
Abstract
In this study, Fourier transform infrared spectroscopy (FTIR) was used to evaluate the effects of a gamma irradiation process on AlloDerm, an implantable tissue device derived from human cadaver skin. Inspection of protein amide bands of dried tissue matrix revealed little difference in overall protein secondary structure between gamma-treated and control tissue matrices, although effects of gamma irradiation became apparent after rehydration. The temperature at which protein denaturation began decreased significantly, from 62.4°C ± 1.0°C in the nonsterilized control tissue matrix to 53.4°C ± 1.2°C, 48.7°C ± 0.3°C, and 46.7°C ± 0.1°C for 5-, 14- and 20-kGy gamma-treated AlloDerm samples, respectively (p < 0.05). Differences were particularly significant in the details of the tissue matrix denaturation profile, which consisted of multiple transitions. The solvent accessibility of tissue matrix was studied by following the rate of protein proton exchange in heavy water (D2O), which was evident from the appearance of the amide-II' band as a function of time to D2O exposure. In control tissue matrix, a rapid hydrogen/deuterium exchange was observed, with 50% exchange in 30 min, whereas the gamma-treated tissue matrix exhibited a much slower exchange, reaching the 50% exchange level after more than 2 h. The data indicate significant alterations of extracellular milieu in the tissue matrix after the gamma irradiation process. This study shows that FTIR is a valuable tool for studying protein stability and interactions in complex biological scaffolds such as extracellular tissue matrix.
Introduction
In this study, FTIR was used to study the effects of gamma irradiation on the protein structure and stability of the AlloDerm. Proteins can be detected using Fourier transform infrared spectroscopy (FTIR) by virtue of several characteristic absorption bands at approximately 1650 cm−1 (amide-I band), 1550 cm−1 (amide-II band), and 1250 cm−1 (amide-III band) arising from the peptide backbone.4,5 The amide-I band, which is most often used for protein analysis, is a complex band arising from the different types of protein secondary structure.4–6 The amide-I band has been extensively studied to determine the relative proportion of the different types of protein secondary structure. The disadvantage of amide-I band analysis is that the strong scissoring vibrational mode of water overlaps with the protein absorption, 6 making accurate subtraction of the water band particularly difficult for protein studies on rehydrated tissues. Recent studies have used amide-III band for protein analysis.7–9 The amide-III vibration is the weakest of the protein amide bands, but different types of secondary structure are better separated than with the amide-I and -II bands.10,11
The objective of the present study was to identify structural protein modifications using a gamma irradiation process in AlloDerm tissue matrix using FTIR. AlloDerm is a native extracellular scaffold of structural proteins and possibly signaling proteins that promote tissue reconstruction. It is derived from human cadaver skin and undergoes a series of tissue processing steps before becoming the final product. FTIR results show that gamma irradiation affects protein structure, stability, and protein–protein interactions, which lead to significant alterations of extracellular milieu in the tissue matrix.
Materials and Methods
Sample preparation
The decellularized AlloDerm tissue matrix was treated with a carbohydrate solution and freeze-dried using a patented process, which is described elsewhere.12–14 Three different lots were prepared for FTIR analysis.
Gamma irradiation of freeze-dried AlloDerm
AlloDerm tissue matrix sheets used in this study were 0.1 to 0.15 mm thick and were packed in a controlled dry environment (<2% relative humidity). Oxygen was present in the dry chamber used for packing the sample material. Tissue matrix samples were gamma irradiated at dose levels of 5, 14, and 20 kGy at room temperature (22–25°C) using a commercial facility (STERIS Isomedix, Whippany, NJ). Non-irradiated tissue samples were used as controls. Gamma-irradiated samples were stored at 4°C until used for further experimentation.
FTIR studies
To prepare samples for FTIR testing, the AlloDerm was rinsed with 0.9% saline several times to rehydrate. Tissue matrices were rehydrated for at least 1 h at 4°C. Infrared absorption measurements were carried out using a Nicolet Magna 6700 Fourier transform infrared spectrometer (Thermo-Nicolet, Madison, WI), equipped with a triglycine sulfate detector. The following acquisition parameters were used: 4 cm−1 resolution, 32 co-added interferograms, and 4000-900 cm−1 wavenumber range. Studies on dried tissue matrix were done in attenuated total reflectance (ATR) mode to avoid interference from paper backing. Studies on hydrated tissue matrix were done in a temperature-controlled transmission cell. The rehydrated samples of AlloDerm were sandwiched between two calcium fluoride windows and kept hydrated with 10 to 20 μL of 0.9% saline. The windows were sealed with grease to keep the sample hydrated, and the temperature was regulated using a temperature controller (Minco Products Inc., Minneapolis, MN). The temperature of the sample was recorded separately using a thermocouple located close to the sample. The temperature dependence of the FTIR spectra was studied by heating the sample from 20.0°C to 90.0°C at a rate of approximately 2.0°C min−1.
