
Abstract
Repair of the tendon–bone interface (TBI) remains a significant clinical challenge due to its complex biomechanical environment and hierarchical structure. Conventional surgical approaches often fail to fully reestablish native tissue architecture and function. In recent years, tissue engineering strategies have increasingly emphasized the application of mesenchymal stem cells (MSCs) and platelet-derived products to promote regeneration. MSCs possess multilineage differentiation potential and immunomodulatory capabilities, making them attractive candidates for TBI repair. Platelets, through their rich secretome, orchestrate essential regenerative processes such as cell recruitment, angiogenesis, and immune modulation. This review explores the molecular crosstalk between MSCs and platelets, critically examines current approaches utilizing platelet-rich plasma (PRP)–MSC combinations and platelet-derived exosome therapies and underscores the urgent need for standardization to optimize therapeutic outcomes in PRP–MSC-based regenerative strategies.
Impact Statement
This review underscores the critical interplay between platelets and mesenchymal stem cells (MSCs) in tendon–bone healing, highlighting their synergistic roles in promoting osteogenesis, chondrogenesis, angiogenesis, and anti-inflammatory responses. By elucidating the molecular and cellular mechanisms governing these interactions, this work sheds light on potential regenerative therapies that leverage platelet-derived growth factors and MSCs. Such insights aim to optimize healing outcomes for tendon–bone injuries, providing a foundation for innovative clinical strategies that restore the unique and complex structure of the tendon–bone interface.
Keywords
Introduction
Tendons are essential for transmitting muscle contractions into movement by connecting muscles to bones. Tendon injuries, therefore, can severely impair function, causing pain and disability. 1 With an estimated global incidence of 80 to 90 ruptures per 100,000 people annually, amounting to 6–7 million cases worldwide, tendon injuries represent a significant health burden. 1 The tendon–bone insertion (TBI) features a complex hierarchical structure—from tendon to uncalcified cartilage, calcified cartilage, and bone—crucial for distributing mechanical stress. 2 Current treatments for TBI injuries primarily focus on tendon and ligament repair, which requires secure grafting into the bone tunnel. The success of these procedures depends heavily on the proper integration of the grafted tendon. 3
However, despite advances in autologous ligament repair, key challenges remain. A lack of gradual transition at the tendon–bone interface (TBI) often leads to stress concentration, impeding the healing process.4,5
Mesenchymal stem cells (MSCs) are extensively utilized as progenitor cells in regenerative medicine due to their capacity for self-renewal, pluripotent differentiation, immunomodulation, and promotion of angiogenesis. 6 During TBI healing, platelet activation and the subsequent release of bioactive growth factors are critical. Recently, platelet-derived products, such as platelet-derived exosomes and platelet-rich plasma (PRP), have been incorporated into regenerative medicine therapies due to their abundance of growth factors. However, the underlying molecular mechanisms governing the interaction between platelets and MSCs are not fully elucidated. 7 In this review, the immunoregulatory and angiogenic functions of platelets, the multidirectional differentiation, and immunoregulatory and angiogenic functions of MSCs in tendon–bone healing are summarized, and the interaction between platelets and MSCs is discussed (Fig. 1).
