
Abstract
Peripheral nerve injury (PNI), a challenging neurosurgery issue, often leads to partial or complete loss of neuronal functions and even neuropathic pain. Thus far, the gold standard for treating peripheral nerve deficit remains autografts. While numerous reviews have explored PNI and regeneration, this work distinctively synthesizes recent advancements in tissue engineering—particularly four-dimensional (4D) bioprinting and exosome therapies—with an emphasis on their clinical translation. By consolidating findings spanning molecular mechanisms to therapeutic applications, this review proposes an actionable framework for advancing experimental strategies toward clinically viable solutions. Our work critically evaluates emerging innovations such as dynamically adaptive 4D-printed nerve conduits and exosome-based therapies, underscoring their potential to match conventional autografts in achieving functional restoration.
Impact Statement
Although several previous reviews have been made on describing with great detail the degenerative and regenerative mechanisms of the peripheral nervous systems, as well as the several existing and exploratory treatment strategies, we focus more on the latest advancements of each of those topics.
Introduction
The nervous system in the human body comprises the central nervous system (CNS) and the peripheral nervous system (PNS). 1 Neuronal damage initiates an autonomous regenerative process. The bands of Büngner, composed of Schwann cells (SCs) within tubes of the basal lamina, are crucial for subsequent axonal regrowth. 2 Evidence suggests that PNS has an intrinsic ability to regenerate more effectively than the CNS, as it can rapidly clean debris and form bands of Büngner instead of a glial scar.3,4 Peripheral nerve injury (PNI), primarily caused by acute mechanical trauma 5 such as traffic accidents and high-risk sports, is a critical social problem that can result in permanent motor and sensory deficits. Currently, new cases of PNI in developing countries range from 13 to 23 per 100,000 people per year, which shows a gradually increasing trend, but the effectiveness of surgical treatment for PNI varies between 36% and 51.6%, which is influenced by the site and severity of the injuries. 6
The process of peripheral nerve damage and repair is highly complex. Significant progress has been made in understanding its potential mechanisms. 7 Crucially, one of the primary prerequisites of the successful outcome of peripheral nerve regeneration (PNR) is the sustained survival of neurons, 5 as their condition is essential for receiving the retrograde signal and activating SCs. Additionally, factors such as the injury site, timing of repair, and patient age significantly influence successful neuronal regeneration. 8
While numerous reviews have explored the mechanisms and treatments of PNI, this review distinguishes itself by focusing on the latest advancements in tissue engineering and regenerative medicine, particularly the integration of four-dimensional (4D) bioprinting and exosome-based therapies. We aim to provide a comprehensive analysis of how these emerging technologies can address the limitations of current treatments, such as the static nature of traditional nerve conduits and the challenges of achieving complete functional recovery. By synthesizing recent findings from molecular mechanisms to clinical applications, this review offers new insights into the potential of dynamic, biomimetic approaches to nerve regeneration. Furthermore, we highlight the translational potential of these technologies, providing a roadmap for future research and clinical implementation.
Neurobiology of PNI and Regeneration
Response to nerve injury
Upon peripheral nerve cut, the distal nerve segments undergo Wallerian degeneration, an important response to trauma. 7 Neurons experience axonal degeneration after being physically separated from their cell bodies due to nerve injury. Augustus Waller first observed and described axonal degeneration after axotomy in 1850. 9 Specifically, the soma of neurons in the proximal nerve segment undergoes chromatolysis featuring swelling and Nissl granule disintegration within 48 h. 10
Axon degeneration is caused by calcium infiltration through ion-specific channels, which subsequently stimulates axonal proteases; the disintegration and degeneration of the axolemma and axoplasm occur within 24 h in small nerve fibers and 48 h in large nerve fibers. 11 After injury, mature neurons use axonal mRNA translation to transmit injured signals toward the nucleus of the neuron. Following PNI, the de novo synthesis of importin-β and vimentin contributes to an importin-activated Erk-vimentin complex, which recruits retrograde motor dynein to link the nuclear import mechanism with retrograde injury signaling.12,13 The initial breakdown of myelin postinjury relies on the activation of phospholipase-A2 (PLA2) enzymes. 14 Both cytosolic and secreted PLA2 forms hydrolyze phosphatidylcholine, a phospholipid, into lysophosphatidylcholine (which triggers myelin disintegration) and arachidonic acid (which can be converted into proinflammatory eicosanoids). 15 Notably, cytosolic and secreted PLA2 forms are increased in SCs and macrophages a few hours after injury and remain elevated for up to 2 weeks. 16 This timeline aligns with the progression of Wallerian degeneration after PNI, and the return to baseline PLA2 levels 3 weeks post-PNI coincides with axon regeneration and remyelination. De et al. also discovered that inhibiting PLA2 expression/activity in the sciatic nerve distal to the injury site significantly prolonged the clearance of degenerating myelin and axons. 16 Thus, PLA2 activity is essential for initiating myelin breakdown and advancing Wallerian degeneration following PNI. SCs then detach from the axons and transdifferentiate into repair cells under the influence of transcription factors such as c-Jun. 4 Loss of c-Jun results in delayed repair and functional deficits. These repair cells are unable to establish the myelin sheath but contribute to inflammatory debris removal via autophagy within the first 5 days after PNI. 17 Previous studies have elucidated the mechanisms by which several immune cells initially detect injury to adjacent axons. Toll-like receptors (TLRs), initially characterized for their role in identifying microbial pathogens and initiating intracellular inflammatory responses, 18 have been found to recognize tissue injury through the combination with endogenous ligands that do not typically exist in the extracellular environment such as heat shock proteins, mRNA, and degraded extracellular matrix (ECM) components.19,20 SCs express a range of TLRs, with TLR3, TLR4, and TLR7 being constitutively expressed in unstimulated cells, and TLR1 being elevated following transection. 21 The expression profile of TLRs demonstrates that SCs serve as sentinels in the PNS. In fact, PNI triggers TLR-dependent alterations in the activation of transcription factors and the process of Wallerian degeneration, as well as functional recovery. Notably, resident macrophages are triggered to enable circulating macrophages to rapidly infiltrate into damaged neuro-microenvironment. 22 Over time, they undergo phenotypic changes. 23 During the early phase posttrauma, circulating macrophages at the damage site express and include both M1 and M2 phenotypes, with a predominance of M1. 24 They then polarize to an M2 phenotype, which is the primary phenotype to enhance regrowth. 25 During the period from days 2 to 7 following injury, circulating macrophages experience substantial proliferation, peaking between 1 and 2 weeks. 26 Importantly, macrophages can continue existence at the damaged nerve sites for at least 42 days, 27 contributing to debris removal and subsequent regrowth posttrauma. Figure 1 illustrates the process of Wallerian degeneration.