Protein solvent accessibility was determined by monitoring the exchange of hydrogen in the NH amide-II bond for deuterium in heavy water (D2O). This newly formed bond is identified as an amide-II' or ND bond. FTIR spectra of the tissue matrix in D2O were collected and analyzed as a function of time. The kinetics of hydrogen/deuterium (H/D) exchange of endogenous tissue matrix proteins was quantified from the increase of the amide-II' band (1500-1400 cm−1) as a function of time to D2O exposure.
Protein secondary structure analysis and heat-induced protein denaturation studies in tissue matrices were performed using temperature scanning FTIR analysis as previously described. 15 The amide-I band (1700-1600 cm−1) analysis on the dried tissue matrix and the tissue matrix in D2O was done by curve-fitting the amide-I band. The Pearson band model (a combination of Gaussian- and Lorentzian-shaped bands) was used for the curve-fitting procedure as described by Chen and Garland, 16 and the analysis was performed in PeakFit (Jandel Scientific, Corte Madera, CA). Heat denaturation studies of rehydrated tissue matrix in water were done using amide-III band analysis because the amide-I and amide-II bands were distorted in transmission mode because of sample thickness. The spectral region 1320-1200 cm−1, which contains the amide-III absorption band, was selected, and after heat-induced protein denaturation, band shape changes were monitored. The increase in area of the band between 1285 and 1240 cm−1 was used to determine the denaturation profile. Protein denaturation was visible as an abrupt increase in the area of this band.
Results
Dried tissue analysis
Figure 1 depicts ATR-FTIR spectra of control and 20-kGy gamma-treated tissue matrix in the dried state. Gamma treatment did not appear to affect the shape of the amide bands, indicating that irradiation does not measurably affect the overall protein secondary structure in dried tissue matrix. Gamma treatment, however, seems to affect the spectral region between 1200 and 1000 cm−1, which the protective molecules that have been used to stabilize the tissue matrix during freeze-drying dominate. 17 The mixture of these stabilizing molecules is termed the “excipient,” and it is composed of low-molecular-weight carbohydrates and maltodextrin. The composition of the excipient is disclosed elsewhere.12–14 The protein:excipient ratio decreases after gamma treatment, as can be seen from the decreased intensity of the bands in the fingerprint region of the excipient, which could indicate that gamma treatment causes reactions between the excipient and protein molecules.

Attenuated total reflectance Fourier transform infrared spectroscopy spectra of control and gamma-treated tissue matrices in the dried state. The spectral region labeled with “Sugars” refers to the fingerprint region of the excipient.
Protein secondary structure after rehydration
Figure 2 depicts amide-I band profiles of dried (Fig. 2A) and rehydrated (Fig. 2B) control tissue matrix in D2O. To analyze the secondary structure of proteins, the amide bands were curve fitted with the Pearson band model in PeakFit. 16 Absorption features were attributed as follows: approximately 1680 cm−1 (β-sheets); approximately 1668 cm−1 (β-turns); approximately 1658 cm−1 (α-helix); approximately 1647 cm−1 (unordered); approximately 1638 cm−1 (triple helix); approximately 1625 cm−1 (β-sheets) based on Petibois et al. 18 These band assignments are indicated in Figure 2. The band assignments take into account the shift observed in the peaks due to rehydration in D2O. The spectral amide-I features of AlloDerm are in good agreement with FTIR studies on other collagenous tissues 18 but not identical because of the shift of peaks in D2O. The dried tissue matrix contains a larger relative contribution of extended β-sheet structures. Upon rehydration, the relative contribution of extended β-sheet structures decreases, whereas random coil structures increase. It is important to note the overlap of several secondary structures in this region, which makes absolute quantitative assessment of individual secondary structures difficult.