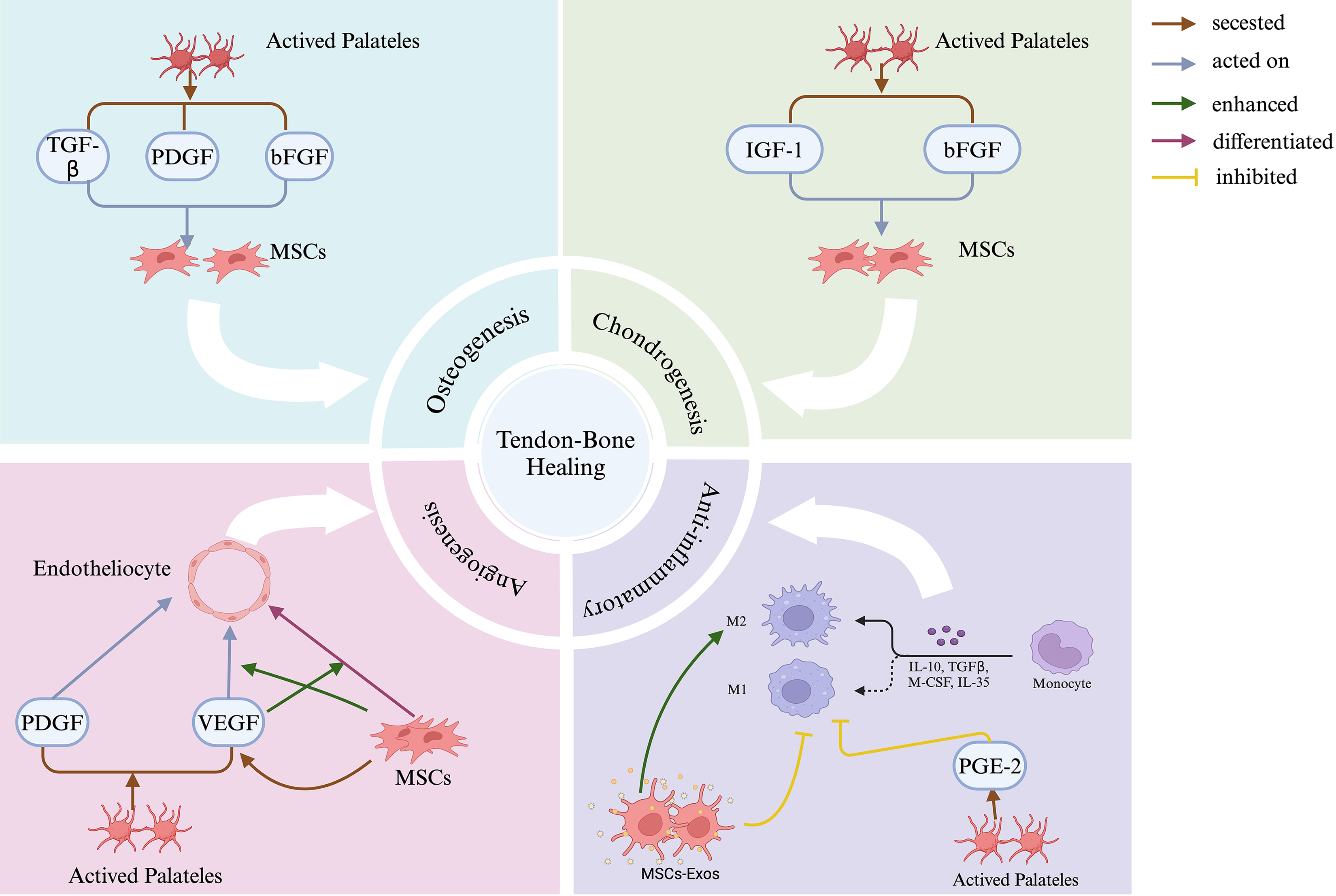
Platelets and MSCs work synergistically to promote tendon–bone healing through four critical pathways: osteogenesis, chondrogenesis, anti-inflammation, and angiogenesis. Activated platelets secrete key growth factors, including PDGF, TGF-β, bFGF, and IGF-1, which act on MSCs to stimulate their proliferation, differentiation, and functional activities across these pathways. In osteogenesis and chondrogenesis, MSCs differentiate into osteogenic and chondrogenic lineages under the influence of specific growth factors. During angiogenesis, PDGF and VEGF secreted by platelets enhance MSCs activity, promoting neovascularization. For anti-inflammatory effects, MSCs-Exos and mediators such as PGE-2 shift macrophages from an M1 to an M2 phenotype, fostering a regenerative, anti-inflammatory environment. Together, these processes contribute to the complex healing and integration of the TBI. MSCs, mesenchymal stem cells. PDGF, platelet-derived growth factor. TGF-β, transforming growth factor-β. bFGF, basic fibroblast growth factor. IGF-1, insulin-like growth factor-1. VEGF, vascular endothelial growth factor. MSCs-Exos, MSCs-derived exosomes. PGE-2, prostaglandin E2. TBI, tendon–bone interface. Created in BioRender. Wu, Y. (2024) https://BioRender.com/t80j036.
Tendon–Bone Interface
Unique features of the tendon–bone interface structure
The TBI is a common structure in the human body, composed of four distinct anatomical regions: tendon, uncalcified fibrocartilage (UFC), calcified fibrocartilage (CFC), and bone2,8 (Fig. 2). These regions exhibit a continuous gradient of cell phenotypes without distinct boundaries, such as the transition from tenocytes in the unmineralized region to fibro chondrocytes in the mineralized region, and from fibro chondrocytes in the un-mineralized fibrocartilage to hypertrophic chondrocytes in the mineralized fibrocartilage. Simultaneously, the mineral content of the extracellular matrix (ECM) gradually increases. In the hard tissue region, the main cell types are osteocytes, osteoblasts, and osteoclasts, and the ECM is characterized by relatively disordered collagen fibers and high mineral content. It is currently believed that this transitional structure aids in transferring loads between the relatively soft, elastic tendons and the rigid, nonelastic bones. Additionally, the gradient change in mineral content is considered the key mechanism for stress dissipation at the mature fibrocartilage interface. 8 In different regions of the TBI, various types of collagens are distributed and function differently. For example, type I collagen is predominantly found in tendon and bone tissue and serves as the primary structural protein, while types II, IX, and XI collagen are primarily located in the UFC and CFC regions, contributing to the biological functions of cartilage tissue. Type X collagen is mainly found in the CFC region, synthesized by hypertrophic chondrocytes, and plays a role in the mineralization process. In addition to collagen, the TBI contains several nonfibrillar collagens and other proteins, such as types VI and XIV collagen, which are generally involved in regulating the biomechanical properties of tissues. 2 These features highlight that the tendon–ligament–bone interface is a complex and functionally diverse biomechanical zone that facilitates a smooth transition from soft tissue to hard bone tissue, maintaining functionality through its unique cellular composition, ECM, and adaptive mechanical properties. 9
