic diseases (diabetes), have also emerged as potential agents for the control of dental pulp inflammation. For instance, zoledronic acid, a third-generation amino bisphosphonate that is used for the treatment of bone metastatic cancers at high doses, can modify immune cell profiles in rat dental pulp, thereby increasing the number of macrophages and the expression levels of some proinflammatory markers, such as iNOS, IL-1β, and TNF-α, independent of the NF-κB pathway. 180 Another example is metformin, a drug for treatment of type 2 diabetes, has been found to facilitate the healing of apical periodontitis by regulating the iNOS expression and NO production. 181
NOS activities and mechanical stress
In addition to the above chemotherapeutic strategies, physiotherapeutic strategies, such as mechanical stress and photodynamic therapy, have been considered as candidate approaches for pulp disease treatment. Mechanical loading is key to tissue growth, especially bone remodeling; fluid flow through the bone canalicular system generates shear stress, which stimulates the anabolic activity of osteoblasts and osteocytes that governs an adaptive process of bone healing and remodeling. 156 Hence, fluid shear stress within the physiological magnitude is usually applied by pulsating fluid flow in vitro to mimic in vivo loading through the lacunar/canalicular network on osteocytes. Relatively high fluid shear stress values, that is, those between 0.5 and 2 Pa, can result in the production and fluctuation of signal molecules such as intracellular Ca2+, eNOS-mediated NO and PGE2, which are required for cell anabolic responses; the latter two are also regulated by the mechanosensitive gene COX2. 182
Similar to bone cells, DPCs are also sensitive to mechanical stress, which can significantly promote cell proliferation and odontogenic differentiation. 183 Previous studies demonstrated that pulsating fluid flow can stimulate higher levels of NO, PGE2, and COX2 gene expression in mature DPCs than in immature DPCs within 5 min, suggesting the following two possibilities: (1) that the rapid stimulation of NO production results from the activity of eNOS (Ca2+-dependent NOS) but not iNOS and (2) that there is a relationship between cell maturation stage and cell mechanosensitivity.184,185 However, iNOS expression (typical of inflammatory conditions) in pulp subjected to orthodontic forces was present but had no obvious change compared with control dental pulp, which could be explained as a response to extractive stresses. 186 Moreover, mechanical irritants can provoke a range of inflammatory responses in dental pulp, such as the production of diverse cytokines and chemokines related with immune responses.6,184 Another illuminating case is that occlusal forces can promote periodontal ligament healing through inducing NOS expression and NO production.187,188 Thus, exertion of optimum mechanical stress (e.g., occlusal stimuli) on DPCs or odontoblasts combined with the use of specific bioactive materials [e.g., Ca(OH)2 and MTA] may be a possible method to stimulate dentin formation through activation of NOS/NO signaling.
Triggering of NO production by phototherapy in pulp inflammation
Phototherapy or low-level light therapy, also termed photodynamic therapy or photobiomodulation, involves the use of low-powered lasers in the far-red to near-infrared spectrum (600–1000 nm) or light-emitting diodes (LEDs) of <500 mW to reduce inflammatory reaction, relieve pain, and promote tissue repair and has emerged as the novel therapeutic method in a range of medical and health fields. 189 Although early studies identified mitochondrial cytochrome c oxidase (COX) in the electron transport chain as an endogenous photoreceptor for phototherapy, the molecular and cellular mechanisms underlying the effects of phototherapy remain unclear. It is said that the extracellular release of NO, ATP, or some specific growth factors may activate autocrine signaling such as the secretion of IL-1 by macrophages, resulting in the beneficial actions of phototherapy.190–192 iNOS/NO system may contribute to the mechanism of phototherapy by affecting mitochondrial physiology.31,193 Light can dissociate bound NO from mitochondrial COX, which restores oxygen consumption (because oxygen can rebind COX) and resumes respiratory chain activity, inducing ATP synthesis and Ca2+ signal. Recent investigations have demonstrated that phototherapy may increase the bioavailability of NO by releasing it from intracellular stores, mainly heme proteins.191,192 Notably, during cell stress and subsequent inflammatory defense response, high levels of NO exert an inhibitory effect on COX activity and thereby suppress tissue repair events.190–192