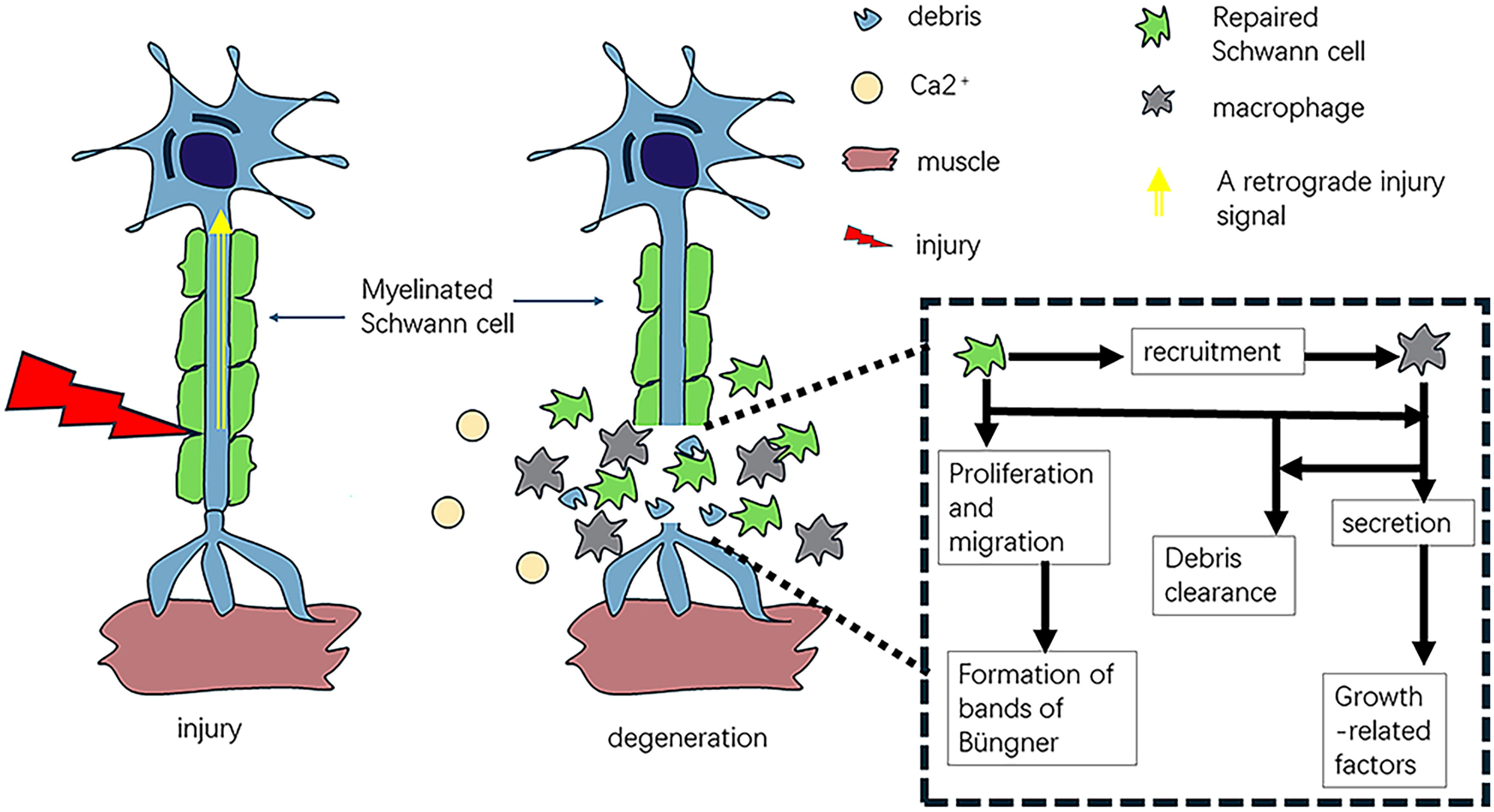
Wallerian degeneration. Upon peripheral nerve lesions, a retrograde injury signal is transferred to neuronal soma; as a result, Schwann cells proliferate, dedifferentiate, and recruit macrophages. These two cells then exert the phagocytosis functions to contribute to debris removal, a prerequisite that is critical for axonal outgrowth.
Nerve regeneration
Axonal regeneration primarily comprises several key steps. Initially, genetic programs of axon regrowth are triggered within damage nerves. 28 Myelin protein levels are downregulated in injured neurons, while the secretion of adhesion molecules, growth factors (GFs), and their receptors such as neuregulin, glial-derived neurotrophic factor (GDNF), insulin-like growth factor 1 (IGF-1), transforming growth factor-β (TGF-β), neurotrophins, and fibroblast growth factor 2 is promoted. 4 Additionally, dorsal root ganglion (DRG) sensory neurons upregulate regeneration-associated genes (RAGs) while downregulating nerve identity-related genes. Peripheral sensory nerve transection leads to increased acetylated histone H3 and nuclear P300/CBP-associated factor (PCAF), promoting various RAGs. 29 Beyond neurons, extrinsic stimulation that primarily includes SCs and macrophages is essential to enhance regeneration.30,31 After an injury, SCs reprogram to adopt reparative phenotypes, releasing various neuroregenerative factors to transmit positive signals toward damaged nerves. 32 Recruited macrophages, activated by SCs, create a conducive milieu to support nerve regeneration by balancing pro- and anti-inflammatory factors and removing debris. 33 Because of these, millions of axons sprout and extend toward the distal targets. 31 The further step indicates the directed extension of neurites, which is induced by SCs. 34 Repaired SCs collect to form cell cords and move to guide axons across the damaged sites. 35 Macrophages assist SC migration. Posttransection, a hypoxic nerve bridge forms between the distal and proximal nerve segments, 36 which can be detected by macrophages within the bridge, resulting in activating transcription factor hypoxia inducible factor-1α (HIF-1α) to enhance vascular endothelial GF (VEGF)-A secretion. VEGFA, in turn, promotes a polarized vasculature within the bridge, which can serve as a substrate for SC movement. The final step includes the reinnervation. SCs induce motor or sensory neurites to extend to form many branches by generating various neuroregenerative molecules and chemokines. 37 In addition, nonmyelinated postsynaptic SCs located in the neuromuscular junction endplate area guide axon buds to reinnervate adjacent muscle fibers that have lost nerve supply by extending their protrusions. Actually, axon elongation is directed toward the target tissue, forming the synaptic partly by the growth cone, a motile and actin-based architecture at the distal tip of axons, 38 guided by multiple attractive and repulsive signals. 39 In the region distal to the injury site, regenerating axons face numerous challenges before effectively reinnervating the target organ. Axons that are misdirected toward incorrect targets can lead to suboptimal functional outcomes, even if a substantial number of axons regenerate. 40 However, this issue is somewhat mitigated by a process known as “pruning,” which eliminates growth cones that fail to reach the appropriate target or lose the support of their endoneurial tubes. 41 The absence of neuronal contact in the distal stump results in chronically denervated SCs. These SCs downregulate GFs and enter a dormant state, rendering them incapable of supporting axon extension. 42 Furthermore, the denervated distal tissue experiences a depletion of trophic factors and muscle fibers undergo atrophy. 5 These detrimental effects significantly hinder functional restoration posttrauma.
Current Therapeutic Strategies for PNI
Surgical treatment
Surgery treatments for peripheral nerve defects focus on mitigating mechanical pain and restoring motor functions. Several repair strategies are tailored to different types and sites of nerve injuries. Over the past decades, surgical approaches to neuronal repair postinjury have primarily included nerve autografts, nerve allografts, and nerve transfers. The gold standard of large-gap nerve injury, especially brachial plexus injuries, is nerve autograft, which leads to better regenerative outcomes. Nerve allografts and nerve transfers are also traditional surgeries for repairing nerve damage and have been extensively reviewed. 43 However, these neurosurgical strategies have several disadvantages that limit their application, necessitating the search for alternative approaches to neuronal repair. Recently, the concept of a nerve regeneration chamber has been proposed, which creates a permissive milieu for nerve regeneration by hampering fibroblast accumulation. 44 This emerging technology addresses some of the limitations of traditional surgery, such as neuroma formation and limited donors. In the following section, we will elucidate the latest advancements in tissue engineering to enhance peripheral nerve regrowth.
Nerve-guided conduits
Nerve-guided conduits (NGCs), tissue-engineered tubular architectures composed of natural and/or artificial biopolymers, can bridge large gaps in neuronal damage and have attained extensive attention in recent years. 45 NGCs assist in PNR by closely mimicking the natural nerve-regenerating milieu. Furthermore, they can also be loaded with neurotrophic factors, collages, and mesenchymal stem cells (MSCs). However, they must be highly biocompatible, biodegradable, elastic, and mechanically stable in order to achieve better regenerative outcomes. 46
Chitosan has been identified as an effective material for decorating biological scaffolds to promote PNR in recent years. Previous studies have demonstrated that porous micropatterning containing chitosan that was developed by using the technology of micromodeling and lyophilization showed promising capacities in coordinating SC functions, ultimately enhancing successful neuronal regeneration. 47 Hydrogel has also been used for PNI treatment, especially when incorporated into nanofiber. Zheng et al. 48 conducted an in vivo experiment where aligned poly (l-lactic acid) nanofibers and 0.25% porcine decellularized nerve matrix hydrogel combined to form NGCs, which were applied to repair 5 mm sciatic nerve transections in rats. Walking track analysis indicated that the sciatic functional index (SFI) of the NGC-treated group significantly improved compared with the control group after implantation, suggesting that this hydrogel NGC has a beneficial impact on functional recovery after peripheral nerve transection. Furthermore, histological assessments and immunostaining showed significant enhancement in axonal elongation, SC migration, and myelin sheath reestablishment. Taken together, these findings suggest the robust therapeutic potential of NGCs consisting of nanofibers and hydrogel in promoting neuronal regrowth. 48 Yang et al. 45 constructed an aligned fibrin/functionalized self-assembling peptide (AFG/fSAP) interpenetrating nanofiber hydrogel. In this study, the AFG/fSAP hydrogel was used to repair a 15 mm transection injury of the sciatic nerve in rats, resulting in an increased number of myelinated nerve fibers and improved motor functional recovery. In vitro experiments indicated that this hydrogel could induce SC collective migration and promote nerve growth-related factor secretion, 45 suggesting its effectiveness in PNI repair. Moreover, Soury et al. reported that the implantation of the fibrin-collagen hydrogel NGC seeded with adipose-derived MSCs (ADSC) in a 15 mm-sized sciatic nerve defect upregulated nerve growth-related factors such as neuregulin 1 gene, c-Jun, and activating transcription factor 3 compared with hollow chitosan NGC treatment, 46 resulting in better outcomes of PNR. Taken together, these data illustrate that NGC decorated with hydrogel serves as a favorable strategy to encourage neuronal regrowth and synapse formation.