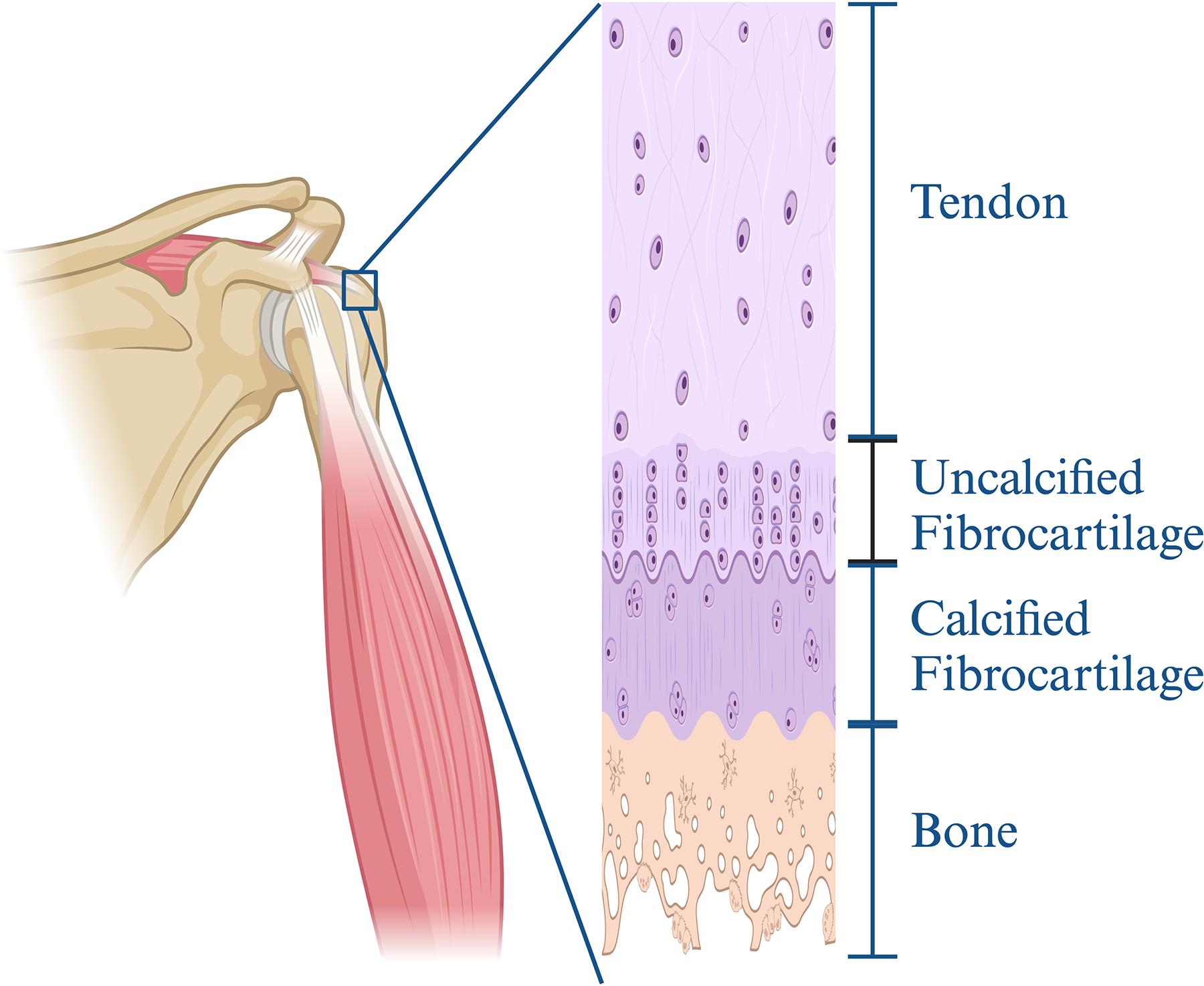
The four-layer structure of the TBI. Created in BioRender. Wu, Y. (2024) https://BioRender.com/k15w133.
Pathophysiology after tendon attachment injury
Injury to the TBI can compromise tissue integrity and function, leading to symptoms such as pain and stiffness. Using rotator cuff and Achilles tendon injuries as examples, tendon damage in the early stages can trigger an inflammatory response characterized by the infiltration of inflammatory cells, such as macrophages and neutrophils. These cells release cytokines and growth factors to initiate the healing process. However, if the inflammation is excessive or prolonged, it may cause further tissue damage. Injured tendon cells (tenocytes) may become elongated and thin, producing less ECM, which disrupts the collagen fiber structure. This results in ECM disorganization and weakens mechanical properties of the tendon.10,11 In the chronic course of the disease, chronic tendinopathy leads to the breakdown and disorganization of collagen fibers over time. Histologically, tendinopathy manifests as increased inflammatory tissue, decreased vascular supply, and changes in collagen structure; however, these changes may not cause corresponding symptoms.
10
After the injury to the tendon–bone junction, the healing process can generally be divided into four main stages:
Inflammatory phase: Following the injury, blood vessels rupture, leading to local bleeding. Platelets release and deposit fibrin and fibronectin at the injury site to form an initial blood clot. These platelets also release growth factors, such as insulin-like growth factor-1 (IGF-1), platelet-derived growth factor (PDGF), and transforming growth factor-β (TGF-β), to initiate the healing process.
12
Simultaneously, inflammatory cells are recruited to the injury site, and the healing process begins with the infiltration of a large number of inflammatory cells and the release of cytokines. Researchers also observed the formation of fibrovascular tissue at this stage, laying the foundation for subsequent tissue regeneration.
13
Proliferation phase: During this phase, stem cells proliferate and differentiate, forming new ECM. Fibrovascular tissue gradually increases, and new blood vessels grow, providing nutritional support for tissue regeneration. Fibrocartilage begins to form between tendons and bones, which enhances the mechanical strength of the tendon–bone connection.13,14 Matrix synthesis phase: The newly formed ECM is gradually synthesized and modified. In response to TGF-β, recruited fibroblasts begin synthesizing collagen and other matrix components. The matrix reorganizes into dense connective tissue,
15
and fibrocartilage gradually transforms into mineralized cartilage, eventually forming bone tissue. New bone gradually grows during this stage and begins to form structural support at the junction of tendon and bone, enhancing the mechanical strength of the interface.
