
Abstract
The design of cell-instructive biomaterials for tissue engineering and regenerative medicine is at a crossroads. Although the conventional tissue engineering approach is top-down (cells seeded to macroporous scaffolds and mature to form tissues), bottom-up tissue engineering strategies are becoming appealing. With such developments, we can study cell signaling events, thus enabling functional tissue assembly in physiologic and diseased models. Among many important signaling pathways, the Notch signaling pathway is the most diverse in its influence during tissue morphogenesis and repair following injury. Although Notch signaling is extensively studied in developmental biology and cancer biology, our knowledge of designing biomaterial-based Notch signaling platforms and incorporating Notch signaling components into engineered tissue systems is limited. By incorporating Notch signaling to tissue engineering scaffolds, we can direct cell-specific responses and improve engineered tissue maturation. This review will discuss recent progress in the development of Notch signaling biomaterials as a promising target to control cellular fate decisions, including the influences of ligand identity, biophysical material cues, ligand presentation strategies, and mechanotransduction. Notch signaling is consequently of interest to direct, control, and reprogram cellular behavior on a biomaterial surface. We anticipate that discussions in this article will allow for enhanced knowledge and insight into designing Notch targeted biomaterials for various tissue engineering and cell fate determinations.
Impact statement
Notch signaling is recognized as an important pathway in tissue engineering and regenerative medicine; however, there is no systematic review on this topic. The comprehensive review and perspectives presented here provide an in-depth discussion on ligand presentation strategies both in 2D and in 3D cell culture environments involving biomaterials/scaffolds. In addition, this review article provides insight into the challenges in designing cell surrogate biomaterials capable of providing Notch signals. To the best of the authors' knowledge, this is the first review relevant to the fields of tissue engineering.
Introduction
Over the past two decades, significant advances have been made in tissue engineering and regenerative medicine along with new biomaterial design, fabrication technologies, and cellular reprogramming. The discovery of induced pluripotent stem cells (iPSCs) has opened new frontiers to accelerate engineered human tissue fabrication both to treat disease and to model physiological processes.1,2 In tissue engineering and regenerative medicine strategies, cell–cell signaling molecules (e.g., growth factors) or cell–matrix signaling molecules (e.g., Arginine-Glycine-Aspartic acid [RGD] peptide or full-length extracellular matrix [ECM]-embedded proteins) are frequently utilized. Since tissue-resident cells are composed of multiple subpopulations of cells, heterotypic cell communication is critical for engineered tissue function. Cells continuously send and receive signals that are essential for development, homeostasis, and repair of various tissues and organs. They are able to adjust to their microenvironment and communicate with each other through a complex network of signaling pathways.
In general, cells receive information through four types of signaling—autocrine, endocrine, paracrine, and juxtacrine (Fig. 1A–D). In tissue engineering, cells signal predominantly by paracrine or juxtacrine signaling. 3 The difference between paracrine and juxtacrine signaling is the mode of ligand presentation.4,5 In paracrine signaling, the ligand is secreted by one type of cell and is released into the neighboring target cells as a diffusible soluble factor. In juxtacrine signaling, the ligand is anchored on the signal-sending cell surface to bind the receptor on another cell, thus requiring cell–cell contact for proper function.
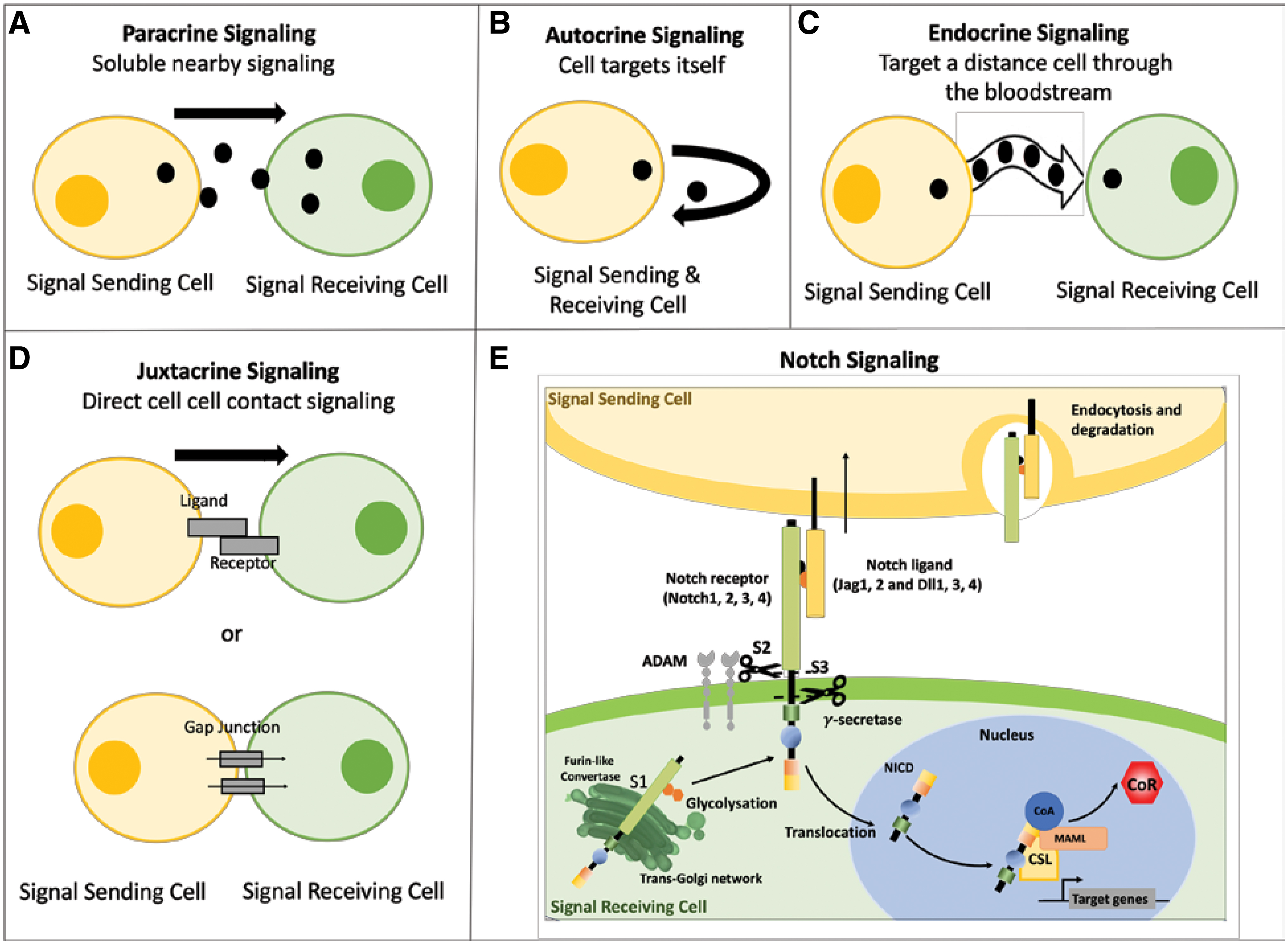
Cell signaling types and an overview of the canonical Notch signaling cascade. There are four types of cell signaling mechanisms identified based on the distance the signal has to travel and how the signal is delivered.
Notch signaling is a critical heterotypic juxtacrine cell-to-cell signaling pathway that is active in numerous cell fate decisions, including development, tissue maintenance, homeostasis, as well as disease progression.6–8 Its various tissue-specific roles allow for Notch signaling to be a promising target for cellular control. Notch signaling is an evolutionarily conserved mechanism that is regulated by interactions with transmembrane proteins of the Jagged (Jag) or Delta-like (Dll) family of ligands. Mammalian tissues express various combinations of four Notch receptors (Notch1, Notch2, Notch3, and Notch4) and five Notch ligands (Jag1, Jag2, Dll1, Dll3, and Dll4), and signaling is activated through three sequential cleavages, named as S1, S2, and S3 cleavages. In S1, Notch is cleaved into a heterodimer by Furin-like convertase, which undergoes O-fucosylation by O-Fucosyltransferase and glycosylated by Fringe in the Golgi before the receptor is transported to the cell membrane. This processing step controls the abundance of Notch receptors at the cell surface.
The binding interaction between a specific Notch receptor with a corresponding ligand initiates the regulated intramembrane proteolysis, which results in a conformational change of the Notch extracellular domain (NECD) of the receptor. This conformational change exposes an S2 cleavage site for the metalloprotease ADAM17/TACE to initiate. In the absence of ligand binding, the Notch receptor is maintained in an autoinhibited and protease-resistant state. 9 There is extensive evidence implicating NECD endocytosis in Notch signaling and degradation by the signal sending cell; however, the mechanism and purpose are still highly debated.9–13 Full activation of Notch is achieved upon S3 cleavage of the Notch extracellular truncation (NEXT) by presenilin, the proteolytic subunit of the γ-secretase complex, which cleaves the Notch intracellular domain (NICD). Once released from the plasma membrane, the NICD translocates into the nucleus, where it binds to the transcriptional repressor RBPJ (recombination signal-binding protein for immunoglobulin κ J region also known as CSL and CBF1). Finally, the NICD activator complex promotes transcription of downstream gene targets, including Hairy Enhancer of Split (HES) and HES related with YRPW motive (HEY), as well as various downstream functional genes and proteins14,15 (Fig. 1E).