The structure and mechanical compatibility of NGCs
The mechanical attributes of NGCs are crucial for bridging nerve gaps. One of the key mechanical functions of an NGC is to form a protective barrier that shields nascent axons from invading scar tissue, prevents surrounding tissues from squeezing the regenerating tissue, and offers stable structural support until the nerve has regenerated sufficiently. 49 Moreover, the NGC must be able to withstand tearing caused by sutures if they are employed. 50 Given that peripheral nerves are subjected to stretching, compression, and shearing forces, the NGC must also be flexible and capable of withstanding these mechanical stresses. 51 To produce NGCs with appropriate strength and flexibility, it is necessary to estimate the mechanical properties of the target area, and therefore, the choose of materials should be taken into consideration to ensure they match the unique mechanical characteristics of the target tissue. 52 Size mismatching is a primary limitation while utilizing autografts. Among various tissue engineering materials, poly(e-caprolactone (PCL)-based polymers have mechanical properties comparable to a natural-nerve nerve. 52 Hinüber et al. demonstrated that the conduit fabricated from a poly(3-hydroxybutyric acid) (P3HB)/ PCL blend demonstrated superior flexibility and adequate mechanical strength compared with conduits made of pure P3HB or PLA. 53 In addition, in vitro experiments indicated that mouse fibroblasts were able to survive and proliferate within sealed porous tubes. An in vitro model of axon regrowth, utilizing DRG and sympathetic cervical ganglia, revealed that the NGCs effectively enhanced neurite regrowth. The incorporation of fibrillar lumen fillers facilitated oriented neurite growth, whereas coating with ECM proteins further enhanced ganglia attachment and cell migration. 53 This study demonstrates the potential of P3HB-based NGC scaffolds for long-gap peripheral nerve repair strategies, suggesting that PCL has good mechanical compatibility.
Importantly, the architectural design of fibers within NGCs plays a pivotal role in guiding axonal elongation and SC migration. Neuronal differentiation encompasses several specific events, such as the formation and elongation of neuritic processes, thereby highlighting the significance of establishing an aligned fibrous template to facilitate cell extension and elongation. 54 An aligned topographical structure represents a versatile and well-established straightforward strategy to enhance nerve regeneration, even in the absence of additional biological or physicochemical stimuli.54,55 Current strategies primarily focus on three types of fiber arrangements, including aligned fibers, random fibers, and gradient or hierarchical structures. First, the aligned fibers mimic the native endoneurial tubes of peripheral nerves, providing topographical cues for directional axonal growth. For example, aligned poly (l-lactic acid) nanofibers significantly enhance SC alignment and accelerate functional recovery in rat sciatic nerve models. 48 Additionally, in vitro cell culture studies indicate that aligned fibrous scaffolds offer improved cellular phenotype, increased cell proliferation, and enhanced neurite outgrowth compared with randomly oriented nanofibers. 54 Specifically, the cultivation outcomes of rat neuronal-like cells (B65 cell line) on poly (3-hydroxybutyrate) (PHB)/chitosan electrospun were aligned, and random fibrous scaffold was compared. The Methylthiazolyldiphenyl-tetrazolium bromide (MTT) assay results demonstrated enhanced proliferation of B65 cells on the electrospun-aligned PHB/chitosan scaffolds, suggesting that aligned fibrous scaffolds have robust abilities to promote nerve repair. 54 Second, while less effective in guiding axonal directionality, random fiber networks promote cellular infiltration and angiogenesis due to their high porosity. However, they may lead to disorganized axonal regrowth in large-gap injuries. 56 Finally, emerging designs incorporate gradients in fiber density or biochemical cues (e.g., neurotrophic factor concentration) to recapitulate the spatial heterogeneity of native nerves. For instance, a recent study demonstrated that chitosan conduits with radially aligned inner layers and porous outer layers improved both mechanical support and nutrient diffusion. 56 Although aligned fibers are widely adopted for their directional guidance, their static nature may limit adaptability to dynamic injury microenvironments. Future studies could explore hybrid designs combining aligned core fibers with bioactive outer layers to balance structural guidance and microenvironment modulation. Additionally, future NGC designs should prioritize dynamic fiber architectures, such as 4D-printed conduits capable of postimplantation shape adaptation. For example, stimuli-responsive hydrogels that transition from random to aligned structures under physiological cues (e.g., pH or enzymatic activity) could better mimic the temporal evolution of nerve regeneration. Additionally, integrating computational modeling to predict optimal fiber density and alignment patterns based on injury-specific parameters may accelerate personalized conduit development.
Evaluation methods for the efficiency of NGCs
It is important to recognize that the most important requirement for NGCs is clinical applicability. When designing NGCs with maximized efficiency, several requirements should be taken into account. First and foremost, a nerve conduit must be biocompatible, meaning it should not induce any mutations or carcinogenic, nor should it trigger inflammatory reactions. Synthetic piezoelectric polymers are generally biocompatible, and incorporating natural polymers can improve this property. 52 In addition to biocompatibility, NGCs must have biodegradability, ensuring that their absorption rate in the tissue is coordinated with the speed of nerve recovery. Once the scaffold is sutured at the damaged nerve, the biodegradable scaffold can be rebuilt, ultimately being substituted by the original nerves. The biodegradable speed is crucial for NGCs, as it is supportive for nerves during the entire regenerative process. Importantly, merging piezoelectric polymers with other polymers such as chitosan can change the speed of degradation. 54 Additionally, several methods are used to evaluate the efficacy of NGCs 52 : (1) assessing axonal regrowth via histological cross-sections of regenerated nerves; (2) measuring neuro-electrophysiological parameters, such as compound muscle action potentials (CMAPs); (3) evaluating motor functional restoration through behavioral metrics, such as rat gait analysis (e.g., footprint patterns) and the SFI scores; and (4) analyzing morphological changes and measuring the wet weight of distal gastrocnemius muscles. 57
Surface topography and water wettability of materials
The surface topography of scaffold materials is key to cell adhesion and proliferation. 58 Xia et al. developed a polylactic acid (PLLA) scaffold decorated with polydopamine (PDA) through piezoelectric charges produced by ultrasound. 59 Leveraging the electrostatic attraction between dopamine (DA) molecules and the piezoelectric charges stimulated by ultrasound, the in situ modification on PLLA nanofiber retained its nanofibrous structure. This engineered nanofibrous scaffold significantly promoted the proliferation and differentiation of neural stem cells cultured in the standard medium by day 7. 59 It is important to highlight that only under conditions with highly favorable adhesion, characterized by sufficient protrusion and pseudopodia, can the topography of oriented nanofibers be readily captured, thereby inducing clear directional growth of cells along the aligned nanofibers and achieving the desired neural cell morphology. This study illustrates the relationship between cell adhesion and the impact of electro-topographical cues on neural stem cells, which could inform the design of electroactive scaffolds in the future.