Phototherapy has been applied for the treatment of many oral conditions, including multiple oral mucosal diseases, temporomandibular joint disorders, orofacial pain, postextraction paresthesia, endodontic infections, periodontal diseases, caries, and peri-implantitis.194,195 With regard to dental pulp, most studies have stated that phototherapy can promote the viability of DPCs 196 and modulate the expression of cytokines, growth factors, and other inflammatory mediators. 195 A previous study indicated that 653 nm LED irradiation exhibited a promotive effect on cell growth and responses relevant to tissue repair in rat DPCs by significantly increasing ATP, NO, (likely not induced by NOS but rather released from COX), and mitochondrial metabolic activity. 190 In a later study on LPS-stressed human DPCs, an 855 nm infrared LED stimulation was shown to obviously suppress NO and ROS production, while human DPC metabolism was not significantly affected. 104 Hence, phototherapy can serve as a future clinical tool for biomodulation of inflammatory processes in pulp tissue favoring tissue repair.
Exploration of NOS/NO involved stem cell and gene therapy in regenerative endodontics
In addition to the abovementioned treatment methods, some NOS/NO-mediated therapeutic strategies for which relevant studies are lacking in dental pulp tissue may be applied to dental pulp repair and regeneration. For example, NO is believed to participate in the immunomodulation, epigenetic regulation, and multilineage differentiation of stem cells and thus has been used as a therapeutic agent for numerous diseases through its interaction with various stem cell types (e.g., neuronal stem cells in stroke neuron damage repair, endothelial progenitor cells in neural vessel repair, adipose-derived mesenchymal stem cells in calvarial defect repair, and BMSCs in multipotent tissue repair).107,197 In addition to external NO supply from exogenous NO donors and novel NO carriers, gene-based internal NO regulation through insertion of the NOS gene into various types of stem cells or suppression of NOS gene appears to be a plausible alternative to control NO production in situ and has been tested in several types of cardiovascular diseases and neurological disorders.197–199 However, there is no relevant research on dental pulp tissue. Thus, further work is required to understand the in vitro and in vivo effects and mechanisms of NOS gene-mediated cell therapy and to translate its benefits into clinical practice for regenerative endodontics.
Conclusion and Perspective
Much progress has been made toward characterizing the roles of the NOS/NO system in various physiological and pathological processes of dental pulp tissue, but we still lack a mechanistic understanding of how NOS/NO shifts between a promoter and inhibitor of tissue repair or regeneration during pulp inflammation. Several issues need to be addressed in future studies.
The biological significance of species-specific differences in NOS/NO signaling is not well understood. While different species (e.g., mice, rats, and humans) and cell types share similar properties in the chemistry of NO synthesis and the protein structure of iNOS, they exhibit diverse regulation patterns in iNOS expression. For instance, LPS and cytokine challenge can induce iNOS expression in murine endothelial cells, but is not effective in human endothelial cells.27,98 Most of the current knowledge regarding the functions of the NOS/NO system in dental pulp has been obtained from mice; hence, further work is needed to identify the specific characteristics of NOS/NO in human samples.
In general, NO shows antiapoptotic effects in many cell types at low concentrations, whereas it tends to exert cytotoxic effects at high concentrations. The opposing roles of NO in apoptosis mediate reciprocal regulatory effects in many human diseases. For example, its antiapoptotic effect is neuroprotective but tumor promoting, while its proapoptotic effect induces neurodegenerative diseases but inhibits tumorigenesis and tumor progression. 25 With regard to pulp tissue, the most puzzling question is how NO exerts dual effect on cell behaviors depending on its levels and cell lineages, such as DPCs, odontoblasts, endothelial cells, neurons, etc. Certainly, relevant investigations into responses should be conducted from the molecular and cellular levels to the whole-tissue and organ levels in vitro and in vivo.