Notch Signaling in Developmental Biology
The strategies of tissue engineering and regenerative medicine are inspired by developmental biology and pathophysiology principles. 16 Notch signaling is simple in design with few core components, as shown in Figure 1E; however, it is very complex from a regulation perspective as it affects numerous distinct cell fate decisions that are important in the development of many tissues.17–19 To utilize Notch signaling during development as inspiration for engineered tissues, an understanding of ligand-receptor specificity is necessary.20–22 Unlike other signaling pathways that are enzymatically amplified, Notch signaling instead depends on stoichiometric receptor-ligand interactions to activate it, and any imbalance may inhibit the process.18,21 Distribution of the Notch signaling components in different tissues varies considerably, thus interpreting the context-specific effects of Notch will ultimately require that we understand the wiring of the regulatory networks in which it operates 7 and the importance of specific ligand-receptor pairings to develop new therapeutic targets. Understanding the role of Notch in tissue specification during development and disease state is beneficial to translate these mechanisms into tissue engineering. 23 Notch signaling plays a role in several tissues and organs, including the heart, vasculature, bone, liver, breast, ear, skin, and lymphatic systems, demonstrating its extensive and diverse control of many cell types. Although the principle of tissue engineering can be equally applied to the above-listed tissues, the vast majority of engineered tissue studies to date is in cardiovascular (cardiac, vascular, and heart valves) and musculoskeletal (bone and cartilage). In view of this, the importance of Notch signaling in these tissues is the main focus of the next section, to highlight important biological processes driven by Notch that can be leveraged by tissue engineers.
Notch modulation in vasculogenesis and angiogenesis
Vasculogenesis is the differentiation of precursor cells (angioblasts) into endothelial cells (ECs) and the de novo formation of a primitive vascular network, whereas angiogenesis refers to the growth of new capillaries from preexisting blood vessels. Notch signaling is critical in both vasculogenesis during development and in angiogenesis.24,25 It protects the endothelium, 26 controls vascular smooth muscle cell (VSMC) phenotype,27–30 promotes neoangiogenesis,31–33 tip–stalk cell patterning, 34 and regulates arteriovenous specification.35,36 The various roles of Notch signaling in the vasculature, including its influence on ECs and smooth muscle cells (SMCs), have been reviewed previously.8,37 Herein we highlight important Notch receptor-ligand pairings in the vasculature. VSMCs dominantly express Notch1, Notch2, and Notch3 receptors, while ECs express Notch ligands Jag1, 2, and Dll4 and to some extent Dll1 in the remodeling vasculature.38,39 The context in which these ligand and receptors occur is essential; however, the anatomical location within the vascular bed and the associated physiological forces (flow and stress) are likely to impact the context-dependent Notch activation. 31
The development of the vascular system involves tip cell selection, sprout formation, tip cell migration, stalk cell proliferation, and ultimately vascular stabilization, 40 which are collectively influenced by Notch signaling. Vascular sprouting is guided by the migration of tip cells in response to a matrix-bound vascular endothelial growth factor (VEGF) gradient,34,41 with Dll4 acting downstream as a negative regulator.42,43 Tip–stalk cell fate plays a significant role in angiogenesis since tip cells direct new blood vessel growth. Interestingly, the role of Notch in both tip and stalk cells is evident within the distribution patterns of Notch signaling ligands and receptors. VEGF signaling induces Dll4 in tip cells, and tip cells then suppress tip–cell features in adjacent stalk cells through Dll4/Notch-mediated lateral inhibition. 34 Simultaneously Jag1 antagonizes Dll4-mediated Notch activation in stalk cells to increase tip cell number, which consequently enhances vessel sprouting. 44 Hence it is this Jag1 and Dll4 “salt-and-pepper” pattern that dictates the tip–stalk cell phenotypes within this niche.
The function of Notch signaling varies greatly depending on the location of the vascular bed and also the type of the vessel. The arteriovenous specification is established early in development through a variety of transcription factors. A key Notch-defining factor in this specification is the Notch3 receptor found in arterial SMCs, which is notably absent in veins, and Jag1 expression by ECs is responsible for this maturation. The transcription factors Foxc1 and Foxc2 and VEGF signaling are primarily responsible for arterial fate36,45; the upregulation of these transcription factors results in increased expression of Dll4. In contrast, vein identity is regulated by the repression of Notch1. 36
Recruitment of mural cells (VSMC and pericytes) and the formation of a fully endothelialized lumen are hallmarks of arterial vessel maturation during development. 36 Upon activation of Notch in VSMC, ligand-receptor signaling is initiated throughout the VSMC lamellae by the process of lateral induction. The propagation of Notch signaling is crucial for regulating VSMC phenotype throughout the vascular wall and, hence, is a critical phenomenon for inducing differentiation of the complete VSMC layer toward the homeostatic contractile phenotype. 46 Notch3 targeting is more prominent for SMC control and differentiation27,47,48; Notch1, in contrast, has been shown to regulate EC metabolism, proliferation, and monolayer regeneration. 49 Communication between ECs and SMCs in the vascular wall is essential; thus, diminished Jag1 expression in ECs leads to abnormal smooth muscle development. Endothelial expression of Jag1 is required for activation of Notch3 on VSMC maturation, differentiation, and contraction. 50
The impact of physiological forces within the vascular system on Notch signaling becomes apparent when considering the ability of the endothelium to respond to force patterning. ECs modulate its Notch component expression in response to hemodynamic forces. For instance, Notch1 activation in EC is sensitive to blood flow, where high shear stress has a critical role in the acquisition and maintenance of arterial identity through its role in endothelial quiescence. 51 The indication that EC appears to respond to predetermined arterial or venous patterns is evident through current research, and there is a certain level of plasticity cues of the local niche suggested to be imposed by physical forces, such as hemodynamics. 52 Taken together, ligand specificity and selective Notch activation regulate differential phenotypes and function within the vascular tissue.46,53,54 Understanding tissue distribution of Notch signaling components is highly relevant to translate them into tissue engineering and regenerative medicine strategies.
Notch modulation in cardiac tissue specification and repair
Notch signaling also has a critical role in both cardiac development and cardiac repair. Notch signaling is very active during embryonic cardiac development and has roles in both the myocardium and the endocardium. Notch regulates ventricular trabeculation, 55 cardiomyocyte differentiation and proliferation, 56 and valve formation. 57 We do not delve too much into Notch-mediated cardiac development, disease, and regeneration as it has been recently reviewed58–61 ; however, we provide a brief discussion relevant to tissue engineering. Notch1 and Jag1 are expressed in immature cardiomyocytes to protect them from apoptosis and stimulate proliferation and differentiation into fully differentiated and beating phenotypes.56,62,63 Although much attention is focused on the role of Notch in the myocardium, there is an essential function of Notch in the endocardium. Early endocardial Notch activity contributes to the patterning of the embryonic endocardium for the development of valve and chamber formation, 60 and then later, it regulates signaling critical for outflow tract valve morphogenesis64,65 and ventricular chamber development. 66 The endocardium is a specialized endothelial lining that closely juxtaposes to the myocardium to communicate. At a cellular level, Notch1 expression is downregulated in the cardiomyocyte lineage and is enriched in the endocardial lineage, corresponding to antimyogenic activity and its functional requirement in the endocardium.58,67 Notch plays a critical role in endocardium–myocardium communication that underlies trabeculation 61 ; Dll4 is found in the endocardium where trabeculae are formed. 68 In contrast, Jag1 is expressed in the cardiomyocytes forming the trabeculae and is also present in the compact myocardium. Although Notch is required for trabeculation, it is not active in the ventricular endocardium during trabeculation, but otherwise controls trabeculation through intermediate signaling pathways. 69 The Dll4-Notch1 pairing actively regulates gene expression, encoding signals that connect the ventricular endocardium and myocardium, but also promotes cardiomyocyte proliferation, differentiation, and metabolism.58,70
Interestingly, Notch signaling is silenced in the postnatal heart and only restored after injury. Notch1 has been identified as a critical determinant receptor in the heart, as it interacts with various ligands in cardiac repair following injury. 59 Because Notch signaling is directly linked to repairing mechanisms to drive regeneration, 71 rather than scar formation, it is an interesting target for therapeutic cardiac injury intervention. The inhibition of endothelial Dll4 in the heart resulted in impaired fatty transport, leading to metabolic and vascular remodeling. 70 The upregulation of Notch1 in a hypertrophic heart controls the adaptive response of the heart to stress, which limits the extent of hypertrophic response and contributes to cardiomyocyte survival. 72 This demonstrates the cardioprotective role of Notch1, controlling the response to injury in the adult heart.
Notch modulation in the developing bone and repair
Skeletal tissues, specifically bone, is another tissue where Notch signaling plays an important role during development and disorder.73,74 The roles of Notch signaling include control of bone regeneration,75,76 skeletal remodeling, bone fracture repair,77,78 congenital bone defects, and malignancies of the bone. 79 Since Notch is tissue specific, Notch directed cellular response is dictated by the specific cells targeted and its stage of maturation. A coordinated effort by both chondrocytes and osteoblasts during embryogenesis leads to skeletal tissue specification. It is then continuously regenerated by the action of bone-forming osteoblasts and bone-resorbing osteoclasts to maintain homeostasis.
In early-stage skeletal development, Notch activation inhibits cell differentiation and causes cancellous bone osteopenia because of impaired bone formation. 80 Notch ligands, specifically Jag1, are involved in cell commitment; Notch inhibitors can control intramembranous ossification where mesenchymal tissue is replaced directly by bone through osteoblastogenesis. 81 Although initial studies have implicated both Notch1 and Notch2 receptors in the regulation of osteoblastogenesis, 82 subsequent studies concluded that Notch2, instead of Notch1, plays the predominant role. 83 Cell commitment becomes an important factor in Notch control since the activation of Notch signaling inhibits terminal differentiation of osteoblast progenitors (through Jag1) but does not affect mature osteoblasts. 76 Synergistically, osteoclasts work with osteoblasts to control bone homeostasis, and this is influenced by Notch. Osteoclastogenesis from macrophage precursors is similarly regulated by Notch signaling; however, Notch1 is the predominant receptor. Similar to the vasculature Dll and Jag ligands have opposing functions on bone tissue response but are again context dependent. The Notch1/Jag1 axis suppresses osteoclastogenesis, whereas the Notch2/Dll1 axis promotes the effect. 84 In bone remodeling, the disruption of Notch in osteocyte differentiation in the mineralized matrix can affect calcium phosphate deposition 75 and alter crystal mineral structures, linking Notch with dysregulation and its role in bone mineralization. 74 Collectively, it is evident that Notch signaling regulates several aspects that control osteoclasts, osteoblasts, and osteocytes, which are critical in bone tissue.