The water wettability of a nerve conduit material surface, which refers to its hydrophobicity or hydrophilicity, is a critical factor affecting cell–material interactions and subsequent cell behaviors. 60 Surface wettability is significantly influenced by the presence of surface functional groups and the surface roughness of the material.60,61 Generally, cells exhibit a preference for surfaces with moderate hydrophilicity for adhesion and growth. In contrast, polymer surfaces that are superhydrophilic (contact angle below 5°) or superhydrophobic (contact angle above 150°) are not conducive to cell attachment and growth. 52 Regrettably, for example, most piezoelectric polymers, such as polyvinylidene fluoride (PVDF), PLLA, and polyhydroxyalkanoates, are hydrophobic. 59 The hydrophobic nature of their surfaces hinders sufficient cell adhesion. To address this issue, methods such as plasma treatment, chemical hydrophilic functionalization, or polymer blending are commonly employed to enhance the hydrophilicity of piezoelectric polymer scaffolds.59,62 Indeed, these treatments often necessitate the use of costly equipment or involve cumbersome procedures. 59 Moreover, the hydrophilicity of plasma-activated surfaces tends to deteriorate over time. Additionally, many hydrophilic treatments can compromise the porous structure of fibrous scaffolds, which is particularly problematic for electrospun scaffolds.62,63 An unsuitable surface treatment can even damage the nanofibrous structure, which is the most sensitive component for generating in situ electrical stimulation (ES) by sensing the subtle force of cell traction. 59 Therefore, it is crucial to develop an ideal hydrophilic nanofibrous surface that facilitates cell adhesion while preserving the intrinsic nanostructure of the scaffold.
Piezoelectric and conductive materials
Beyond biochemical cues, piezoelectric and conductive biomaterials have emerged as pivotal components in next-generation NGCs. 64 PVDF is the most extensively studied piezoelectric polymer, boasting a piezoelectric coefficient of 20 pC/N.65,66 As a ferroelectric material, PVDF exhibits a piezoelectric effect following poling treatment, a process that aligns the polymer’s dipole moments to enhance its piezoelectric properties. 67 PVDF converts mechanical stimuli into electrical signals, mimicking endogenous bioelectricity to guide SC migration and axonal growth. 68 In addition, piezoelectric substrates, which generate negative and positive surface charges, have been shown to enhance metabolic activity and promote the maturation of neurons in primary cultures. 69 Emerging evidence suggests that PVDF with a nanoscaled stripe array that can produce surface piezoelectric potential through cell traction directed the neuron-like differentiation of rat bone marrow MSCs (rbMSCs). 58 The rbMSCs on the PVDF stripe array differentiated into β-III tubulin-positive neuron-like cells, a phenomenon not observed in cells on the nonpiezoelectric polyvinyl chloride stripe array. 58 This finding indicated that piezoelectricity tends to guide rbMSCs toward neuronal differentiation. Javidi et al. designed a new core/shell scaffold. 70 The coelectrospinning technique was employed to fabricate a fibrous shell composed of PCL/PVDF, gelatin, and polyaniline/graphene (PAG) nanocomposite. The major part of the scaffold was decorated with a chitosan-gelatin hydrogel loading PAG and zinc oxide nanoparticles. This scaffold exhibited antibacterial properties, electrical conductivity, and piezoelectric characteristics. The impact of the scaffold on cell differentiation was assessed by examining the expression of differentiation markers Nestin and microtubule-associated protein 2 (MAP2). These findings indicated that the scaffold notably enhances Nestin and MAP2 genes in PC12 cells. 70 Taken together, these studies shed new light on the application of piezoelectric polymers to the neuronal scaffold by regulating cellular behavior. The mechanism underlying the promotion of nerve tissue regeneration by piezoelectric scaffolds is quite intricate and likely involves a variety of factors. The success of piezoelectric stimulation in nerve regeneration can be attributed to the unique electrical capacities of neural cells. The membrane of neural cells is characterized by a negative charge on the intracellular side and a positive charge on the extracellular side. 71 Consequently, even without the neural transmission, a resting membrane potential of approximately 70 mV exists. Piezoelectric stimulation induces polarization of signaling molecules, leading to asymmetrical activation of these molecules and the downstream cytoskeleton, thereby guiding cell movement within the electric field (EF). 72 Movements toward the cathode or anode are triggered by ion channels NaKA and NHE3. 52 During exposure to the EF, NaKA and NHE3 accumulate at the cathodal or anodal edge of the migrating cell. 52 This activation, involving Na+ and Ca2+ influxes, creates an ion gradient along or against the EF, resulting in cell depolarization and a redistribution of the cytoskeleton.
Concurrently, conductive scaffolds such as polypyrrole (PPy) establish an electroactive microenvironment that regulates ion channel dynamics and neurotrophic factor secretion, synergistically accelerating functional recovery. Basurto et al. evaluated the efficacy of collagen-glycosaminoglycan (CG) scaffolds in promoting functional muscle recovery in a rat tibialis anterior model of volumetric muscle loss. 73 Functional recovery was assessed by in vivo ES of the peroneal nerve at 4, 8, and 12 weeks posttrauma, comparing nonconductive CG scaffolds with electrically conductive CG-PPy scaffolds. After 12 weeks, posttreated tissue exhibited significantly higher maximum isometric torque compared with untreated controls. Scaffold-treated muscles also showed increased infiltration of M1 and M2 macrophages within the damage site. Notably, conductive CG-PPy scaffolds demonstrated enhanced neovascularization. 73 Taken together, these data suggest that NGCs decorated with various piezoelectric and conductive materials have the potential nerve repair abilities by regulating cellular behaviors. Currently, piezoelectric patches (such as BTO@PCL/GO@GelMA) and conductive conduits (such as MF-NGC) have achieved repair outcomes comparable to autologous transplantation in animal models, with some entering the clinical trial phase. However, the long-term biocompatibility of piezoelectric materials (e.g., the inflammatory risk associated with PVDF degradation products) and the mechanical stability of conductive materials (e.g., the fatigue fracture of graphene) need to be resolved. 52 Additionally, by integrating 4D printing technology, dynamic response conduits have been developed to adapt to the mechanical demands of various damaged areas. Taken together, piezoelectric and conductive materials, by simulating physiological electrical signals, regulating the immune microenvironment, and activating key pathways, have demonstrated irreplaceable roles in nerve regeneration. In the future, further exploration of their synergistic mechanisms and promotion of clinical translation research are required.
3D and 4D bioprinting
Traditional methods for developing NGCs include solvent casting, phase separation, gas foaming, freeze-drying, and electrospinning, yet these methods have shortages such as the generation of random and highly disordered fiber and low porosity.74,75 Recently, 3D printing has gained popularity due to its customizability, overcoming the limitations of traditional methods in developing NGCs. 76 3D bioprinting, an additive manufacturing-based technology, can create biomimetic nerve scaffolds that are customized for patients in a layer-by-layer manner, allowing for precise transplantation according to the shape and length of damaged nerves. 3D printing requires bio-inks, which mainly include bio-inks and decellularized inks. Bio-inks, decorated with agents, factors, and molecules, must be developed in a permissive milieu for cell survival, 44 whereas decellularized inks can withstand harsher milieus, making them more preferable for 3D printing. 44 Evidence from several studies already indicates that the application of soft polymers with good mechanical strength in 3D bioprinting drives better regenerative outcomes as the elastic modulus of the human brain, spinal cord, and peripheral nerves are 1, 10, and 100 − 500 kPa, respectively. 77 The basic biomaterials for 3D printing include polymeric hydrogels, poly(lactide-cocaprolactone) (PLCL), and nanofibers.78,79 Jin Yoo et al. 78 designed oriented collagen hydrogels using 3D printing. Furthermore, researchers developed elastic NGCs based on PLCL. In this study, the use of NGCs combined with 3D-printed hydrogels significantly promoted myelin sheath reestablishment and axonal elongation upon sciatic nerve damage in rats. Zhang et al. developed a micron track conduit decorated with chitosan and delivered neurotrophin-3 into the conduit. The results showed that this customized conduit supported the directed migration of SCs and enhanced peripheral nerve regrowth postinjury. 80 Taken together, 3D bioprinting technology provides promising potential for reconnecting large nerve gaps. However, it also has potential shortcomings, such as limited printing speed and poor printing accuracy primarily due to the immature technology of the nozzle. These limitations hamper the widespread application of 3D bioprinting. Thus, emerging technology such as 4D bioprinting has been proposed to create more complex and elaborate constructions. 4D printing extends traditional 3D printing by incorporating materials that undergo time-dependent, stimulus-responsive transformations postfabrication. 81 While 3D printing creates static scaffolds with predefined geometries, 4D printing enables dynamic changes in material properties (e.g., shape, stiffness, porosity) in response to external stimuli such as temperature, humidity, light, or magnetic fields. 82 This technology integrates 3D-manufactured structures with biomimetic actuating mechanisms to program the timing of shape transformation, enhancing physiological relevance. Three primary methods achieve these changes: (1) using thermoresponsive shape-memory polymers that revert to a predefined shape upon heating; (2) leveraging multimaterial structures with mismatched properties (e.g., swelling ratios or thermal expansion coefficients); and (3) exploiting stress gradients between layers in multilayer composites. 83 It should be recognized that the implants fabricated by 3D printing will cause secondary damage to injured nerves owing to their static nature, whereas 4D printing provides an exciting opportunity to create dynamic shape-changing structures using stationary components while leveraging the benefits of near-net-shape manufacturing for intricate and tailored parts. 84 The optimal material for 4D printing is hydrogels because they swell after water uptake. Joshi et al. proposed 4D printable hydrogels programmed to undergo predetermined shape changes in vivo following 3D printing for tissue regeneration. 84 The bicomponent alginate and methyl cellulose (A/MC) hydrogels, featuring varied A/MC ratios, were 3D printed along distinct layers with specific filling patterns, enabling them to autonomously alter their initial printed forms while triggering though the absorption of water. The gels decorated with gelatin-rich nanofibers enhanced cell adhesion. The 4D-printed tubular structures were achieved via in vivo controlled shape deformation of the 3D-printed gels, serving as intraoperative self-folding NGCs. 84 Subsequently, histological and functional assay results were evaluated to determine the ability of the 4D-printed conduits to regenerate peripheral nerve injuries. Specifically, the staining results of the cross-sections and longitudinal sections of the sciatic nerve with NF-160 (a neurofilament marker), S-100 β (a SC marker), and nuclei indicated that the 4D-printed conduits significantly promoted the proliferation of these markers, 84 suggesting a robust nerve regenerative ability of this conduit. Recently, a nerve scaffold featuring aligned microgrooves was successfully developed utilizing the photolithography-stereolithography tandem printing technique. 85 The integration of photolithography and stereolithography printing contributed to well-defined micropatterns on the printed scaffolds, which provided significant advantages for guiding oriented cell growth. A mussel-inspired PDA coating was subsequently applied to immobilize GFs, thereby enhancing the neural differentiation of resident human MSCs. This study presents a new strategy for developing multifunctional 4D nerve conduits that combine topographic guidance and biological cues. 85 Taken together, these studies have illustrated the potential of 4D-printed tissue constructs in peripheral nerve repair and regeneration, while also emphasizing the promising prospects of 4D printing for additional applications within regenerative medicine.