14
Matrix remodeling phase: The newly formed bone and tendon matrix mature gradually, and type III collagen in the scar tissue is replaced by type I collagen, forming a more stable and organized tissue structure.
12
However, excessive and prolonged inflammation may result in excessive scar tissue with lower mechanical properties than the normal four-layer transition structure, leading to suboptimal tendon–bone healing.
15
In summary, tendon–bone healing is a complex and dynamic process, with each phase characterized by specific biological activities and functions. Understanding the timing and sequence of these phases is critical for developing effective treatments.
The Key to Tendon–Bone Healing
Based on the fundamental process of tendon–bone healing, we believe that to reconstruct the natural structure of the TBI, interventions should focus on four key areas: osteogenesis, chondrogenesis, anti-inflammation, and angiogenesis.
The Role of Platelets in Tendon–Bone Healing
Platelets play a crucial role in the early response to tissue injury. Growth factors released by platelets, such as PDGF, TGF-β, and vascular endothelial growth factor (VEGF), not only promote angiogenesis and tissue regeneration but also recruit and activate MSCs to participate in the repair of the injured site.
Platelets promote tendon–bone healing by affecting inflammation
The inflammatory response plays a crucial role in tendon–bone healing. Inflammatory cells can phagocytose and remove necrotic tissue and cells. The inflammatory factors they release can also recruit MSCs, promoting their involvement in the healing process. Therefore, platelets can facilitate tendon–bone healing by modulating the inflammatory response (Fig. 3).
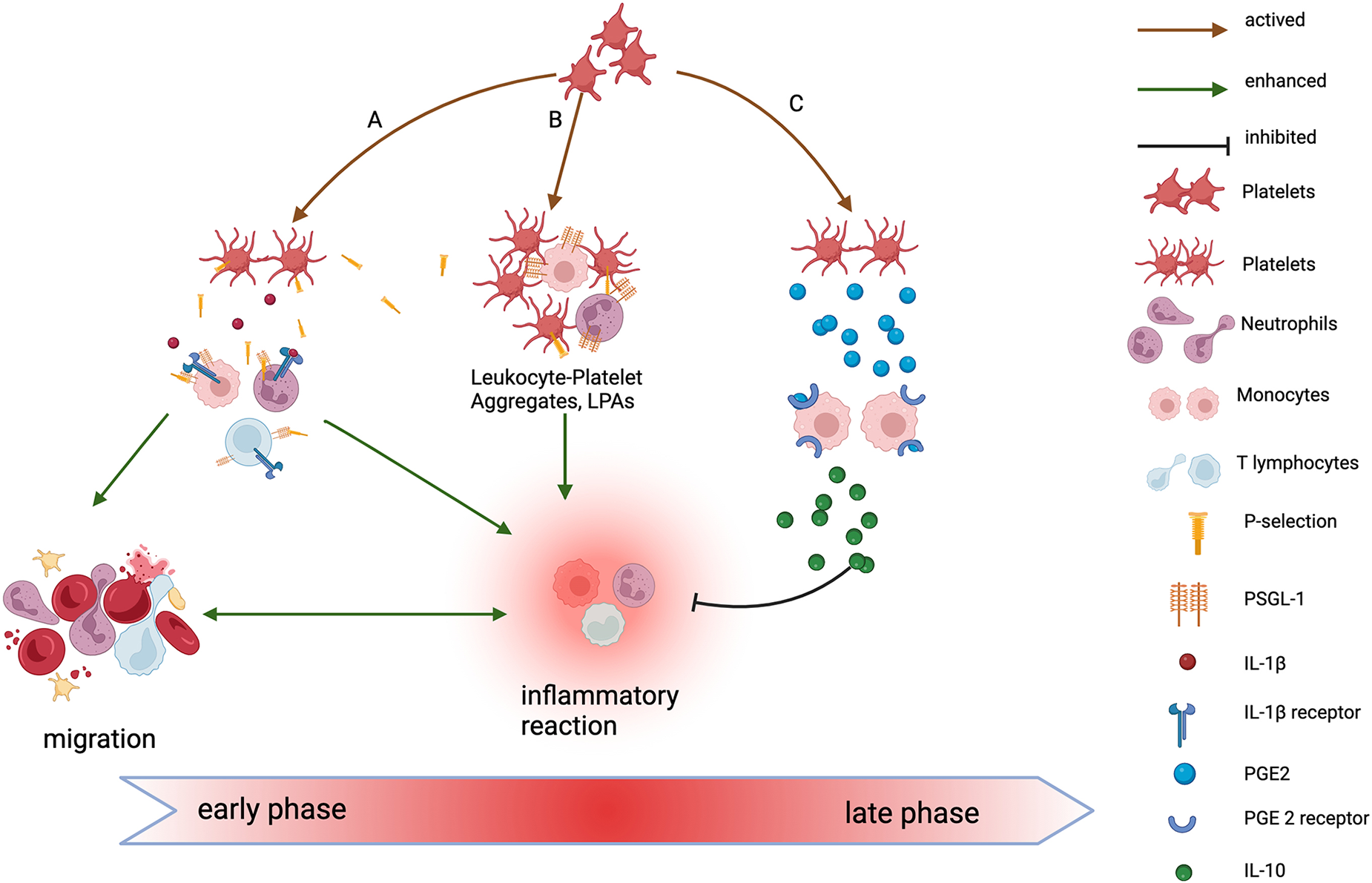
A In the early phase of injury, after platelet activation, P-selectin and IL-1β secreted by platelets bind to receptors on the surface of inflammatory cells, activating them and triggering an inflammatory response. These activated inflammatory cells then migrate to the injury site, releasing inflammatory factors, which further amplify the inflammatory reaction. B In the early stage of injury, activated platelets form LPAs by interacting with leukocytes via P-selectin and its ligand, PSGL-1, further enhancing the inflammatory response. C In the late stage of injury, PGE2 secreted by activated platelets binds to receptors on macrophages, inducing them to secrete anti-inflammatory factors such as IL-10. This promotes the shift of macrophages to a regenerative, anti-inflammatory phenotype, helping to resolve inflammation. IL-1β, interleukin-1 β. LPAs, leukocyte-platelet aggregates. PSGL-1, P-selection glycoprotein ligand-1. PGE2, prostaglandin E2. IL-10, interleukin-10. Created in BioRender. Wu, Y. (2024) https://BioRender.com/d37n737.