Notch signaling is also important in bone repair following injury. Fracture healing is caused by endochondral ossification and intramembranous bone formation. 81 Therefore, achieving a balance between osteoblastogenesis and osteoclastogenesis, alongside with the infiltration of blood vessels, mediates proper fracture healing. The interaction of mesenchymal, hematopoietic, and vascular cells plays a Notch-dependent role. For example, in bone marrow-derived mesenchymal stem cells (MSCs) reduced Notch signaling demonstrated depletion of this population, leading to fracture nonunion.74,78 The role of Notch in fracture healing is complex, with both upregulation and downregulation playing a part depending on ligand-receptor selectivity, and needs more research to fully understand the mechanisms. To highlight ligand-receptor specificity, Jag1 delivery has been shown to induce osteoblast commitment, 76 mineralization deposition, 75 and promote healing of craniofacial and appendicular defects. 85 In contrast, disruption of Jag1 signaling has proven to decrease femoral trabecular bone formation, 85 accelerate cartilage regeneration, 86 and decrease joint inflammation. 86
Diseases and disorders associated with Notch dysregulation
Notch signaling has distinct and diverse biological roles in many tissues; therefore, it is not surprising that its dysregulation has been implicated in human diseases and disorders. 87 In many cases, mutations at the genetic level are attributed to disease and disorder; however, in others, the cause of the disease is less direct and associated with improper ligand-receptor distribution or dysregulation of receptor-ligand activation complexes. Both loss- and gain-of-function activity of Notch signaling are unique to each disease/disorder. Table 1 summarizes the roles of Notch signaling and Notch related diseases/disorders in selected tissues relevant to tissue engineering (the list is not exhaustive). Although Notch signaling may not be the primary cause for these diseases/disorders, Notch has a role in each of these diseases and could be a useful therapeutic target.
Diseases and Disorders Driven by Notch Signaling
EC, endothelial cell; SMC, smooth muscle cell; VSMC, vascular smooth muscle cell.
Notch Therapeutic Targets
Understanding the Notch signaling pathway and how Notch affects both normal tissue function, as well as disease and disorder, allows us to design various therapies harnessing the innate Notch behavior and response. These therapies can be specifically designed to be cell or tissue specific based on biological principles harnessed. Potential avenues for Notch intervention include controlling ligand–receptor interactions, modulating receptor processing, targeting of intracellular trafficking and downstream effectors, and integrating signaling cross talk between other signaling pathways.
Targeting ligand–receptor interactions
To utilize Notch signaling during development as inspiration for engineered tissues, an understanding of ligand–receptor binding and specificity is necessary20–22 ; therefore, one approach is to modulate the ligand–receptor interaction. As stoichiometric ligand–receptor interactions are necessary for pathway modulation, a distinction between cis- and trans-interactions among the ligands and receptors becomes crucial. 20 All ligands, except for Dll3, can activate receptors in trans 6 and trans-activation, whereby the signaling ligand and its receptor are distinct, but contacting cells is the most common mechanism. Delivery of Notch ligands in their soluble form will not activate Notch signaling and rather inhibits activation.28,112,133
An alternative strategy is if both the ligand and the receptor are expressed on the same cell surface. There are, in fact, cells that express both the Notch receptor and the ligand. As an example, in embryonic rat VSMCs, Notch3 and Jag1 are expressed in equal abundance. 134 Despite this, Notch signaling was not activated when these cells were cultured alone or cocultured with Jag1 presenting ECs. 134 Depending on the relative abundance of receptor to its ligand on the same cell surface, it can either be a ligand presenting (signal sending) or signal-receiving cell. However, when the Notch receptor binds to a ligand present on the same cell, signaling is inhibited. This phenomenon of cis-inhibition is an emerging feature of Notch signaling and relies on the relative abundance of the ligand and receptor.135,136 Although it remains to be elucidated further, cis-activation is implied when trans-ligands are effectively competing with cis-ligands for a common pool of Notch receptors. 136 Currently, Notch therapies targeting Notch ligand–receptor selectivity and binding include the use of binding proteins,137,138 monoclonal antibodies (mAbs),139–141 and engineered Notch decoy ligands.142,143 These strategies are used to preferentially block Dll or Jag type binding specific binding. For example, in engineered soluble peptides composed of different Notch EGF repeats 11-24 (N11–N24) fused to IgG γ-heavy chain, the Notch decoy N11-13 preferentially blocked Dll-type ligands, whereas N10-24 was specific to Jag-type inhibiton. 142 The use of soluble Notch factors may also be used as a binding inhibitory mechanism.28,112,133
Controlling Notch receptor processing
Notch receptor processing is another avenue that can be harnessed for enhancing or inhibiting proteolytic cleavages within the signaling cascade to control various cellular responses. S1 cleavage is controlled by Furin convertase; therefore, Furin inhibitors are ideal targets to control the abundance of receptors on the cell surface. Notch2 is insensitive to Furin inhibitors, whereas the surface expression of Notch1 is mildly reduced when S1 processing is impeded. 144 Furin convertase targeting has been suggested to direct but not inhibit signaling and has some other target proteins such as transforming growth factor β1 (TGFβ1); therefore, there is an increased risk for off-target effects 6 and is not advantageous. The second cleavage found in the Notch cascade is the S2 cleavage, which is engaged upon ligand–receptor binding and is controlled by ADAM10 or ADAM17. ADAM inhibition is a possible therapeutic target for ligand processing and Notch modulation 145 ; however, off-target risks remain. 6 The third receptor processing step occurs at the S3 cleavage site by γ-secretase and has been suggested as a possible avenue for intervention. Gamma-secretase inhibitors (GSIs) are used to inhibit S3 cleavage, attenuating the response of all Notch receptors. Targeting through GSIs diminishes the release of Notch from the plasma membrane and subsequent generation of NICD. The effectiveness of GSI treatments (e.g., DAPT) has shown promising results, and some have advanced into clinical trials, 146 including its use as a tumor suppressor in cancer therapy.147,148 Limitations of GSI treatments to inhibit the effects of Notch signaling include low treatment effectiveness, toxicity, and off-target effects blocking numerous other protein functions, 149 suggesting a need for additional therapeutic S3 cleavage approaches. mAb inhibitors are potentially advantageous due to their specificity allowing for the targeting of individual Notch receptors. mAbs have been directed toward Notch receptors, both targeting the extracellular negative regulatory region or blocking the conformation change that allows the ADAM protease cleavage to occur, and clinical attempts have been made.150,151 The process of endocytosis and endosomal sorting of Notch receptors is another therapeutic target through membrane-transport networks. Because of several excellent reviews on this topic,9,152–154 it is not discussed here.
Notch targeting through signaling cross talk
The ability for Notch signaling components to interact, impact, and cross talk with multiple signaling pathways adds another opportunity in controlling Notch signaling and developing new therapies. It is clear that Notch signaling is not an isolated pathway and interacts with signaling pathways mediated by TGFβ/bone morphogenic proteins (BMPs),155–158 VEGF, 104 Wnt, 159 and YAP/TAZ. 160 Many proteins have been identified that interact with the Notch ICD (these have been summarized previously 161 ). Although our current understanding of the molecular mechanisms involved in Notch cross talk is limited and emerging, it is evident that the pathways are integrated and can cause an attenuated or amplified behavior with a purpose to influence tissue specification. This integration is interesting because it might be able to explain the initial asymmetries found in Notch activity before ligand–receptor binding.
TGFβ/BMP signal convergence with Notch signaling is a well-established cross-talk interaction. Notch is known to have an influence controlling the TGFβ pathway with the expression of R-Smads and the synergistic regulation and interactions with the NICD and Smad3 to regulate a subset of Smad3 target genes.126,134 In addition, the integration of Notch and Smad1/5 signaling might be important. 34 These interactions, however, are cell dependent; in some contexts such as within keratinocytes and epithelial cells, there is a cooperative amplified behavior, and conversely, in other cases, the NICD was found to antagonize TGFβ growth arrest and transcription. 162 Although other pathways, for example, YAP/TAZ interactions have been studied and hypothesized to be linked, for example, with Notch related to the Hippo pathway signaling,163,164 a strong mechanistic insight has not been given as to the exact interactions of these related pathways. The cooperative effort and cross talk of various signaling pathways with Notch have been demonstrated in many cells and tissues, including cardiac tissues,57,61 epithelial cells, 57 in the epidermis, 165 angiogeneses,34,165 and myogenic differentiation162,166 to name a few. It is evident that Notch interacts and cross talks with various signaling pathways, and the response frequently differs between cell types and tissues. A further understanding of the mechanisms present in this interaction is needed to further understand from a regulation perspective of how this information can be used in various tissue-engineered therapies. Notch signaling alone or combined interaction with other pathways could be harnessed to create proper homeostasis within various tissues and similarly regulate physiological and pathological processes.