4D-manufactured structures remain several challenges despite they exhibit favorable promoted regeneration capacities: (1) material biocompatibility: long-term safety of smart materials such as residual nanoparticles requires validation; (2) stimulus precision: overexposure to external triggers such as heat may damage adjacent tissues; and (3) scalability: current 4D printers lack the resolution and speed for large-scale clinical use. Beyond the integration of electrical cues, other cues, such as mechanical and biological factors, have not been incorporated into 4D-manufactured NGCs. Further studies should aim to develop multifunctional NGCs that meet specific clinical requirements. A variety of neuro-GFs conjugated with bioactive materials can act as biological links for the production of advanced dynamic NGCs, capable of ensuring rapid regeneration. For hollowed-out conduits, 4D manufacturing technology could be designed for more complex structures. The in situ monitoring of the conduit represents a significant clinical challenge. With the latest advances in materials science, attention should be paid to the development of radiopaque bio-link/4D materials that can be easily observed under X-rays. Moreover, self-healing materials can be integrated with advanced 4D manufacturing technology to ensure enhanced patency of the conduit after implantation. Mathematical models are an indispensable part of 4D manufacturing. While recent studies have confirmed the efficacy of mathematical models in predicting shapes, their application in ensuring a perfect fit with damaged nerve endings upon in vivo implantation warrants more stringent investigation. 85 These advancements can help develop NGCs with exceptional patient-specific fit, minimize surgical complexity, and provide a paradigm shift from the gold standard that typically requires sutures to place NGCs.
Table 1 lists various synthetic NGCs for PNR. NGCs demonstrate significant potential for bridging peripheral nerve defects, yet their clinical translation remains limited by unresolved challenges. For example, chitosan-based conduits exhibit superior biocompatibility but lack the mechanical strength required for long-gap repairs in load-bearing nerves. Conversely, synthetic polymers such as PLCL provide robust structural integrity but risk chronic inflammation due to slower biodegradation rates. A critical question emerges: Can hybrid materials—such as chitosan-PLCL composites—achieve a balance between biodegradability and mechanical strength while minimizing immune responses? In addition, it is important to recognize that although various NGCs containing beneficial cues have been developed to boost peripheral nerve regrowth in recent years, they cannot replace autografts, which remain the gold standard of PNI treatment. However, some in vivo and in vitro studies indicate that sophisticated NGCs designed to mimic the topographical and biochemical features of peripheral nerves can significantly promote neuronal extension, achieving results comparable to autografts.45,46 Thus, future generation of NGCs, incorporating 4D bioprinting, may potentially substitute autografts and yield even better outcomes of PNR.
Various Nerve-Guided Conduits for Nerve Regeneration
ATF3, activating transcription factor 3; BDNF, brain-derived neurotrophic factor; DTI, diffusion tensor imaging; NGC, nerve-guided conduit; NRG1, neuregulin 1 gene; PLLA, poly (
Gene therapy has also been utilized to boost nerve reconstruction. Gene therapy involves introducing nonnative and therapeutic genes into living cells to deliver novel treatments or modify the cell’s typical physiology. 86 In the context of neuronal regeneration, gene therapy aims to promote repaired phenotypes or create a microenvironment within the damaged nerves that supports axonal outgrowth through SCs. 43 Applications of gene therapy indicate targeted delivery to DRG and SCs. To enhance the self-regenerative capabilities of peripheral nerves, gene therapy should target DRG or the ventral horn of the spinal cord. Adeno-associated viral vectors have proven effective for gene transfer and are preferred in human applications due to low immunogenicity and good gene transfer capabilities. 87 Gene therapy targeting the NTF gene can enhance its expression, which in turn triggers axon extension, yet it presents a drawback: Excess NTF may trigger a detrimental feedback mechanism that hinders axon outgrowth. 88 These findings imply that future research is needed to optimize the therapeutic effects of neuronal outgrowth. Consequently, integrating 3D- or 4D-printed NGCs with gene therapy emerges as a promising strategy for treating PNI.
Conservative treatment
MSCs and extracellular vesicles
MSCs are multipotent stem cells produced by various body tissues, such as bone marrow, adipose tissue, umbilical cord, amniotic membrane, human placenta, dental pulp tissue, and skeletal muscles.89–91 Among these, ADSCs are recognized for their superior biocompatibility and popularity. 91 Thus, ADSCs are usually used in peripheral nerve defect treatment and neuropathic pain mitigation. MSCs can release diverse anti-inflammatory and neurotrophic factors in a paracrine or autocrine manner. After implantation combined with tissue engineering in injury nerves, they not only differentiate into SCs but also induce the release of extracellular vesicles (EVs) and exosomes. 92 Emerging evidence suggests that transplanting ADSC-derived mitochondria dramatically alleviates erectile dysfunction elicited by cavernous nerve injury and promotes smooth muscle content. In vitro, ADSC-derived mitochondria transplantation downregulates the expression of reactive oxygen species and cleaved-caspase 3 but upregulates superoxide dismutase and adenosine triphosphate (ATP) expression. 93 Overall, this study indicates that ADSC transplantation into cavernous nerve injury rats can significantly mitigate the symptoms of erectile dysfunction by functions of antioxidative stress and antiapoptosis. 93 Other researches concerning the clinical application of MSCs in PNR are summarized in Table 2.
Summary of Studies Concerning the Application of Mesenchymal Stem Cells in Peripheral Nerve Regeneration
ADMSCs, adipose-mesenchymal derived stem cells; DPSCs, dental pulp stem cells; GMSCs, gingiva-derived mesenchymal stem cells; PMSCs, placental mesenchymal stem cell; sEVs, small extracellular vesicles; SMSCs, skin-derived mesenchymal stem cells; UCMSCs, umbilical cord mesenchymal stem cell.