Noval Rivas et al. 16 demonstrated that platelets can interact with both immune and nonimmune cells to regulate inflammation. Platelets recruit and activate immune cells, enhancing inflammatory responses by releasing various chemical signals and cytokines, such as platelet factor 4 (PF4), interleukin-1β (IL-1β), and C-X-C motif chemokine ligand 4 (CXCL4). In addition, activated platelets form aggregates with white blood cells, such as monocytes and neutrophils, enhancing the functions of these cells and promoting thrombosis and inflammatory responses. Research by Thomas Gremmel and colleagues 17 showed that P-selectin secreted by platelets can bind to P-selectin ligand-1 (PSGL-1) on white blood cells, promoting the activation, adhesion, and migration of these cells, thereby amplifying the inflammatory response. Furthermore, research by Su et al. 18 demonstrated that PRP significantly promotes macrophage M2 polarization and fibroblast migration. Macrophage M2 polarization enhances the secretion of TGF-β, which promotes the chondrogenic differentiation of MSCs, thereby contributing to tendon–bone healing. Research by Ludwig et al. 19 demonstrated that platelets regulate inflammatory responses by releasing prostaglandin E2 (PGE2) and TGF-β. PGE2 increases the production of the anti-inflammatory cytokine interleukin-10 (IL-10) by binding to Prostaglandin E2 receptor 2 (EP2) and Prostaglandin E2 receptor 4 (EP4) on the surface of macrophages and monocytes, thereby alleviating the inflammatory response. Meanwhile, TGF-β promotes the differentiation of regulatory T cells (Tregs), which guide macrophages toward anti-inflammatory M2 polarization, promoting tissue repair and reducing inflammation. Excessive inflammatory responses can lead to scar tissue formation, but platelets help mitigate scar formation at the TBI by regulating the resolution of inflammation.
Platelets promote tendon–bone healing by affecting angiogenesis
Although PDGF and VEGF released by platelets do not directly promote tendon–bone healing, they facilitate angiogenesis and increase local blood supply, thereby creating a favorable microenvironment for tendon–bone healing.
According to literature reports, 20 the PDGF family consists of four members: PDGF-A, PDGF-B, PDGF-C, and PDGF-D. Among them, PDGF-C plays a particularly important role in the angiogenesis process. PDGF-C stimulates the migration and proliferation of endothelial cells and vascular smooth muscle cells by binding to the α-type PDGF receptor (PDGFR-α), thereby promoting the formation of new blood vessels. PDGF-D primarily activates vascular smooth muscle cells by binding to β-type PDGF receptor (PDGFR-β), stimulating their proliferation and migration, and thereby supporting new blood vessel formation. Additionally, both PDGF-C and PDGF-D can activate matrix metalloproteinase-9, promoting ECM degradation and creating space for the formation of new blood vessels.
According to literature reports, 21 damaged tissue cells and activated platelets in a hypoxic environment can release VEGF, which binds to VEGF receptors (primarily VEGFR-1 and VEGFR-2) on the surface of vascular endothelial cells. This binding causes receptor dimerization and phosphorylation, thereby activating downstream signaling pathways. VEGF also promotes the proliferation and migration of endothelial cells through the Ras-MAPK, PI3K-Akt, and PLCγ pathways, ultimately facilitating new blood vessel formation. The study by Levoux et al. 22 demonstrated that platelets can also enhance the angiogenic function of MSCs through mitochondrial transfer and gene encoding. In the experiment, the researchers used Mito Tracker Green, a mitochondrial fluorescent dye, to label platelet mitochondria and observed that platelets released mitochondria into the extracellular space via exocytosis. The released mitochondria could be in free form or encapsulated in microparticles. MSCs absorb these mitochondria through dynamin-dependent, clathrin-mediated endocytosis. The mitochondria within the endocytic vesicles are then transported into the intracellular environment of MSCs and integrated into the mitochondrial network of the MSCs. The platelet-derived mitochondria maintain their respiratory function, enhance the metabolic activity of MSCs, promote the activation of the de novo fatty acid synthesis pathway, and increase adenosine triphosphate production. As a result of this activation, MSCs secrete more angiogenic factors, such as VEGF, which promotes the formation of new blood vessels at the site of injury, improves blood and oxygen supply to the injured tissue, and facilitates tendon–bone healing.
In addition, PF4 released by platelets exerts an antiangiogenic effect and regulates the process and distribution of angiogenesis around damaged tissues.