Notch Signaling in the Context of Tissue Engineering
Extending developmental biology principles and mechanisms to engineer tissues is important to recapitulate native cell–cell and cell–matrix interactions. Tissue engineering and developmental biology can provide complementary perspectives for reverse engineering of biomimetic tissues and to advance functional material development. 23 Tissue-engineered substitutes are not expected to completely mimic morphogenesis, but we can attempt to recapitulate key developmental signaling pathways to accelerate in vitro tissue formation. Despite the multicellular nature of tissues (e.g., osteoblasts and osteoclasts in bone, VSMCs and ECs in vascular tissues, or cardiomyocytes and fibroblasts in cardiac tissues), tissue engineering strategies often focus on seeding the main cellular component of a particular tissue to scaffolds and culture them for tissue maturation. For instance, VSMCs are seeded to scaffolds and cultured in vascular tissue engineering, osteoblasts in bone tissue engineering, and cardiomyocytes in cardiac tissue engineering. Given the role of heterotypic Notch signaling in these tissues, the signal sending cell (ligand-bearing cell) is often missing, caused by injury or disease. The main question is then: how can we present the Notch ligands to direct cellular response? One strategy is to present it as a soluble factor either by adding it together with culture media or using a delivery system. Indeed, the use of biomaterials for delivering soluble ligands to cells and tissues is not new. The soluble factor is loaded to a biomaterial and released either by diffusion or by cleavage, delivering the soluble factor in a paracrine manner. Although the immobilization of paracrine signaling ligands to biomaterial surfaces is common, it is not required from a signaling point of view but may help slow the release or increase cellular accessibility.
For Notch signaling, which is juxtacrine in nature, the ligand must be anchored on the signal sending cell and requires cell–cell contact for proper function and signal propagation. As stated earlier, soluble ligands do not activate Notch signaling.28,112,133 Embedded and immobilized delivery of Notch ligands through a biomaterial surface facilitates juxtacrine cell–cell signaling found in the native tissues. Engineered biomaterials can be introduced to take on the role of the signal-sending cell and act as cell-surrogate biomaterials to replicate the cell–cell contact needed for signal propagation allowing for better spatial and temporal control of delivery. From developmental biology, we can learn the various effective ligand–receptor pairings and proper regulation mechanisms that can be utilized to design more functional engineered tissues. Up until now, specific Notch targets and the influence of ligand–receptor selectivity have been suggested as a useful element to control when designing tissue-engineered substitutes (Fig. 2A-i). The remainder of this review will talk about various other elements to consider during Notch biomaterial design, including ligand presentation strategies, biomaterial topographical cues, spatial/temporal cues, and mechanotransduction (Fig. 2Aii–v). By targeting one or a combination of these elements, it is possible to better control and direct cellular behavior in biomaterial-based systems, which may be superior to soluble factors.
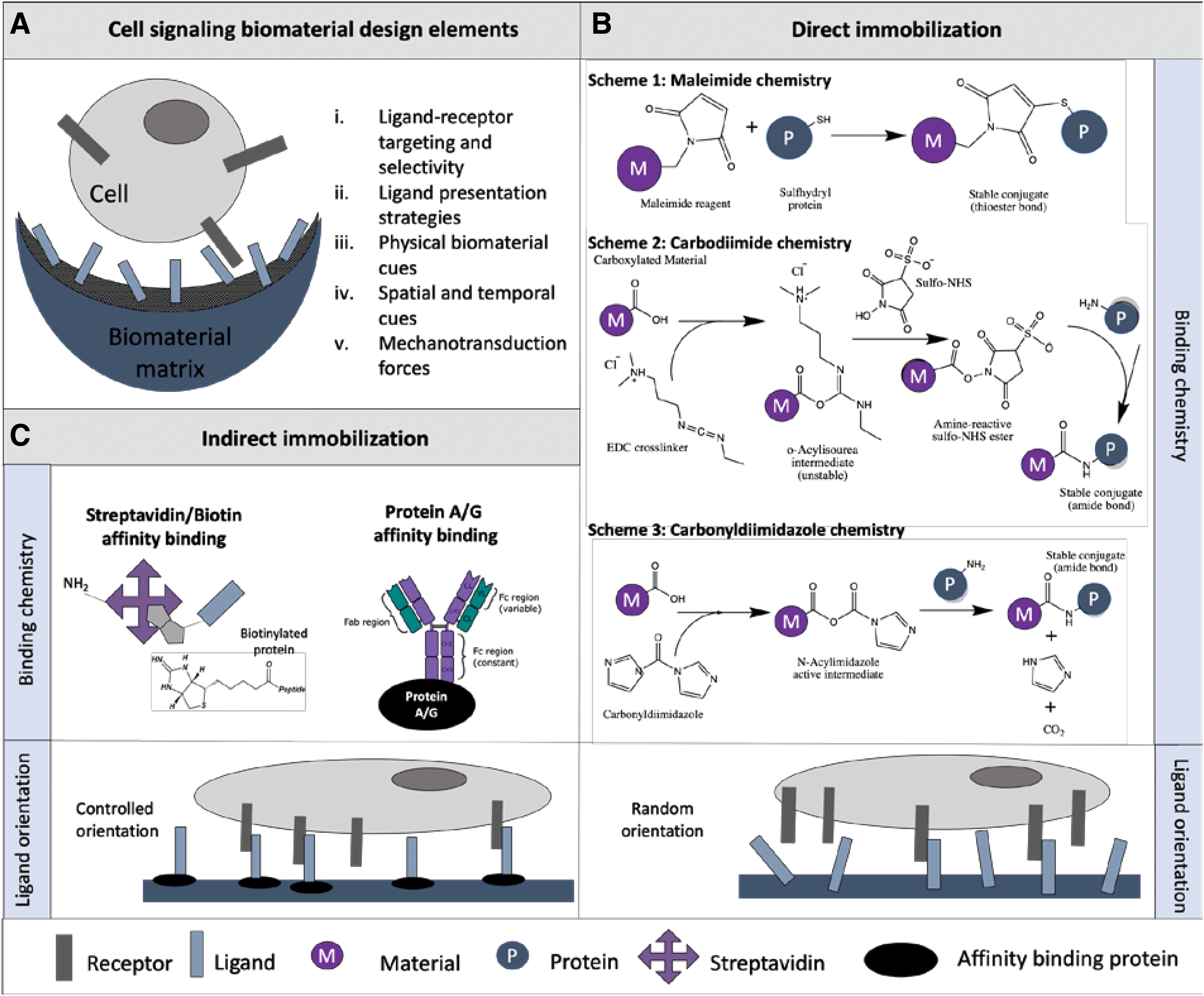
Notch ligand presentation strategies for cell signaling biomaterial design.
Relevance of Notch therapeutics to tissue engineering and regenerative medicine
Most Notch targets to date are focussed on cancer therapies, but there are other applications in tissue engineering and regenerative medicine (Fig. 3). Notch signaling can be amplified or attenuated to differentiate stem cells into a target lineage for regenerative purposes. Delivery of Notch ligand Jag1 to full-thickness wounds significantly enhances wound healing 167 suggesting the need to incorporate the ligand to wound dressing biomaterials. Activation of Notch1 in injectable cardiomyocytes enhances cardiac repair following infarction. Delivery of Jag1-containing hydrogels also counteracts cardiac fibrosis, primarily through inhibition of myofibroblast differentiation, thus accelerating cardiac repair. 168 Impaired VSMC differentiation following stent deployment and the subsequent excessive proliferation causes restenosis due to endothelial denudation; therefore, Notch ligand delivery using stents could be useful to modulate restenosis. 169 Similarly, activation of Notch signaling is needed to repair nonunion bone fracture. 78 Instead of activation, attenuation of Notch by either soluble ligands or by one of S1–S3 cleavage inhibitors promotes injectable cell survival and chondrogenic differentiation, thereby increasing the therapeutic potential. 86 These illustrative examples underscore the relevance of Notch therapeutics to tissue engineering and regenerative medicine and the need for Notch activation through appropriate ligand presentation strategies.
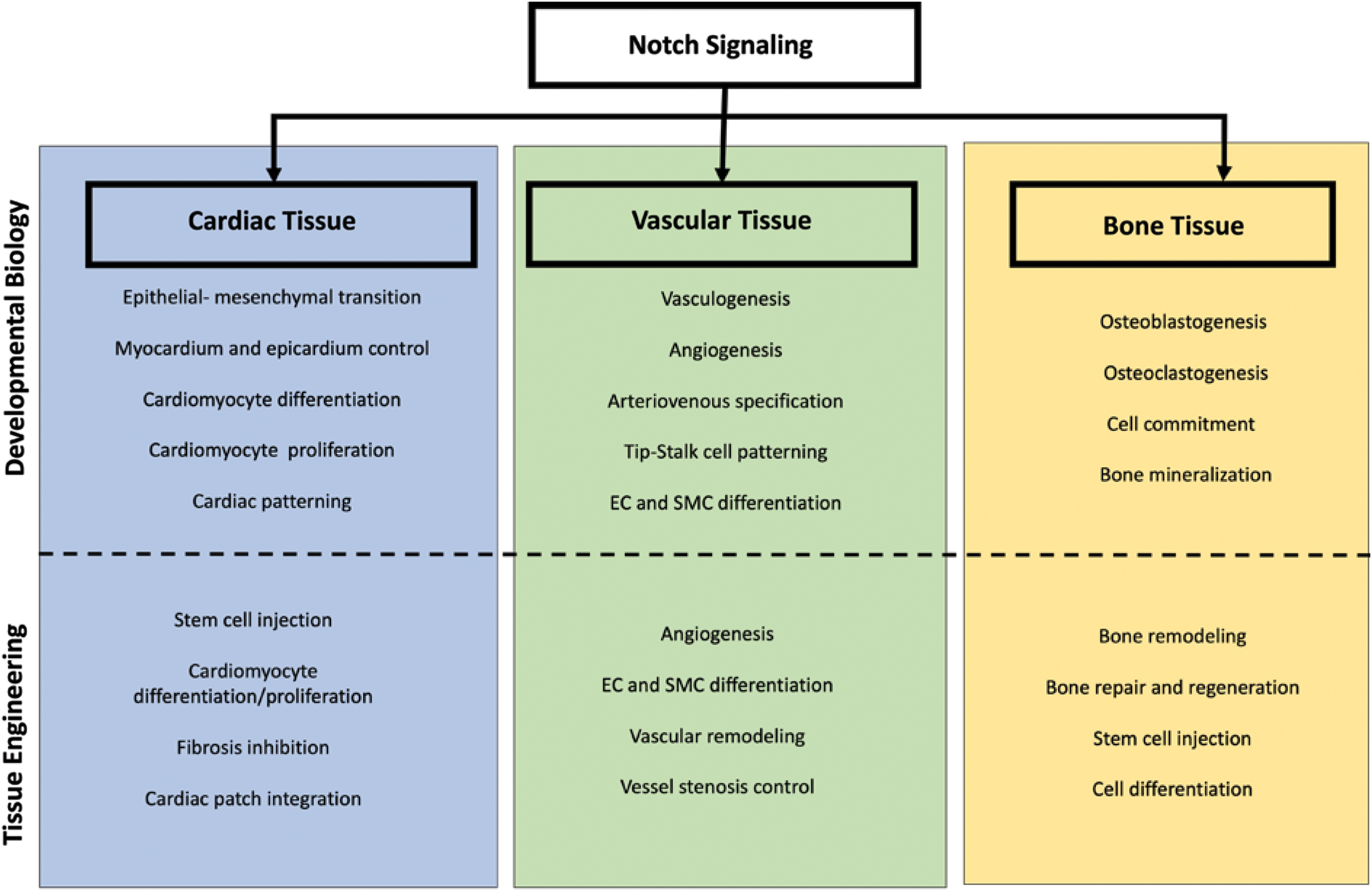
Translation of Notch signaling developmental biology principles to tissue engineering and regenerative medicine. Notch signaling plays a role in tissue development and pathophysiological principles. This signaling pathway is prominent in a wide range of tissues and organs within the body, which include but are not limited to cardiac, vascular, and bone tissue. Here we present selected examples of how Notch signaling in developmental can be translated into a tissue engineering and regenerative medicine approach. Color images are available online.