EVs are nanosized (50–200 nm) vesicles with a lipid bilayer membrane; they derive from diverse types of cells and enhance intercellular communication. As mentioned earlier, MSCs have already been identified as a useful tool for promoting nerve regeneration. However, the therapeutic potential of human umbilical cord MSC-derived EVs (hUCMSC-EVs) in PNI treatment has yet to be fully confirmed. However, Ma et al. 94 reported that the use of these special-derived EVs improved SFI and reduced gastrocnemius atrophy in vivo within sciatic nerve cut injury rat models. In this study, hUCMSC-EVs accumulated at the injured nerve sites, reducing interleukin-6 (IL-6) and IL-1β expression while enhancing IL-10 expression. 94 These findings suggest the therapeutic potential of hUCMSC-EVs in PNI. In addition to hUCMSC-EVs, the small EVs (sEVs) isolated from human placenta-derived MSCs (hPMSCs-derived sEVs) were also able to alleviate mechanical hypersensitivity after sciatic nerve defects in rat models. 95 This study also reported that intrathecal injection of miR-26a-5p agomir contributed to neuropathic pain mitigation by activating Wnt5a, whereas overexpression of miR-26a-5p downregulated Wnt5a. 95 Thus, these data suggest a favorable possibility of applying hPMSCs-derived sEVs to promote PNR. While both hUCMSC-EVs and hPMSC-sEVs demonstrate potential in PNI repair, their mechanisms differ significantly. hUCMSC-EVs primarily modulate inflammatory cytokines (e.g., IL-6 and IL-1β reduction), 96 whereas hPMSC-sEVs target miRNA-mediated pathways (e.g., miR-26a-5p/Wnt5a axis). 95 These differences suggest that the choice of EV source may depend on the specific pathological context (e.g., neuroinflammation and axonal guidance). Notably, none of these studies directly compared the efficacy of hUCMSC-EVs and hPMSCs-derived EVs in the same experimental model, raising questions about their relative advantages. Furthermore, while both strategies are cell-free and avoid immunogenicity concerns, scalability and standardization remain challenging. For example, hPMSCs-derived sEVs require complex isolation protocols compared with hUCMSC-EVs, which may hinder clinical translation. 95
Exosomes, a subset of EVs with a diameter of 30–200 nm, 97 are isolated from various cells through interactions with intracellular vesicles and the plasma membrane. 98 Their compositions include nucleic acids, proteins, cytokines, mRNAs, and miRNAs, which partially mirror the properties of their parent cells. Exosomes can deliver cargoes to target cells. Mounting evidence indicates their therapeutic potential in facilitating PNR. For example, Zhao et al. 17 reported that BMSC-derived exosome treatment in vitro significantly upregulated the levels of PMP22, VEGFA, NGFr, and S100b of DRG compared with phosphate-buffered solution treatment following a rat sciatic nerve defect. In vivo, high-dose administration of exosomes into the gastrocnemius muscles of nerve-injured rats improved SFI and latency of thermal pain, suggesting that exosome treatment after sciatic nerve defects facilitated axonal elongation and nerve functional recovery. 17 Additionally, the differentiation of ADSCs into SCs, elicited by SC-derived exosomes, was mediated by miRNA and the PI3k signaling pathway, 99 both of which were also involved in the process of PNR facilitated by endothelial cell-derived exosomes. 32 Moreover, Liu et al. 98 developed two types of exosomes, uExo and dExo, derived from human adipose-derived MSCs and dedifferentiated hADMSCs, respectively. The results indicated that dExo had a greater ability to modulate inflammatory microenvironment, elevate the levels of miRNA, and promote axonal regrowth, thus accelerating nerve functional recovery. Taken together, these findings provide important insights into using cell-free strategies to promote PNR, yet critical gaps persist. For instance, discrepancies in dosing regimens and delivery methods across studies hinder direct comparisons. Zhao et al. administered exosomes via gastrocnemius muscle injection, 17 while Huang et al. delivered them beneath the sciatic nerve epineurium, 32 complicating direct comparisons. A unified framework for dosing, timing, and administration routes is urgently needed to validate these strategies. Additionally, the efficacy of exosomes may depend on context: while they excel in acute inflammation regulation, their inability to provide structural support necessitates a combination with NGCs. Detailed safety data on exosome surface modification and gene loading are lacking, and clinical side effects are still unknown. 32 Furthermore, their clinical use has ethical and safety problems due to challenges in sourcing (e.g., allogeneic vs. autologous) and risks of contamination with nonexosome proteins, which have not been fully addressed in studies. 100 Even autologous MSC-derived exosomes face scalability issues, particularly for early nerve injury treatment. 99 Mechanical stimulation and dynamic culture systems are thus critical to enhance exosome generation, although parameters must be tightly controlled to resolve the unbalance of inflammatory factors or shedding of nonexosome vesicles. Despite their promise in nerve repair, exosomes remain a novel therapeutic agent in tissue engineering. Further research on clinical efficacy and scalable production is essential prior to applying it to human safely.
Platelet-rich plasma
Platelet-rich plasma (PRP) is produced from autologous blood through powerful centrifugation and comprises various neurotrophic molecules, adhesion molecules, and GFs, including NGF, brain-derived neurotrophic factor (BDNF), IGF-1, platelet-derived growth factor, VEGF, and TGF-β. 101 PRP has been widely used in clinical and fundamental research for decades102–106 due to its safety, convenient accessibility, and extremely low immunogenicity. PRP can remarkably boost PNR through six functions: neuroprotection and prevention of cell apoptosis, promotion of angiogenesis, facilitation of axonal extension, amelioration of the inflammation microenvironment, suppression of the denervated target muscle atrophy, and improvement of human nervous system parameters. 107 For example, PRP creates a supportive anti-inflammatory microenvironment to promote PNR. In vitro, the application of platelet-rich GFs (PRGF) that are produced by PRP in M0 macrophages (unpolarized state) blunts the expression of tumor necrosis factor-α, IL-1β, and IL-6 but upregulates CD206 expression, 108 suggesting a potential transition of M0 to M2 macrophages after using PRGF. In addition, Yoshihiro et al. 109 reported that SC proliferation and migration and nerve growth-related factors generation were enhanced by 5% PRP administration in sciatic nerve defect rat models. This study suggests the therapeutic potential of using PRP to facilitate PNR by modulating SC functions. PRP also exhibits robust therapeutic efficacy in PNI through combination with tissue engineering. Ye et al. 110 developed a poly (lactic-coglycolic acid) conduit composed of PRP and differentiated bone marrow MSCs. They then implanted this conduit into sciatic nerve cut injury rabbit models, and the results showed an increased number of regenerating nerve fibers and myelin sheath thickness and improved CMAP and nerve conduction velocity after 12 weeks. Inside-out vein graft can greatly bridge small-gap nerve deficits. By contrast, Ji Yeong Kim et al. 111 reported that PRP-seeded inside-out vein graft exhibited more myelinated axons and thicker myelin sheath after 6 weeks. In addition, Hadi Samadian et al. 112 used the PCL/gelatin nanofibrous NGCs seeded with PRP and citicoline in critical-size sciatic nerve damage rat models, and the results showed improved SFI and hot plate latency time, suggesting that PRP-seeded NGCs can significantly promote nerve regrowth and motor and sensory function recovery. Lu et al. 113 reported that chitin biological conduits combined with small autogenous nerves and PRP increased the number of regrowth axons and improved composite action potential intensity, myelinated nerve fiber density, and gastrectomy tissue condition 2 weeks after implantation in sciatic nerve deficit rat models. Additionally, Zhu et al. 114 conducted a study in which 75 rabbits were divided into 5 groups: hollow, microtissues, PRP, microtissues + PRP, and autografts. The group of microtissues + PRP and autografts then exhibited significant outcomes of nerve regeneration such as promoted proliferation, migration, and growth-related factor secretion of SCs compared with other groups 12 weeks after surgery. Moreover, Dong et al. 115 developed a synthetic NGC composed of PRP, gelatin methacrylate, and sodium alginate hydrogel and found this NGC could greatly repair a 10 mm sciatic nerve defect in rats and improve functional recovery. Furthermore, it contributed to blood vessel formation and axon elongation in vitro through the sustained release of growth-related factors such as VEGF. 115 Table 3 summarizes various studies using PRP to facilitate PNR. Taken together, these reports demonstrate that PRP and various NGCs combine to facilitate PNR. Notably, most studies on PRP and NGCs have employed rodent sciatic nerve injury models with gaps ≤15 mm. However, their translational relevance to human clinical scenarios—such as brachial plexus injuries with >50 mm gaps—remains uncertain, as larger animal models (e.g., nonhuman primates) are infrequently used.
Summary of Platelet-Rich Plasma for Peripheral Nerve Regeneration
IGF-1, insulin-like growth factor-1; PDGF-BB, platelet-derived growth factor-BB; PRP, platelet-rich plasma; PRGF, platelet-rich growth factors; SANs, small autogenous nerves.