During the development of dental caries, the invasion of cariogenic bacteria into tooth hard structures triggers immune and inflammatory responses in the underlying pulp tissue, the attenuation of which is believed to be a prerequisite for the regeneration of impaired pulp/dentine complex (Farges et al. 46 ). High rates of iNOS-mediated NO production play double roles in pulpitis. On the one hand, NO has beneficial effects that inhibit microbes, modulate immune responses, and prevent the formation of microvascular thrombi. On the other hand, NO has detrimental effects, as exaggerated generation of NO can affect blood homeostasis and cause toxicity toward pulpal tissues.44,200 Furthermore, local high NO-elicited excessive vasodilation and enhanced vascular hyperpermeability can raise the hydrostatic pressure inside low-compliance pulp tissue; sustained elevations in pressure can compress the pulpal venules, significantly damage pulpal perfusion, and even cause pulpal necrosis. 44 The transition between these opposing roles might depend on the local level of NO and its toxic derivatives, such as ONOO−, and the site of the pathological process, 8 but the exact mechanism of action remains unclear. Thus, from the perspectives of infectious diseases and wound healing in the context of dental pulp inflammation, five aspects of NO should be focused on in the future: (a) the different natures of NO-mediated inflammation in the dental pulp and in other parts of the body, (b) the concrete roles of NO at specific levels as a cytotoxic molecule from phagocytes in an antimicrobial system, (c) the exact nature of NO as an endothelium-derived relaxation factor during inflammation, (d) the potential role of NO as a regulator in odontoblastic activities of DPCs inside inflamed pulp tissue, and (e) the action of nonclassical NO signaling (e.g., protein S-nitrosylation) in pulpitis.
We should note the latent involvement of systemic diseases in pulp inflammation through stimulation of NOS/NO signals. Unhealthy lifestyles (e.g., smoking and alcoholism) and cardiovascular (hypertension) and metabolic diseases (diabetes), can impair the synthesis and effects of NO and result in systemic and regional circulatory disorders.12,201 The representative example is diabetes, which has profound effects on modification of structural components of dental pulp (e.g., decreasing melatonin expression) and the expression of inflammatory mediators; for example, it increases iNOS levels and causes uncontrolled NO production, which can negatively mediate diabetic complications through the promotion of oxidative/nitrosative stress.55,202
Studies on primary and immortalized cells from single donors have demonstrated the qualitative differences in some specific properties, such as pattern recognition receptor profiles, drug responsiveness, and pathogen-sensing expression. Primary DPCs cultured in vitro tend to undergo senescence after serial subcultivations, which makes it difficult to obtain reproducible results. 106 To deal with this problem, immortalized DPCs, which replicate readily and are easy to maintain in culture, should be used to enable convenient, comparable and cost-effective experimentation. In addition, the DPCs used in experiments should be isolated from at least three to five different donors and cultured separately before being used for subsequent analyses.
In conclusion, the NOS/NO system not only contributes to the regulation of physiological activities in the dental pulp but also plays crucial roles in initiating and mediating the immune/inflammatory responses triggered by external stimuli. Therefore, NOS and NO could serve as reasonable targets and tools for novel therapeutic strategies to avoid the irreversible damage on the dental pulp tissue and to promote the regeneration of dentin/pulp complex. Notably, due to its multifaceted effects, NO must be administered at a specific site with the right dose. Future evaluations should be toward to define NO-mediated potential beneficial interactions between inflammation and tissue healing or regeneration in human dental pulp and to explore the exact underlying mechanism(s) of action.
Footnotes
Disclosure Statement
The author has explicitly stated that there are no conflicts of interest in connection with this article.
Funding Information
This study was supported by the Guangdong Basic and Applied Basic Research Foundation (grant no. 2019A1515010072), and Fundamental Research Funds for the Central Universities (Grant no. 16ykpy01).