Notch ligand presentation strategies in tissue engineering
In general, signaling molecules can be presented to cells in one of three ways: (i) adding them as a soluble factor, (ii) direct conjugation or conjugation through a flexible molecular arm to a scaffold, (iii) and affinity immobilization to the scaffold through antibody-binding proteins. As discussed above, adding the soluble form of the ligand does not seem to be effective in activating Notch signaling; hence, attachment of Notch ligands to biomaterial surfaces has been explored and evaluated based on ligand bioactivity on the material surface.170,171 Covalent conjugation chemistries utilize free reactive groups on the surface scaffolds in a random and uncontrolled manner. Numerous covalent conjugation chemistries can be used to immobilize Notch ligands depending on the reactive groups presented on the scaffold surface (Fig. 2B): SH-targeting maleimide chemistry, OH-targeting carbonyldiimidazole chemistry (CDI), and COOH-targeting carbodiimide chemistry (EDC/NHS). If not already present on the biomaterial surface, reactive groups might also be introduced to the surface of the scaffolds through surface modifications like chemical vapor deposition, plasma vapor deposition, and self-assembled monolayers (SAMs), among others. 172 These allow for chemical alterations to the surface of materials without significantly affecting its bulk properties. Covalent immobilization assures ligand presence, but the active site of the ligand is not always accessible for binding due to this random attachment to the surface.
Immobilization through a flexible molecular arm has been applied using polymeric spacers such as polyethylene glycol (PEG),76,173–175 poly(acrylic acid), 176 and disuccinimidyl suberate 175 to allow for better accessibility and activity of the immobilized ligand. Spacer immobilization linked with Notch signaling needs to be investigated further because the interplay between tether length, ligand spacing, and ligand accessibility has not been elucidated. Optimal ligand surface coverage can be maximized with spacers due to the ability of the polymer-bound proteins to form a layer and disperse the ligands in space to optimize binding and minimize lateral repulsions. 177 In fact, the added mobility afforded by longer spacer arms enhanced the formation of focal adhesion due to the ability of cells to reorganize tether peptides on the nanometer length scale. 178 In translation to the Notch signaling system, there is some evidence for target selectivity of specific Notch receptors by which the ability of Notch ICDs to form dimers might influence the activation of downstream targets, including receptor recruiting of gene response. 161 Interestingly, the configuration of CSL binding sites appearing as monomer or dimers has influenced the likelihood of recruiting Notch1 or Notch3 ICD. 179 Therefore, when designing Notch biomaterials, the flexibility of the chemical spacers of varying lengths may enhance the dynamic nature of the ligands. To mimic the dynamic regulation of signaling ligands, polymer chemistry can be harnessed to create chemical spacers to improve biomolecular recognition, ligand accessibility, and increase the dynamic behavior of immobilization. 180
Notch orientation-regulated immobilized scaffolds have been engineered using indirect affinity immobilization strategies. Antibody binding proteins such as Protein A/G, Streptavidin/Biotin binding, as well as anti-Fc antibodies, have been harnessed to control ligand accessibility and orient the ligands with the active site available for binding (Fig. 2C). Compared to covalent immobilization strategies, affinity binding strategies allow indirect immobilization where the active site is oriented controllably for maximized receptor binding capability. Most Notch engineered biomaterials utilize affinity immobilization as it most effectively controls attachment and enhances cellular behavior. Although widely used, affinity immobilization strategies are limited by the binding strength, thus having a higher possibility of dissociating compared to the covalent binding. Protein A and Protein G bind Notch ligands by their biospecific targeting of the Fc domain on recombinant proteins (protein G: kd∼ 10−10, protein A: kd∼ 10−9 M).181,182 Streptavidin/Biotin binding (kd∼ 10−15 M) 183 immobilizes streptavidin to a biomaterial surface through its reactive amine group where the compound affinity binds up to four biotinylated ligands per molecule of streptavidin. Finally, anti-human IgG Fc capture has a strong affinity for the human Fc domain (kd∼ 10−6 M). 182 Using different ligand attachment and presentation strategies, the effect of Notch signaling on cellular function can be studied in both two-dimensional (2D) and three-dimensional (3D) microenvironments to gain insight into various aspects of Notch signaling biomaterial design.
Notch immobilization on 2D biomaterial surfaces
Notch ligands have been immobilized extensively to 2D surfaces to study the behavior of Notch-directed signaling of several cell types, including hematopoietic stem cells, ECs, SMCs, pericytes, MSCs, keratinocytes, and various cell lines (Table 2). With the exception of Dll3, all other Notch ligands have been immobilized to various 2D surfaces. Dll3 is presumed to not activate receptors in trans; therefore, it is not relevant in heterotypic cell signaling. 6 Jag1 is the most commonly immobilized ligand and is used to control a number of cell fate decisions, including angiogenesis, 53 osteogenic differentiation,76,184,185 epithelial differentiation, 186 immunoprotection, 187 blood cell expansion, 188 and cardiac differentiation, 189 demonstrating its various functional roles in multiple tissue types.
Two-Dimensional Immobilization Strategies of Notch Ligands
CDI, carbonyldiimidazole chemistry.
Two-dimensional systems allow us to study culture characteristics, including dose, activation kinetics, and cell behavior, without the complexity of the 3D microenvironment. As discussed in Notch Signaling in Developmental Biology Section Notch signaling is driven by stoichiometric interactions (rather than being driven enzymatically). Therefore, immobilized dose and ligand density are extremely important in activating or restricting signaling between interacting cells. Dose and ligand density can help to modulate Notch signaling in terms of its signaling mechanism through cis- and trans-relationships, based upon stoichiometric ligand–receptor levels. The response to trans-Delta activation is graded, while the response to cis-Delta exhibits a sharp, switch-like response at a fixed threshold. 205 Based on ligand–receptor interactions, there is an ultrasensitive switch between mutually exclusive sending (high Delta/low Notch) and receiving (high Notch/low Delta) signaling states. 205 Overall this alludes to the influence of dose and density of Notch ligands, which can control cell-fate outcomes in different organ systems to regulate cellular outcomes.
Dose and activation kinetics can also control cellular patterning; thus spatial control incorporating patterning will be expanded on in Spatial cues to control Notch ligands section. Immobilization dose dependency has been linked to precoat solution concentration. 189 Within a lymphoid cell fate model dose dependence was explored to evaluate cell fate outcomes of hematopoietic precursors to a self-renewed or differentiated cell fate. Lower Dll1 density enhanced generation of CD34+ cell types consistent with myeloid and lymphoid differentiation, whereas increased amounts of Dll1 induced apoptosis of CD34+ cells. 199 In addition, dose dependence of Jag1 that mediated enhancement of store-operated Ca2+ entry in human pulmonary arterial SMCs was evaluated. 100
The combination of nano/microbead platforms and protein immobilization has led to several applications to enhance nanomedicine. Nanoparticles are often engineered for drug delivery applications, but are limited by their ability to target specific locations and have biodistribution problems. In the application of Notch signaling, nanoparticles with peptide functionalization have been suggested as carriers for targeted delivery of GSIs to block Notch signaling.206,207 In addition, nanoparticles can also be used to deliver immobilized ligands to cell surface receptors and act as cell-surrogate biomaterials replacing the signaling cell to direct Notch signaling. Evidently, bone marrow hematopoietic stem cells were directed to a T cell lineage through Dll4 functionalized microbeads. 200 Furthermore, in human embryonic stem cells (hESCs), Jag1 beads were able to significantly increase Notch signal activation, encouraging ectodermal formation after 2 days, as well as cardiomyocyte differentiation of KDR+ progenitor cells. 189 These, as well as other approaches summarized above, demonstrate the success of Notch ligand immobilization to a biomaterial platform to control cell fate decisions. Translation from nonclinically relevant 2D cell culture models into a clinically translatable 3D biomaterial construct is the next step in developing engineered materials to control cell behavior.