Further studies concerning PRP and PNR require larger sample sizes, stringent experimental methods, and standardized assessment criteria. Furthermore, the unified source, timing, and dosage of intravenous injection of PRP are imperative to acquire more reliable and precise outcomes. While PRP-seeded NGCs demonstrate consistent benefits in rodent models (e.g., enhanced myelination and improved sciatic functional index [SFI]), several critical questions remain unresolved. First, the absence of standardized PRP preparation protocols—such as platelet concentrations ranging from 1 × 106 to 5 × 106/μL across studies—hinders cross-study comparability. Second, no clinical trials have evaluated PRP-NGC combinations, despite their preclinical efficacy. We propose that future research should: (1) develop consensus guidelines for PRP characterization and standardization; (2) investigate the potential synergy between PRP and dynamic 4D-printed conduits to achieve spatiotemporal control over growth factor release; and (3) assess and mitigate risks, such as ectopic fibrosis resulting from prolonged TGF-β exposure.
Electrical stimulation
ES has been identified as a highly promising approach to enhance axonal regrowth postinjury. In an in vivo experiment of rodent PNI models, 10 min of ES after therapeutic surgery significantly promoted motor functional recovery and axonal elongation, comparable to 60 min of ES after surgery. 116 Roh et al. reported that immediate ES following a tibial nerve isograft promoted macrophage recruitment and axonal elongation compared with groups without ES treatment. 117 These findings suggested that peripheral nerve regrowth could be promoted by the application of ES after repair surgery within rodent models of nerve damage, which makes the clinical translation of this approach more feasible. ES parameters, including field strength, frequency, and pulse duration, critically influence cellular responses and nerve regeneration outcomes. Emerging evidence demonstrates that neuronal maturation in human neuroblastoma SH-SY5Y cells is related to the amount of charge converged during ES. 118 A higher level of charge accumulation (∼ 50 mC/h) remarkably enhances neuronal growth and branching and increases the levels of synaptophysin, resulting in effects exceeding those of BDNF. In contrast, injecting a smaller amount of charge into the culture (∼ 0.1 mC/h) has minimal induction of maturation but notably enhances cell amplification. Additionally, ES modifies the protein levels of EV. ES with a sufficiently large accumulated charge notably enriches the proteome of the EVs related to neurodevelopment. 118 Koppes et al. also suggested that the combination of direct current (dc) ES and SCs might offer synergistic guidance signals, enhancing axonal growth relevant to the repair of nerve damage. 119 In their study, primary dissociated neurons from neonatal rat DRG and SCs were subjected to an 8-h dc ES (0–100 mV/mm). The results demonstrated that SCs remained viable after ES with 10–100 mV/mm and maintained a morphology similar to that of untreated cells. Neurite regrowth was notably enhanced twofold after exposure to either a 50 mV/mm EF or coculture with unstimulated SCs compared with neurons cultured alone. The presence of both SCs and a 50 mV/mm dc EF further enhanced neurite regrowth. Specifically, this combination resulted in a 3.2-fold increase compared with unstimulated control neurons and a 1.2-fold increase compared with neurons cultured with unstimulated SCs alone or with the electrical stimulus alone. 119 This study provides data on the values of EFs to enhance nerve regeneration. Frequency-dependent effects are also pivotal. While low-frequency ES (1–20 Hz) aligns with physiological nerve signaling, higher frequencies (up to 1000 Hz) modulate membrane polarization and ion channel dynamics. 120 In total, 110 patients with humerus shaft fractures and radial nerve injuries were randomly assigned to receive either conventional exercise treatment or combined low-frequency pulsed ES. 121 The clinical outcomes, muscle strength restoration, and amplitude, as well as a range of motion of the wrist and elbow joints, were analyzed and compared among the patients. The results showed that the treatment effectiveness of patients receiving low-frequency ES (89.09%) was significantly higher than that of patients who only received exercise therapy (69.09%). Compared with the single exercise intervention, the combination of LFS and exercise therapy can more effectively enhance the muscle strength of patients’ wrist extensors and finger extensors, suggesting that LFS facilitates better muscle function restoration. Six months after treatment, it was reported that a greater number of patients in the low-frequency ES group achieved wrist extension and elbow extension angles beyond 45° compared with those in the control group. There was no significant difference in adverse reactions between the two groups of patients. 121 Thus, combining exercise therapy with low-frequency pulsed ES can markedly enhance clinical efficacy, facilitate the recovery of nerve function and muscle strength, and ensure a high degree of safety. In addition, Power et al. reported that patients with cubital tunnel syndrome treated with 1 h of 20 Hz ES after surgery showed improved motor unit number estimation and grip and key pinch strength compared with those treated with surgery alone. 122
High-frequency ES (500–1000 Hz) has demonstrated efficacy in reducing neuropathic pain by suppressing hyperexcitability in nociceptive neurons; however, further investigation of its regenerative effects is required. 123 Collectively, optimal ES parameters are critical for promoting cellular activity and neuronal regeneration.
Recent research has focused on accelerating peripheral nerve regrowth by combining ES and tissue engineering124–126 For example, Xu et al. designed an aligned piezoelectric nanofiber-generated hydrogel NGC that could produce ES induced by ultrasound and the results illustrated that this NGC could enhance axonal elongation, 127 suggesting its potential for promoting peripheral nerve outgrowth. However, NGCs have exhibited shortcomings in many studies such as structural instability. Song et al. fabricated an electrospun shape-persistent conductive NGC. This NGC can not only maintain its original structure within the complex microenvironment of nerves but also possess better conductivity. 128 Interestingly, Hu et al. used bionic microneedle NGCs in cucumber peripheral never-damage sea models, which reduced target muscle atrophy. The outer surfaces of these NGCs featured outward-pointing needle tips that generated ES, aiding in atrophic muscle recovery. 129 Taken together, these studies demonstrate that a combination of ES and NGCs composed of various biomaterials shows promising potential for promoting axonal elongation, releasing NGFs, and preventing distal tissue atrophy. Evidence from several studies already indicates that low-frequency ES leads to more successful neuronal regeneration after damage. 43 However, the optimal frequency of ES remains undetermined, necessitating further research. In contrast, designing scaffolds decorated with biomaterials that show better conductivity is a current hot topic. Addressing this issue could drive more rapid and more successful peripheral nerve regrowth.
Traditional Chinese medicine
The potential of traditional Chinese medicine (TCM) in enhancing nerve regeneration has increasingly attracted attention compared with the above emerging technologies. From time immemorial, TCM has played a vital role in disease treatment. More and more active components with drug action in Chinese herbal medicine have been extracted and studied. Among the plenty of active components, resveratrol (Res) is regarded as a polyphenolic compound with promising prospects. It exhibits antioxidant and anti-inflammatory properties and is a potential agent for disorder treatment. Recently, Res has been identified to enhance peripheral nerve regrowth. Ding et al. 130 reported that Res increased the number of axons and promoted functional recovery via the p300-mediated VEGF signaling pathway in a systemic administration manner in rat models of sciatic nerve lesions. Zhang et al. 131 delivered Res into sciatic nerve crush injury rat models and measured the SFI, MAP light chain 3B, and myelin protein zero levels. They found that Res treatment facilitated SC autophagy to promote myelin removal and improved SFI, suggesting that Res may achieve successful neuronal regeneration by targeting SC functions. 131 Moreover, Zhang et al. 10 reported that the combination of Res and ADSCs showed that the average density of myelin sheath, and the number of motor neurons was significantly increased compared with model groups, which indicated that Res could amplify the proregenerative capacities of ADSCs for PNI.