Role of Stem Cells in Tendon–Bone Healing
In recent years, as researchers explore ways to improve tendon–bone healing, MSCs have been utilized in tissue engineering therapies, yielding promising therapeutic effects. While most MSCs are derived from bone marrow, due to the invasive nature of bone marrow stem cell extraction, researchers have investigated the role of MSCs from alternative sources in tendon–bone healing (Table 1). Despite originating from different sources, these cells typically express positive markers for CD29, CD44, CD73, CD90, and CD105, with low or no expression of CD11b, CD14, CD34, and CD45.23,24 Additionally, some studies have found that MSCs can be extracted from fracture hematomas.25,26
Application of Various Stem Cells in Tendon–Bone Healing

First, MSCs have multidirectional differentiation potential. Under various physiological stimuli, they can differentiate into adipocytes, osteoblasts, tenoblasts, chondrocytes, and other cell types (Fig. 4).
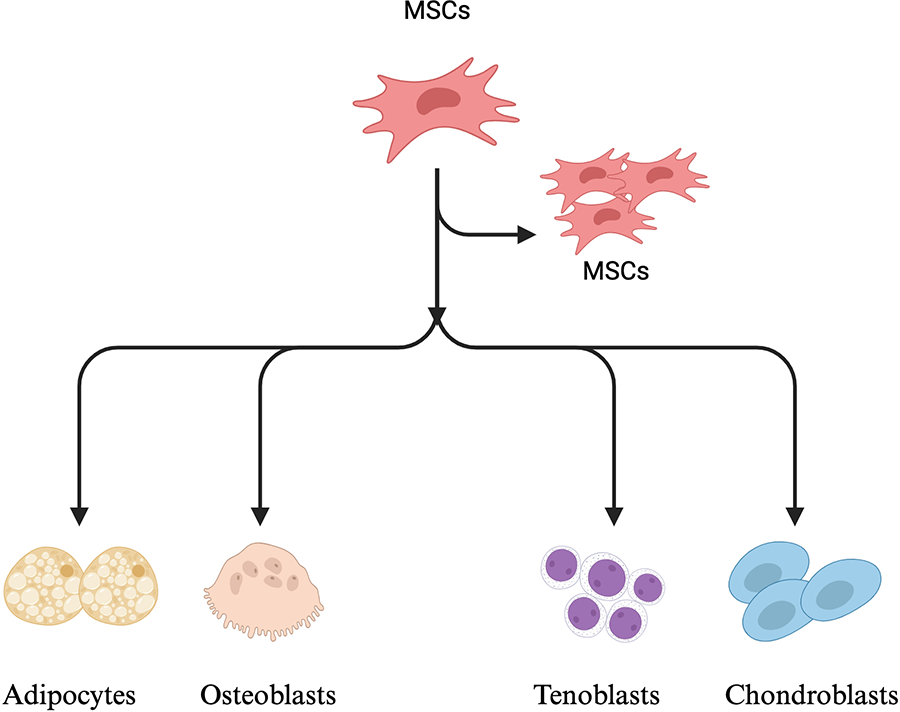
MSCs have proliferation and multidirectional differentiation functions. Created in BioRender. Wu, Y. (2024) https://BioRender.com/h40l181.
Rothrauff et al. 27 encapsulated adipose-derived mesenchymal stem cells (ADSCs) in fibrin or gelatin methacryloyl and used them to repair acute and chronic rotator cuff tears in rats. They observed that the use of ADSCs significantly improved the histological and structural properties of the tendon insertion. Lee Song et al. 28 demonstrated that TGF-β signaling regulates the proliferation of GLI family zinc finger 1-positive (Gli1+) stem cells through the classical Smad-dependent pathway and the nonclassical MAPK/JNK/c-Jun pathway. TGF-β binds to a distal enhancer downstream of the Scx gene via Smad4, thereby upregulating Scx expression, promoting the differentiation of Gli1+ stem cells into tendon cells or related tissue types, and facilitating tendon–bone healing. He et al. 29 used demineralized cortical bone to prepare a biomimetic scaffold coated with an ECM containing tendon–derived stem cells. This scaffold was applied in rabbit rotator cuff injury repair and achieved positive results in promoting tendon–bone healing. The study suggests that a composite scaffold combining hierarchically demineralized cortical bone with stem cell-derived ECM can recruit more mesenchymal cells and promote the formation of bone and fibrocartilage, thus enhancing the healing outcome after rotator cuff repair. Due to the multipotential differentiation capacity of MSCs, tendon–bone healing can be improved by promoting both osteogenesis and cartilage formation.
Second, MSCs have immunomodulatory functions. Li et al. 30 found that exosomes (MSCs-Exos) derived from MSCs before osteogenic differentiation exhibit immunomodulatory properties. These MSCs-Exos can reduce the expression of proinflammatory factors (such as iNOS) and increase the expression of anti-inflammatory factors (such as CD206), promoting the transformation of macrophages from the M1 to the M2 phenotype. M2 macrophages are anti-inflammatory and secrete anti-inflammatory cytokines, such as Arg-1, CD206, and IL-4, which facilitate tissue repair and regeneration. Research by Liu et al. 31 showed that bone marrow mesenchymal stem cell exosomes (BMSCs-EVs) significantly increased the expression of M2 macrophage marker genes (such as IL-10 and Arg1), while reducing the expression of M1 macrophage marker genes (such as IL-6). During tendon–bone healing, the inflammatory response and polarization of inflammatory cells play a critical role. The early inflammatory response is beneficial for wound healing, but excessive inflammation can lead to tissue fibrosis and scar formation, which hinders the restoration of the natural TBI. MSCs can promote tendon–bone healing by influencing macrophage polarization.