Notch Functionalized Biomaterials for Cell Signaling in a Three Dimensional Microenvironment
PEG, polyethylene glycol; PEGDA, polyethyleneglycoldiacrylate; polyHEMA, Poly(2-hydroxyethyl methacrylate); BMPH, N-(β-maleimido-propionic acid) hydrazide; PBAE, Poly(B-amino ester); PDDA, poly(diallyl dimethylammonium chloride); PLGA, poly(lactic-co-glycolic acid); ECM, extracellular matrix.
Notch ligand immobilization to 3D scaffolds
Immobilization of ligands to 3D biomaterial surface offers an approach to study cellular response, which can better recapitulate the native 3D environments experienced through cell–cell and cell–matrix interactions. Most Notch signaling biomaterials in 3D systems focused on hydrogel systems, although new biomaterials have started to emerge as summarized in Table 3.
Hydrogel based systems are attractive because the mechanical properties of the material can be tuned and controlled to recapitulate the proper cellular niche. Hydrogels are also advantageous due to their biocompatibility and tunable biodegradability. 216 PolyHEMA is perhaps the most commonly used biomaterial as it is resistant to protein adsorption, thus providing a “blank slate” for covalent immobilization studies. 186 PolyHEMA has been used to direct both epithelial 186 and keratinocyte 209 differentiation, demonstrating the versatility of this hydrogel system. Hydrogel systems have been developed to engineer bone tissue. Notch signaling activation during mid-to-late tissue development drove osteoblast differentiation and promoted the anabolic activity of committed osteoblasts. 85 Therefore, from a tissue engineering perspective, activating Notch signaling postinjury could be a potential target for increased bone formation. In addition, Notch signaling biomaterials utilizing Jag1 immobilized to a PBAE hydrogel was used as a novel osteoconductive scaffold for bone tissue engineering. 211
Other 3D materials that have been developed include applications in cardiac tissue engineering, epithelial differentiation, and immunity. For cardiac tissue engineering, an elastomeric poly(acrylic acid) brush grown on a substrate of poly(L-lactide-co-caprolactone) was developed. This elastomer upregulated myogenic but not osteogenic differentiation of human MSCs on the engineered surface, suggesting a new strategy for bioengineering a cardiac patch in myocardial infarction repair. 176 Controlling epithelial differentiation has direct application in tissue-engineered organs where a competent epithelial barrier is required. 186 Promoting squamous cell stratification and rapid differentiation could reduce tissue culture time and improve initial performance. This would also be applicable in percutaneous devices where rapid migration of epithelial cells to the biomaterial surface and cornification/growth arrest upon contact could prevent common modes of failure. An engineered perivascular microenvironment mimicked the endothelial–mesenchymal cell relationship by interacting encapsulated MSCs with surface-immobilized endothelial Jag1. 174
Another application for Notch 3D biomaterials is improving immune response for diabetes and other diseases. Surface modified biomaterials have been used to shift the immune system from an inflammatory to the anti-inflammatory environment by utilizing pegylated Jag1 islet surfaces to direct splenocyte response and improve immunoprotection of pancreatic islets. 187 Collectively, these examples demonstrate attempts at designing 3D biomaterials and strategies to recapitulate the native biology. Although progress has been made, further development in biomaterials is needed. Three dimensional microenvironments allow for incorporating both biochemical and biomechanical signaling that can integrate additional signaling tools within an engineered system. The following section will discuss ways to incorporate complementary cues (rather than just biochemical signals) at the biomaterial interface to better mimic 3D microenvironments.
Controlling Notch Signaling Response at Scaffold Interfaces
Not only is the biochemical signaling activity (ligand orientation and ligand activity) crucial for Notch signaling but also the scaffold must recapitulate the native physical environment (topography, porosity, biocompatibility, etc.). Within the body, cells interact in a highly dynamic environment with signaling cues turning ON/OFF to adapt to various physiological changes in the body. Cells respond best to environments that they are familiar with; therefore, matching the biological microenvironment to tissue engineering materials becomes important for overall cell behavior. Both the physiochemical properties (surface hydrophobicity, electrical charge, morphology, and roughness), as well as structural attributes (pore size in combination with surface modification), can control cell behavior on the surface and enhance the cellular integration and 3D maturation of the engineered constructs.
The physical, temporal, and spatial control of chemical and physical microenvironments thus plays a vital role in development, homeostasis, and even disease states. To better mimic the native tissue, developing strategies to understand the spatial and temporal responses to Notch signaling would be important for the design of engineered tissues. Our understanding of how ligand identity can interact in an interchangeable behavior in some tissues and not in others is limited. As an example, both Dll1 and Dll4 can act to promote T cell differentiation in hematopoietic stem cells but have opposing effects on muscle differentiation in myoblast cells. 21 Understanding the molecular mechanisms relating to dose, dynamics, and timing in both normal and diseased tissue is important to develop proper cellular therapies. The role of topographical biomaterial cues, spatial organization, and dynamic temporal regulation in the delivery of Notch ligands will be further discussed (summarized illustratively in Figure 4A–C).
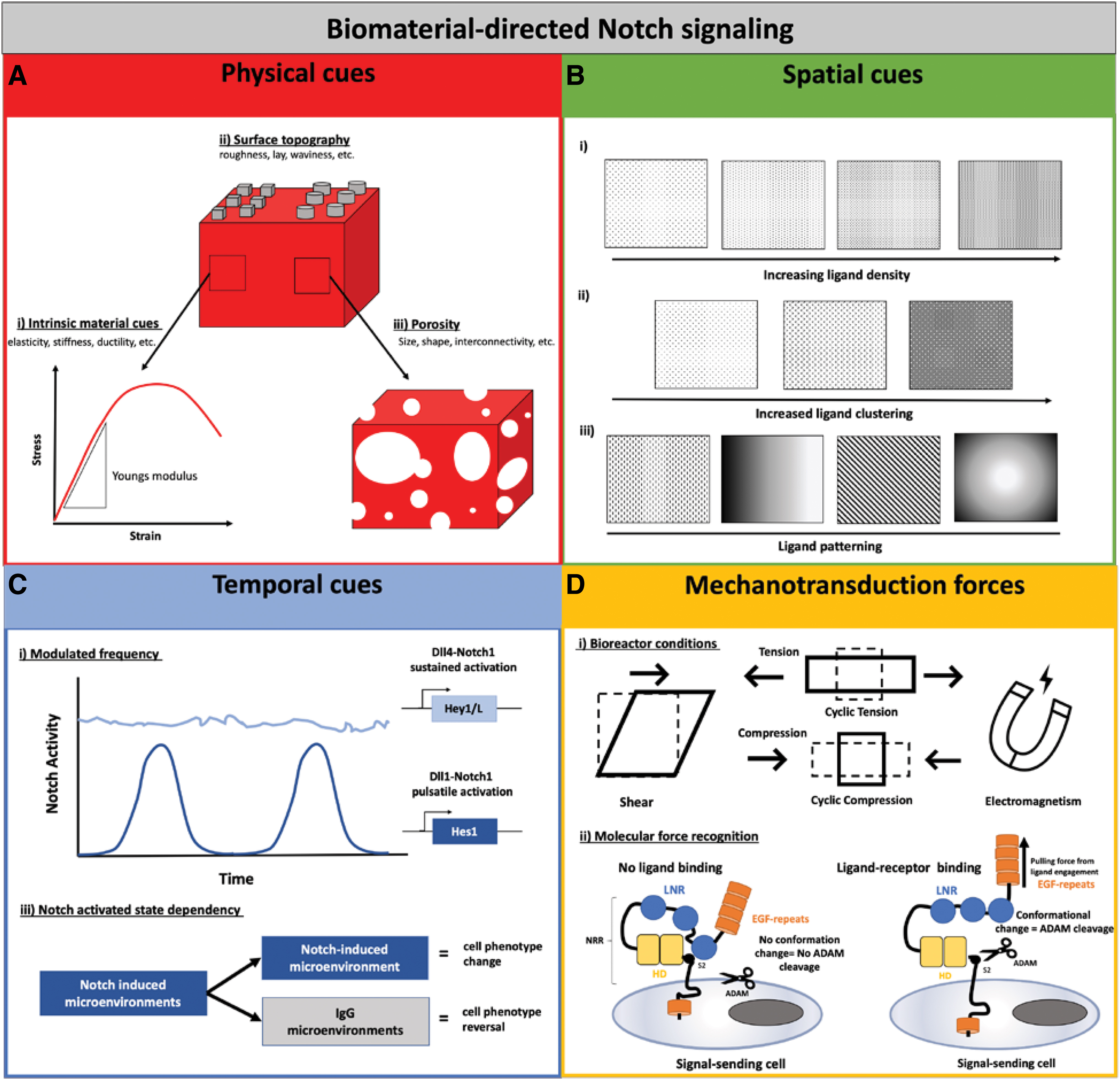
Methods to control and promote Notch-directed cell fate decisions. Cells respond and interact differently in every tissue niche, which is driven by a combination of niche factors. These factors can be incorporated when designing engineered biomaterials to properly replicate each specific niche and direct cellular behavior, including physical, spatial, and temporal cues, as well as mechanotransduction forces.
Physical cues to control Notch signaling: matrix stiffness and topography
Matrix stiffness is a mechanical property that can cause effects on cellular adhesion, migration, proliferation, and differentiation. Cell response has lineage-specific commitments when cultured on substrates matching the stiffness of their corresponding tissues in vivo. Diseased microenvironments are often correlated with increased ECM stiffness and dysregulation of cellular response. 217 Studies have demonstrated that soft substrates are beneficial for a neurogenic or adipogenic differentiation, substrates of intermediate stiffness favor a myogenic lineage commitment, and stiff substrates are beneficial for osteogenic differentiation.218,219 Depending on the cell type seeded on the scaffold, the biomaterial substrate should be designed to properly promote cell behavior so that trans-differentiation into other undesirable cell types is minimized. Scaffold stiffness is, therefore, critical as materials can be designed to have various ranges of modulus, which will control cell behavior differently. 220 Therefore the intrinsic material properties of the scaffold are important for controlling Notch signaling (Fig. 4A-i).