Res has also been identified to ameliorate neuropathic pain following nerve lesions.132,133 For instance, Xu et al. 134 found that intraperitoneal administration of Res after sciatic nerve chronic constriction injury in rats contributed to the mitigation of thermal hyperalgesia and mechanical allodynia. Furthermore, Res treatment was found to upregulate the levels of several inflammatory factors such as IL-4Rα, IL-1RA, and IL-1R2. These data suggest that Res may help mitigate mechanical allodynia by promoting inflammatory response in the spinal cord. 134 Guo et al. 135 used partial sciatic nerve ligation to induce neuropathic pain in rat models and found that consecutive intraperitoneal administration of Res for 21 days could alleviate this pain, potentially through restraining P2X3 and ERK phosphorylation. Overall, Res carries out neuroprotection and analgesic functions to promote PNR and may serve as a potential therapeutic agent for PNI, but this requires further investigation. While Res consistently demonstrates neuroprotective effects in rodent models, its efficacy in nonhuman primate studies remains controversial. Notably, Res exhibits limited bioavailability in nonhuman primates, indicating that formulation optimization—such as nanoparticle encapsulation—may be required for clinical translation. 136
Centella asiatica (L.), another TCM, shows promising therapeutic potential for PNI. Hussin et al. 137 developed nerve conduits decorated with C. asiatica (L.)-differentiated MSCs to apply to 1.5 cm sciatic nerve defect rat models. The results showed that the transplantation of this nerve conduit exhibited higher nerve conduction velocity, myelin basic protein (MBP) expression, and myelin sheath thickness compared with the transplantation of conduits decorated only with MSCs, 137 suggesting that C. asiatica (L.) had neuroprotective functions. In fact, several active compounds produced by C. asiatica have also been investigated for many years, including asiaticoside, madecassoside, asiatic acid, and madecassic acid. 138 Many studies have indicated that these have promising therapeutic effects on CNS disorders.138,139 However, there is a lack of studies focused on treating peripheral nerve defects, which should attract more attention from researchers.
In addition to Res and C. asiatica, other TCMs such as Achyranthes polypeptide and salidroside have also been found to have neuroprotection effects in recent years. For example, Cheng et al. reported that Achyranthes bidentata polypeptide k (ABPPk), the most active ingredient of Achyranthes polypeptides, enhanced neuronal regrowth in vitro, assessed by immunocytochemistry and western blot analysis, which may be mediated via Erk1/2 singaling. 140 In vivo, they found that ABPPk improved motor functional recovery and axonal elongation in rats. These results indicate that Achyranthes polypeptide may be a potential agent to carry out neuro-treatment effects. In addition, Liu et al. fabricated a nerve conduit composed of salidroside, PLGA, and SCs and found that salidroside significantly promoted SC proliferation and migration in vitro. 141 They then applied this nerve conduit to sciatic nerve defect rat models, and the results showed that it increased SFI and enhanced nerve conduction, which suggests that the combination of salidroside and tissue engineering has promising potential for PNI repair. 141 Table 4 summarizes several TCMs that facilitate PNR.
Summary of Traditional Chinese Medicines for Peripheral Nerve Regeneration
ADSCs, adipose-derived stem cells; EPK, extracellular regulated protein kinases; IL, interleukin; SFI, sciatic function index; VEGF, vascular endothelial growth factor.
Building on the prior discussion, we propose the following research questions. C. asiatica (L.) and Achyranthes polypeptides promote regeneration through distinct pathways. Hussin et al. demonstrated that C. asiatica upregulates MBP, 137 while Cheng et al. linked Achyranthes polypeptides to Erk1/2 activation. 140 Do these pathways exhibit additive or antagonistic effects? Coadministration studies in vitro (e.g., DRG neurons treated with both compounds) could elucidate their interactions. Additionally, TCM faces significant barriers to global clinical translation, such as batch-to-batch variability in herbal extracts (e.g., inconsistent salidroside content in Rhodiola rosea). A lack of FDA-compliant pharmacokinetic and pharmacodynamic data for most TCM compounds further complicates translation. The in vivo administration route and method of these medicines warrant careful consideration. Oral administration is cost-effective and relatively safe but may yield variable efficacy because of the unpredictable feeding habits and behaviors of animals. However, gavage or injection routes require restraint, which may induce stress in animals and influence study outcomes. Treatment schedules also vary widely, with durations ranging from days to months, and treatment frequency can significantly impact results. 142 While standardizing animal handling remains challenging, these factors must be systematically addressed in study design.
Conclusions and Perspectives
PNI can lead to critical consequences and requires intervention timely to prevent chronic denervation. This review highlights the importance of SCs and macrophages in nerve damage and regeneration. Current available strategies, including surgical and nonsurgical approaches, promote peripheral nerve reconstruction partly by modulating the functions of these cells. One of the most promising approaches is the use of NGC, which has shown robust potential in promoting PNR when combined with various cells, molecules, cytokines, and even amniotic membranes. However, the characteristics of NGCs, such as their biocompatibility and biodegradation, should be carefully considered to achieve successful regenerative outcomes. Furthermore, while NGCs developed through 3D bioprinting technology remain a subject of deep research, the advent of 4D bioprinting technology may change the research direction in the future. Additionally, compared with other treatments, clinical cases involving NGCs are relatively scarce, making this an area of ongoing research. Meanwhile, TCM, electric stimulation, and exosomes have also been identified as favorable strategies owing to their minimal maladaptive symptoms. Despite promising preclinical data, critical gaps persist. First, a direct comparison of EVs and PRP in the same injury model has not been conducted. Moreover, combinatorial approaches, such as EVs combined with ES, remain understudied, despite their potential for synergistic effects.
It is important to recognize that even though these options exhibit promising therapeutic potential, no method can achieve complete functional restoration. Several options, such as gene therapy, are still in the preliminary phases of clinical research. Furthermore, most previous studies have primarily focused on singular signaling pathways. Urgently needed are further in vitro and in vivo trials to explore potential therapeutic agents that can address the limitations of current PNI treatments. Additionally, developing standardized animal models and appropriate evaluation criteria may help ensure the accuracy of scientific research. Notably, few studies have focused on the mechanisms by which regrowing axons stop extending once they reach the target tissue and form connections. Thus, further investigations into these mechanisms may enhance our understanding of PNR.
Taken together, the process of PNR involves an integrated multidisciplinary approach, including pharmacology, tissue engineering, and biology. However, current knowledge is limited and does not enable us to achieve perfect outcomes. Key limitations include: (a) material limitations of NGCs: Despite progress, most synthetic nerve conduits (NGCs) exhibit inadequate biodegradation rates and mechanical mismatches with native nerves. For instance, PCL-based conduits often exhibit excessively slow degradation (>18 months), exacerbating chronic inflammation and fibrosis in long-gap injuries. 143 Future strategies could leverage hybrid materials—combining fast-degrading polymers (e.g., PLGA) with decellularized matrices—to synchronize degradation with axonal regrowth while minimizing foreign body reactions. (b) Unresolved mechanisms of axon-target reconnection: Even successful axonal elongation frequently fails to restore precise neuromuscular synapses, leading to incomplete functional recovery. 4 This stems partly from an insufficient understanding of how regenerating axons recognize and terminate at target tissues. Single-cell RNA sequencing of regenerating neurons and SCs may identify novel guidance cues (e.g., semaphorins, netrins) to enhance synaptic specificity. (c) Clinical translation barriers: Promising therapies, such as MSC-derived exosomes and 4D-printed conduits, lack standardized protocols for scalable production and delivery. For example, exosome dosing and biodistribution in humans remain poorly defined.
Collaborative consortia should establish good manufacturing practice guidelines for stem cell products and biomaterials to ensure reproducibility. (d) Incomplete understanding of immune modulation: Macrophages, critical for debris clearance, dynamically influence regeneration via polarization states (M1/M2). 144 Overactivation of proinflammatory M1 macrophages exacerbates fibrosis, while their presence during the early injury phase is essential for resolving inflammation. 145 Strategies to precisely modulate their phenotype in vivo are lacking. Developing NGCs loaded with nanoparticle-delivered anti-inflammatory cytokines (e.g., IL-4, IL-10) could locally skew macrophage polarization toward proregenerative M2 states during critical stages, enhancing regenerative outcomes. To address these gaps, future research should prioritize advanced biomaterial design, such as integrating stimuli-responsive 4D printing with bioactive cues (e.g., GDNF gradients) to create dynamic, patient-specific NGCs. Leveraging omics technologies to map spatiotemporal molecular interactions during regeneration could also facilitate mechanistic insights. Optimizing clinical trials—via standardized animal models (e.g., nonhuman primates for long-gap studies) and functional outcome metrics (e.g., kinematic gait analysis)—is critical for translational relevance. Systematically targeting these challenges could transition the field from incremental improvements to transformative therapies restoring full sensory-motor function in PNI patients.
Footnotes
Authors’ Contributions
C.L. contributed to conception and design, acquisition of data, analysis and interpretation of data, and draft the article. S.L. contributed to acquisition of data, analysis, and interpretation of data. X.M. and C.W. contributed to review critically for important intellectual content and give final approval of the version to be published. All authors contributed to editorial changes in the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Natural Science Foundation of Heilongjiang Province (JJ2024LH0379).