In addition, MSCs can regulate tendon–bone healing by influencing angiogenesis. The study by Lan et al. 32 showed that MSCs release various bioactive factors, such as VEGF, fibroblast growth factor (FGF), and stromal cell-derived factor-1 (SDF-1), which promote blood vessel formation and regeneration by enhancing endothelial cell activity. Moreover, MSCs also release microvesicles and exosomes containing these bioactive molecules. These exosomes can indirectly promote angiogenesis and regeneration by upregulating VEGF expression in macrophages. Qiu et al. 33 demonstrated that miR-221-3p in BMSCs-EVs promotes angiogenesis by targeting Fork head Box P1, leading to the downregulation of Sprouty RTK Signaling Antagonist 1 expression. The in vitro study by Liu et al. 31 further confirmed the angiogenic potential of MSCs Exos, showing that they primarily affect the expression of several classical angiogenic genes in endothelial cells, such as VEGF-A and PDGF-B. During the tendon–bone healing process, the generation of new blood vessels is crucial because it provides the necessary blood and oxygen supply to support the formation of new tissue.
Mutual Influence between Platelets and MSCs
Platelets contain a variety of cytokines and growth factors. These bioactive substances can influence the proliferation, migration, and differentiation of MSCs, thereby playing a crucial role in tissue healing and regeneration (Table 2).
Cytokines/Growth Factors Derived from Various Platelets

PDGF was discovered in 1974. It is a highly conserved polypeptide dimer released by platelets and connected by disulfide bonds. After PDGF binds to its receptor, the receptor dimerizes and autophosphorylates, forming an activated tyrosine kinase receptor complex. The activated receptor regulates various biological processes, such as cell proliferation, migration, differentiation, and survival, by phosphorylating downstream signaling molecules. 20 Numerous studies have shown that PDGF promotes the proliferation, migration, and osteogenic differentiation of MSCs by acting on the PDGFR-α on the surface of MSCs.34–36 Therefore, during tendon–bone healing, PDGF can facilitate the restoration of the natural TBI structure by promoting osteogenesis.
TGF-β is a homodimeric molecule composed of two subunits connected by disulfide bonds and includes the subtypes TGF-β1, TGF-β2, and TGF-β3, with TGF-β1 being the most dominantly expressed. 37 Some studies have reported that N-cadherin, PI3K/Akt, ERK1/2, FAK, and p38 signaling pathways are involved in TGF-β1-induced migration of human BMSCs. 38 Shariat et al. 39 demonstrated that the effect of TGF-β on osteogenesis is dose-dependent: at low doses, TGF-β promotes osteoblast proliferation and differentiation, whereas at high concentrations, it inhibits osteogenesis. Sima et al. 40 showed that under mechanical load, TGF-β is essential for MSCs to form new cartilage. Xiao et al. 41 conducted a more in-depth study on the mechanism of TGF-β in promoting chondrogenesis. During the healing process following tendon attachment injury, the concentration of active TGF-β increased, peaking after 2 weeks before gradually returning to baseline levels. Prrx1+ cells, a subtype of MSCs that play a key role in the development of bone, cartilage, and tendon, migrate from the surrounding tissue to the rotator cuff injury site under the influence of TGF-β1, where they participate in the healing process of the tendon attachment and differentiate into chondrocytes and osteoblasts.
IGF-1 is another growth factor released by platelets that promotes the migration and osteogenic and chondrogenic differentiation of MSCs. Jingjun et al. 42 demonstrated that IGF-1 induces the osteogenic differentiation of rat MSCs by stimulating SOX4 through the MAPK/ERK pathway. Li et al. 43 further explored its potential mechanism and found that IGF-1 enhances the migration response of MSCs via PI3K/Akt-dependent CXCR4 chemokine receptor signaling. Guan et al. 44 discovered that increased levels of IGF-1 and hepatocyte growth factor upregulate the expression of osteoblast-related genes and chondrogenesis-related genes and proteins in BMSCs. Additionally, the study by Dong Mao et al. 45 showed that IGF-1 mediates the progression of traumatic heterotopic ossification through the PI3K/Akt/mTOR signaling pathway and inhibits the IGF-1 signaling cascade reducing heterotopic ossification formation, further confirming IGF-1’s role in the osteogenic differentiation of MSCs.
FGF is an important family of growth factors involved in various biological functions. Of particular interest is the basic fibroblast growth factor (bFGF), which plays a key role in the regeneration of tendons, ligaments, and bone interfaces. 9 Research by Goshima et al. 46 demonstrated that bFGF exerts its effects by upregulating the gene expression of the chemokine CXCL6 in MSCs. The bFGF promotes the expression of CXCL6, which, after binding to the CXCR2 receptor, activates downstream ERK and Akt signaling pathways, enhancing cell migration, proliferation, and chondrogenic differentiation. Research by Park et al. 47 found that bFGF shortens the doubling time of MSCs by activating downstream signaling pathways (such as ERK, Akt), driving cells from the G1 phase to the S phase, thereby accelerating the cell cycle process. Additionally, bFGF promotes MSCs to secrete extracellular vesicles containing anti-inflammatory factors such as TNF-α and IL-1β, which inhibit the inflammatory response at the injury site and promote healing.