There are only a few studies that investigated the relationship between matrix stiffness and Notch signaling. Within the liver cell niche, both biochemical cues (Notch signaling) and biophysical cues (substrate stiffness) have been linked to the control of liver progenitor cells into both hepatocyte and ciliary cell fates. 221 Progenitor differentiation on stiff (30 kPa) substrates directed peripheral localization of Jag1, Dll1, and Notch2; conversely, there was a lack of expression for Jag1 and Notch2 on a soft (4kPa) substrate. 221 The loss of ligand induction on soft substrates suggests higher responsiveness of Notch signaling within a stiffer substrate in a liver progenitor model. This cited study showed that the segregation of liver progenitor fates is dependent on both Notch signaling and substrate stiffness. 221 In addition, hydrogel stiffness can activate Notch1 signaling and regulate the cardiac progenitor cell gene expression and function in vivo. 212 An increased hydrogel concentration in the presence of the Notch ligand Jag1, in turn, amplified Notch1 activation, indicating that stiffness can affect Notch1 activation in 3D. 212 Furthermore, higher concentration hydrogels increased Hes1 expression and its downstream target Jag1, which is a ligand that could sustain Notch1 activation. Interestingly culture on lower stiffness hydrogels resulted in increased endothelial and smooth muscle gene expression, controlled through Notch1 activation. 212 This suggests that substrate stiffness can control Notch signaling cell fate decisions.
In addition, the effect of substrate stiffness has been implied within the MSC niche. A biomaterial system has mimicked this niche by tailoring PEG hydrogels to best promote Notch directed MSC behavior. In combination with Notch ligand immobilization, the hydrogel properties were also tailored to various stiffnesses. Material stiffness triggered the formation of microcapillaries in soft PEG matrices (1–1.3% PEG with 0.074 − 0.276 kPa), but as the hydrogel stiffness increased (3% PEG, 2.157 kPa), networks were almost completely absent, 174 indicating that hydrogel stiffness plays a role in promoting Notch directed cell behavior. Reciprocal interactions of cells within the biomaterial led to a dynamic reshaping of the initial microenvironment, which further allowed development and ECM deposition. Finally, material surface coatings, including layer-by-layer deposition, have been used to tune the stiffness and promote cell contacts. A clay/PDDA film was used to provide a stiff/rigid layer on the hydrogel matrix, followed by a DLL1 layer. 213
In addition to matrix stiffness, various topographical cues created through pore size manipulation, surface treatments, and various scaffold fabrication methods (electrospinning, bioprinting, freeze-drying, solvent leaching, etc.) also influence Notch signaling (Fig. 4A-ii, iii). This is important because surface roughness and patterning can affect the way that cells adhere and orient to the surface. A polycaprolactone/hydroxyapatite (PCL/HA) material compared to PCL alone displayed increased surface roughness, which promoted osteogenic differentiation in periodontal defects directed by immobilized Jag1. 204 Although PCL/HA membranes displayed lower mechanical properties, the surface roughness improved cell attachment and spreading, the nanostructured/hydrophobic surface also increased osteogenic differentiation compared to PCL surfaces alone. To create the thymic stroma, a 3D inverted colloidal crystal (ICC) scaffold coated with DLL1 was fabricated. 213 The ICC topography was tested in a rotary dynamic cell culture system, where convective media flow was the major driving force for cellular motility within the scaffold. It was also determined that the ICC combined with the layer-by-layer surface modification promoted cell adhesion.
To explore topography further, SAMs and supramolecular motifs have been investigated. Surface-modified gold surfaces utilizing SAMs have been explored for Jag1 immobilization, where they were used to create patterns on the surface that encouraged cell adhesion and orientation. 188 In addition, supramolecular UPy-poly(ɛ-caprolactone) (UPy-PCL) modified with a UPy-conjugated Tz (UPy-Tz) additive was also studied using drop cast and electrospun films. 170 Primary human VSMCs endogenously expressing the Notch3 receptor were cultured on UPy-PCL/UPy-Tz electrospun meshes modified with TCO-pG/Fc-Jag1 and found upregulated the expression of Hey1, Hes1, Jag, and Notch3 compared to drop cast films. The larger surface area and potential for higher ligand concentration of the surface provided by electrospun mats were superior to the drop cast films and were able to affect cell adhesion, viability, and Notch functionality. 170
Topographical and intrinsic material cues could be important in Notch signaling because of a recent report that cell–cell contact area can affect Notch signaling and Notch-dependent patterning. 222 Cell fate decisions where larger cell–cell contact communication resulted in a higher expression signal and opposingly smaller cell–cell contact created less signal, which is highly dependent on scaffolding conditions. 222 The various biomaterial surfaces also created a better environment for ligand coating efficiency by forming higher electrostatic charges. 213 Stiffness could also play a factor in Notch signaling cell fate decisions based on the hypothesis that a ligand-mediated pulling force must be exerted on the NECD for Notch activation. 13 Thus, controlling substrate stiffness can control the traction forces and, therefore, Notch activation.
Spatial cues to control Notch ligands
Spatial patterning allows researchers to explore the effects of clustering and density on Notch activation 208 and also precise control of stem cell fate. Biologically, clustering and ligand density are driven dynamically within the cell membrane, and it is known that ligands form clusters and dimers, which are needed as a structural requirement for ligand stabilization.190–192 Extending this to engineered systems, the notion of clustering and oligomerization using spatial patterning is suggested in controlling cell fate decisions.171,203,208,209 Spatial patterning and gradients of various ligands and soluble factors extend past this and are highly important in the development of complex multicellular organisms to emulate tissues. Cellular patterning is important in biological tissues, including retinal patterning and sprouting angiogenesis. The patterning complexity of various ligands, ECM molecules, growth factors, and other factors to control and regulate the cellular niche is important when designing engineered tissue. Multimolecular patterning has thus started to emerge in the biomaterials field to direct cellular behavior and has been done with a variety of molecules, including cell-adhesive RGD peptide, 223 fibronectin, 224 albumin, 225 and others as summarized previously. 226
Up to this point, we discussed the activation of Notch signaling using engineered biomaterials, where immobilization of ligands was homogeneously coated on a biomaterial surface. Since Notch signaling is contact dependent and operates through both lateral activation and inhibition, the patterning of specific ligands influencing the sender/receiver state of cells could be a beneficial technique to control cell fate decisions. Therefore, spatial patterning could include the use of ligand density, ligand clustering, or specific patterning to control Notch-directed cell fate decisions (illustrated in Fig. 4Bi–iii). Patterning of biomaterial surfaces can be integrated during both fabrication (i.e., stereolithography) or postprocessing of a uniform biomaterial using techniques, including microcontact printing. 226
In the vasculature, as discussed previously, Notch ligands Dll4 and Jag1 have opposing effects on angiogenesis. 53 VEGF signaling induces Dll4 in tip cells; tip cells then suppress tip–cell features in adjacent stalk cells through Dll4/Notch-mediated lateral inhibition. 34 Conversely, Jag1 antagonizes Dll4-mediated Notch activation in stalk cells to increase tip–cell number and enhances vessel sprouting. 44 It is this Jag1 and Dll4 salt and pepper pattern that dictates the phenotypes of cells within this niche into tip–stalk phenotypes. Using microcontact printing, patterning of Dll4 ligand immobilized to pink fluorescence beads in parallel lines controlled angiogenic sprouting. 201 Regulation of the location and direction of endothelial sprouting was achieved using micropatterns to locally inhibit angiogenic sprouting and direct the formation and location of new sprouts using the Dll4 negative regulator.
Similarly, microarray patterning in circular patterns of ECM copresented with Notch ligands provided a biochemically and biophysically defined microenvironment for liver progenitor differentiation. Patterning of Notch ligand Dll1, Dll4, and Jag1 showed that cells were spatially localized based on the patterning with segregated differentiation of progenitors toward biliary fates peripherally and hepatocytic fates centrally. 221 Spatial presentation of ligands on biomaterial surfaces consequently has promise in manipulating and controlling cell fate decisions for more controlled regenerative medicine applications. Future directions include exploring various other ligand–receptor patterns, as well as various combinations of ligands.
Temporal cues to control Notch signaling
To further expand the capabilities of Notch signaling biomaterials, temporal control of ligand delivery should be evaluated. Delivery of signals can be controlled temporally by modulated frequency, dose dependence, duration of cultures in a Notch activated state, as well as temporal control of cell life cycle. Temporal frequency in a pulsatile or sustained delivery could affect the cell fate of myoblast cells. 21 Single-cell imaging revealed that Dll1 activated Notch1 in discrete frequency-modulated pulses that specifically upregulate Notch target gene Hes1, promoting myogenesis. In contrast, Dll4 had sustained activation dynamics activating target gene Hey1, inhibiting myogenesis. It has also been indicated that Hes1 activation is relatively insensible to the duration of Notch activation and could be induced strongly by brief pulses or by sustained activation; however, the activation of Hey1 and HeyL was more sensitive to the duration and accumulation as long as Notch activation was maintained (illustrated in Fig. 4C-i). This indicates that temporal delivery of Notch ligands in cellular therapies could be used to selectively activate Hes1 versus Hey1/L activation, which would result in different cell fate determinations. Interestingly increased dose of Dll1 increased the pulse frequency, whereas an increased Dll4 dose caused an increased amplitude by still a steady-state response. Dll1 signaling was deemed as frequency modulated, and Dll4 was amplitude modulated. Although interesting to consider when evaluating the effectiveness of dynamic Notch control using engineered biomaterials, it is still unclear how this translates to other Notch ligand–receptor relationships.