VEGF is a signaling protein that can act on both endothelial cells and MSCs. Its primary function is to promote angiogenesis and provide nutrients for cell proliferation and tissue repair, thereby indirectly influencing tendon–bone healing. Subbarayan et al. 48 demonstrated that VEGF can induce MSCs to differentiate into endothelial cells, thereby promoting angiogenesis. They cocultured MSCs with VEGF and, using flow cytometry, detected that the cells began to express endothelial cell-specific markers, such as VEGF-A and CD31. Yang et al. 21 also showed that VEGF promotes MSC proliferation by activating the MAPK/ERK pathway and supports MSC survival and the expression of osteogenic-related genes by activating the PI3K/Akt pathway, thereby promoting the differentiation of MSCs into osteoblasts. Liu et al. 49 further demonstrated that VEGF promotes the formation of new blood vessels. These new blood vessels can provide a better microenvironment for MSC-derived osteoblasts, thereby promoting bone formation.
In addition, research by Chen et al. 50 showed that platelet-derived exosomes (PL-Exos) inhibit lipid peroxidation by activating AMPK phosphorylation and the downstream Nrf2/GPX4 signaling pathway, thereby reducing tendon stem/progenitor cell (TSPC) ferroptosis and TSPC senescence, while promoting tendon regeneration. In a rat model, it was verified that PL-Exos improved tendon–bone healing performance and mechanical strength. Molin Li et al. 51 encapsulated PRP-derived exosomes (PRP-Exos) in fibrin gel and applied it to the repair of acute rotator cuff injury in rabbits. The results showed that PRP-Exos reduced muscle atrophy and fat infiltration, promoted collagen fiber regeneration, reduced fibrosis, and enhanced new bone formation during tendon–bone healing. Additionally, PRP-Exos significantly promoted the proliferation and migration of TSPCs and induced their differentiation into tendon and cartilage phenotypes.
Advances and Challenges in PRP–MSCs Therapy
In recent years, tissue engineering strategies have increasingly emphasized the combined application of PRP and MSCs to promote tendon–bone healing. PRP facilitates tissue repair by releasing a variety of cytokines and growth factors, such as VEGF and FGF, which support cell proliferation, migration, differentiation, and angiogenesis. Meanwhile, MSCs contribute to tissue regeneration through their multilineage differentiation potential and paracrine signaling capabilities.
Multiple preclinical studies have confirmed the beneficial effects of PRP–MSCs combination therapy in tendon–bone regeneration.24,52,53 For example, Han et al. 52 reported that PRP significantly upregulated osteogenic and chondrogenic gene expression in MSCs, resulting in improved tendon–bone integration in a rat supraspinatus repair model. Similarly, Teng et al. 53 demonstrated the positive effects of PRP combined with BMSCs in enhancing tendon–bone healing. Despite these promising findings, the synergistic effects of this combined therapy remain controversial. Oda et al. 54 found that PRP combined with ADSCs, encapsulated in collagen-rich hydrogels, improved biomechanical strength in a chronic rotator cuff injury model. However, no clear synergy or dose-dependent effect between PRP and ADSCs was observed. These inconsistencies may be attributed to differences in PRP preparation methods, which can significantly impact the concentration and bioactivity of its growth factors. 55 Moreover, the heterogeneity of MSCs sources and the lack of standardized dosing protocols further hinder reproducibility. In addition, PRP-Exos have emerged as a recent research hotspot in the field of regenerative medicine.50,56 Compared with PRP, PRP-Exos exhibit lower immunogenicity, greater structural stability, and a robust capacity to deliver a diverse range of bioactive molecules. 57 Despite these advantages, the lack of standardized methods for quantification and quality control presents a significant barrier to their clinical translation.
Regardless of whether PRP, MSCs, or exosomes are used, the long-term safety, immunogenicity, and efficacy of these combination therapies must be validated through large-scale, longitudinal clinical trials before they can be widely implemented in clinical practice. Further investigation into the mechanisms of PRP–MSC synergy will not only improve the quality and efficiency of tendon–bone healing but may also provide new therapeutic insights for regenerative strategies in musculoskeletal and soft tissue systems.
Conclusion and Future Outlook
The TBI is a structurally complex and biomechanically critical zone that presents significant challenges in postinjury healing. Current evidence highlights that both platelets and MSCs play synergistic roles in modulating the biological processes essential for tendon–bone regeneration, including osteogenesis, chondrogenesis, angiogenesis, and immune regulation. While MSCs serve as potent seed cells with multilineage differentiation and paracrine functions, platelets and their derivatives provide an abundant source of growth factors that stimulate MSC activity and tissue repair. Emerging tissue engineering strategies have increasingly explored the combined use of PRP, MSCs, demonstrating enhanced biological and mechanical outcomes in preclinical models. However, despite promising results, several translational challenges persist, including variability in PRP preparation, heterogeneity of MSC sources, and the lack of standardized dosing and delivery protocols. Furthermore, although PRP-Exos offer lower immunogenicity and improved stability, they too face obstacles related to quality control and clinical scalability.
Future research should prioritize the standardization of biological preparations, elucidate the underlying mechanisms of PRP–MSC synergy, and rigorous evaluation through large-scale clinical trials. Advancing these integrated strategies will be essential not only for restoring the native structure and function of the TBI but also for informing broader applications across the field of regenerative medicine.
Authors’ Contributions
Y.W.: Conceptualization (lead); writing—original draft (lead); validation (lead); formal analysis (lead); and writing—review editing (equal). X.W.: Writing—review editing (equal) and validation (supporting). D.Z.: Writing—review editing (equal). R. L., X.Z., and X.L.: Conceptualization (supporting); writing—original draft (supporting); and writing—review and editing (equal).
Footnotes
Funding Information
This research was supported by Yunnan Revitalization Talent Support Program XDYC-ONRC-2022-0318 and the Kunming Medical University Graduate Innovation Fund Project 2024S283.
Disclosure Statement
No competing financial interests exist.