Not only do various tissues respond differently to the ligand identity but additionally the stimulation of cells within a niche at different times in their cellular specification can lead to different cell fates. Biphasic effects of Notch activation have been demonstrated in cardiac differentiation. Jag1 stimulation and early activation of undifferentiated hESCs promoted ectodermal differentiation and, in contrast, Notch activation in specified cardiovascular progenitor cells increased cardiac differentiation in these cells. 189 Within the pancreatic field of pluripotent derived cell niche, the duration of time cells spent in a Notch active state was also indicated to affect lineage specification. 227 Together, this may indicate a link between temporal delivery and dynamics of Notch signaling in cell fate lineage specification.
From a biomaterial perspective, Jag1 modified 3D microenvironments have induced target genes in MSCs that were found upregulated in cocultures with ECs, indicating that biofunctional biomaterials can be used to reprogram cells in the absence of ECs. There is some indication that biomaterial driven control of Notch signaling can induce a Jag1 mediated ECM switch in MSCs that was reversible when exposed to control IgG-modified microenvironments. Temporal delivery of Jag1 for 7 days followed by a subsequent 7-day exposure to igG microenvironments reverted the BM-MSC ECM from an induced perivascular phenotype back toward a vascular basement membrane phenotype 174 (illustrated in Fig. 4C-ii). This indicates that Notch signaling biomaterials can be involved in reversibly reprogramming cellular environments, and signaling dynamics are involved in various cell fate switches in the body. When the ligand–receptor dynamics is controlled, this results in proper homeostasis; however, when dysregulated ligand–receptor dynamics can cause diseased states. Temporal delivery of ligands can thus be incorporated into the design of Notch biomaterial cellular therapies. Further research should thus be done to investigate the behavior and controlled activation of Notch signaling utilizing dynamic encoding with biomaterials.
Incorporation of Mechanotransduction Forces to Direct Notch Signaling
Multicellular tissues are subject to various external forces, including compressive, tensile, and fluid shear stresses, which help maintain tissue integrity and homeostasis. Mechanotransduction is the mechanism by which cells translate external forces into a biochemical signal. It is thought that there is a common interplay between biochemical and biomechanical patterning in the body, which leads to proper homeostasis of cell-specific niches. 228 Mechanotransduction also occurs at a molecular level whereby mechanosensing involves force modulated interaction and conformation changes in dynamic ligand–receptor interactions. 229 Thus mechanotransduction forces are a potentially useful element to integrate into tissue-engineered systems, both through incorporating various bioreactor conditions for tissue maturation/stimulation, as well as replicating molecular forces native to Notch signaling (summarized in Fig. 4D).
Cellular mechanosensing by bioreactor conditions
Notch signaling has been shown to regulate its behavior by a mechanical stimulus in the cellular microenvironment. Various bioreactor conditions can be controlled and maintained to mature and direct cellular response in engineered tissue (Fig. 4D-i). Mechanical loading of bone primarily through strain forces is sensed by bone cells resulting in biochemical signals that result in the activation of mechanosensors. In vitro, Notch signaling has been activated by mechanical strain loading through cyclic stretching, 230 intermittent compressive stresses,231,232 and pulsed electromagnetic fields. 233 This indicates a functional link between Notch activation and mechanotransduction caused by mechanical or electromagnetic loading, which could be utilized for in vitro Notch activation.
In addition, within the vasculature Jag1 and Notch3 target genes were downregulated with strain magnitude defined by the vascular geometry and blood pressure upon increased or decreased wall thickening caused by vascular morphogenesis. 46 Notch signaling has also been shown to be modulated through shear stress caused by blood flow through the body due to cycles of contraction and relaxation of heart tissue. Endothelial Notch1 is responsive to shear stresses and is necessary for the maintenance of junctional integrity, cell elongation, and suppression of proliferation phenotypes induced by laminar shear stress. 234 It has been shown that both angiogenesis and osteogenesis are defective when blood flow is impaired in vessels, which is coincident with downregulated Notch signaling in ECs. 230 Notch signaling also controls the expression of fluid flow responsive genes in ECs and modulates the formation of fluid flow sensing primary cilia. 234 Mechanotransduction forces help maintain homeostasis of various tissues as discussed; however similar to how ligand–receptor dysregulation can occur, dysregulation can also be driven by mechanotransduction forces. Evidently, mechanotransduction and Notch signaling have been linked in tumor progression and disease. 228 Together these findings substantiate that Notch signaling is influenced by its mechanical environment and is a mechanosensing signaling pathway. Incorporating mechanical signals to cell-seeded scaffolds using bioreactors capable of exerting compressive or shear forces may accelerate our understanding of Notch dynamics in an in vitro system.
Molecular level mechanosensing
There is also evidence suggesting that Notch signaling driven by cell–cell interactions are mechanosensitive at the molecular level (Fig. 4D-ii). Crystal structures have revealed the overall Notch receptor–ligand conformation indicating that the S2 binding site is deeply embedded within the Notch heterodimer Lin12-Notch repeats domain and, thus, is protected from metalloprotease cleavage and creates an autoinhibited conformation.152,235 The Notch pulling model indicates that the Notch regulatory region of the receptor acts as a force sensor that is unfolded by a threshold level of mechanical tension generated across the ligand/receptor bridge. This tensional force in a cell–cell signaling system is proposed to be caused by the endocytosis force at the ligand–receptor complex 131 ; however, this is highly debated. 236 Nonetheless, in the absence of endocytosis, Notch ligands were shown to accumulate on the cell surface and failed to activate Notch signaling on neighboring cells. 10
Various molecular force sensors have been developed to study cellular forces acting through single mechanosensitive receptors. Force quantification techniques for cell biology include 2D traction microscopy, micropillars, cantilevers, atomic force microscopy, fluorescence resonance energy transfer, optical tweezers, magnetic tweezers, and tension gauge tethers (TGTs). 237 TGTs are force sensors which have been widely used to study Notch signaling. Protein G based TGTs were used to study the force magnitude needed for signal activation. Recombinant ligands with IgG-Fc fusion are assembled using DNA tethers with different tension tolerances immobilized through a glass surface passivated with PEG. Notch ligand Dll1 was tethered to the surface, and using a reporter cell line, it was determined that under 12pN of force was required for signal activation.238,239 Force-induced Notch activation has also been studied at the ligand–receptor bridge using Notch ligand immobilized to magnetic biomaterial nanoparticles. Magnets have been utilized using a magnetic tweezer assay to apply a range of pN-scale forces to the Notch receptor on the cell surface, and Dll4-loaded magnetic beads induced Notch signal activation with the addition of this mechanical tension force. 240 This further suggests that force must be applied to bead-tethered ligands to further induce the canonical proteolytic steps responsible for Notch activation. It also indicates that the Notch signaling system is controlled through the interplay of both biochemical and biomechanical signals.
The role of mechanotransduction within the Notch signaling pathway is still controversial and a highly debated topic of research currently. Considering these concepts, the interplay between mechanical force transduction and its interaction with ligand immobilized to the surface of biomaterials poses another important challenge in the integration of these biomaterials into complex cellular processes, including the mechanical conditions and molecular mechanotransduction. Mechanotransduction within Notch signaling thus far has been very context dependent; therefore, various ligand–receptor combinations could prove to be more mechanosensitive than others. In addition, the delivery of chemical signals through a biomaterial surface presents the absence of the mechanical tension force found biologically through the pulling force caused by cell endocytosis of the ligand–receptor complex. Therefore, methods must be investigated in how to provide accurate traction and tension forces within biomaterial-based Notch signaling systems to allow for Notch activation to occur. The application of mechanotransduction to the activation of Notch presenting biomaterials could be a possible target to enhance signaling and repair of various processes.
Conclusions and Future Perspectives
Notch signaling has been identified as a promising target for a vast number of cellular therapies and shows considerable progress and potential in directing and reprogramming cellular behavior. Although considerable progress has already been made in the development of Notch signaling biomaterials, there is more yet to be known. Designing signal-presenting biomaterials for directed cellular therapies is a complex process that requires precise control of signals. While many of these signals come from the ligand of interest, such as ligand orientation, spatial patterning, and temporal delivery, others rely on the properties of the scaffold or the external microenvironment. The interplay between these elements is what creates such complexity in engineering Notch targeted cellular therapies. An ongoing challenge is the integration of the mechanosensitive nature of Notch signaling into the biomaterial system. Many researchers have largely focused on the cellular response driven through a biochemical perspective, but a major limitation in engineered tissues is driven from a biomaterial perspective. Designing cell-instructive biomaterials incorporating the biological activity of proteins is an emerging field. These biomaterials could be applied to the Notch signaling system to create a more dynamic microenvironment and introduce tissue mechanics and activate mechanosensitive receptors with various forces introduced.
Depending on the context, activating or suppressing Notch signaling is a valuable tool to engineer tissues for therapeutic or diagnostic use. Notch signaling can be utilized to engineer prevascularized thick tissues or to develop antiangiogenic cancer therapeutic strategies. This, in turn, allows creating diseased model tissues for studying drug discovery and screening. It has already been demonstrated as a potential strategy to prevent atherosclerosis, 241 promote cardiac valve regeneration, 242 reduce cell senescence in cell sheet engineering, 243 and to engineer vascular tissues.27,28 Since endocytosis of the ligand following receptor binding generates the force to render Notch S2 cleavage, it implies that attaching the ligand to nondynamic surfaces may not be sufficient to activate Notch. Two strategies to address this could be by use of supramolecular biomaterials/scaffolds that having dynamic, interchangeable, and reversible motifs170,244 or by reverse engineering of the Notch receptor for reduced pulling force dependency. 245 Since the latter strategy involves transfection for synthetic Notch expression, supramolecular materials may be an attractive avenue from a tissue engineering perspective. Regardless of the strategy to activate Notch signaling, it remains to be an excellent tool for tissue engineering and regenerative medicine.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Heart and Stroke Foundation of Canada (Grant # G18-0022193) and the Natural Sciences and Engineering Research Council of Canada (Grant # RGPIN2018-06310).