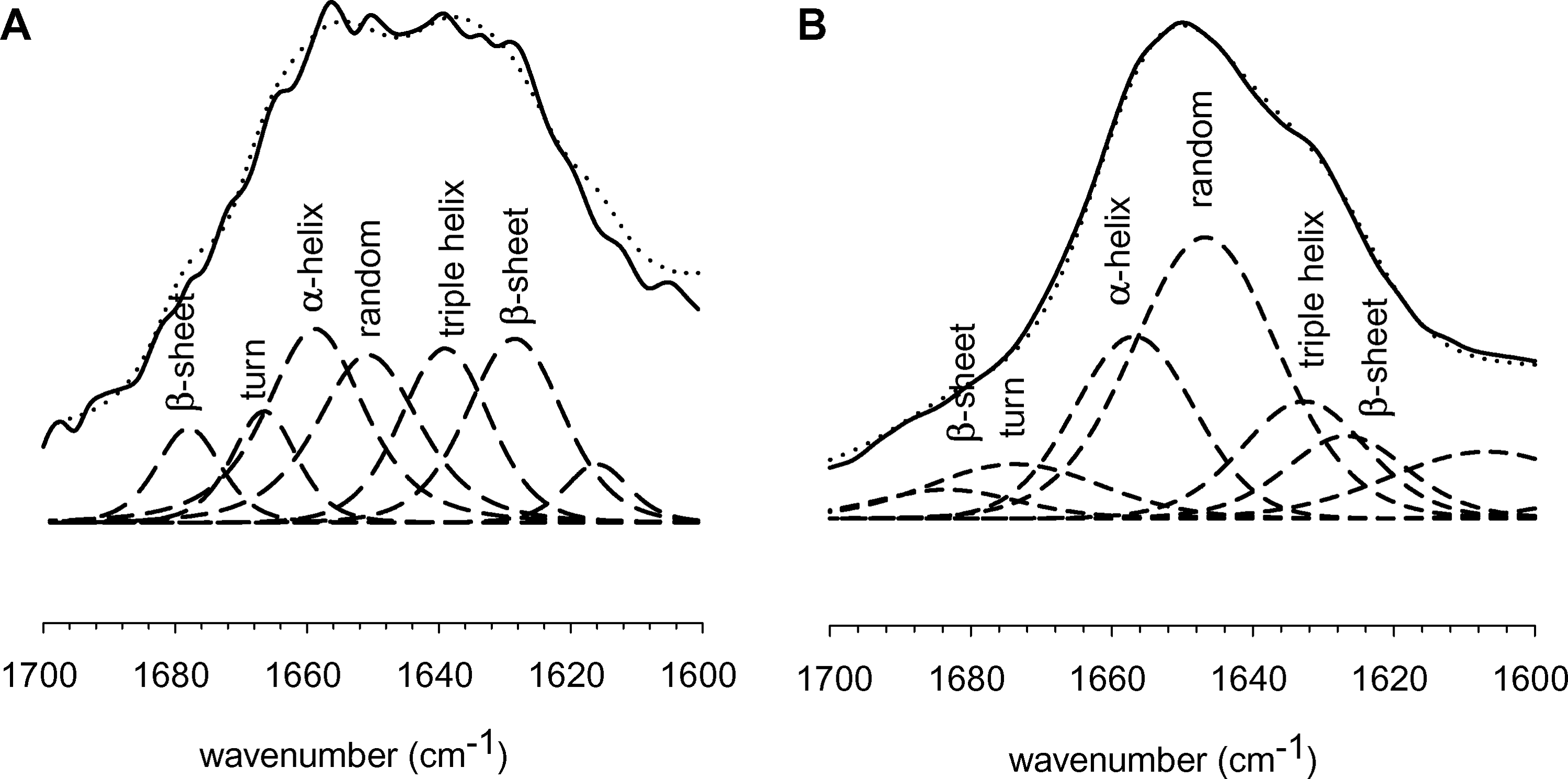
Attenuated total reflectance Fourier transform infrared spectroscopy spectra of control tissue matrix in dried (
Figure 3 shows the effect of gamma treatment on the amide-I band profile of tissue matrix at 1 h (Fig. 3A) and 4 h (Fig. 3B) after rehydration in D2O. Gamma treatment affects the overall protein secondary structure directly after rehydration, but the differences seem to diminish over time. (The difference in the band at ∼1630 cm−1 reduces from 1 to 4 h.) The results suggest that the gamma-irradiated tissue initially has a more-condensed protein matrix with more intermolecular extended β-sheet structures. This condensed protein matrix of the gamma-treated tissue slowly dissociates over time, but measurable changes in protein secondary structure remain.
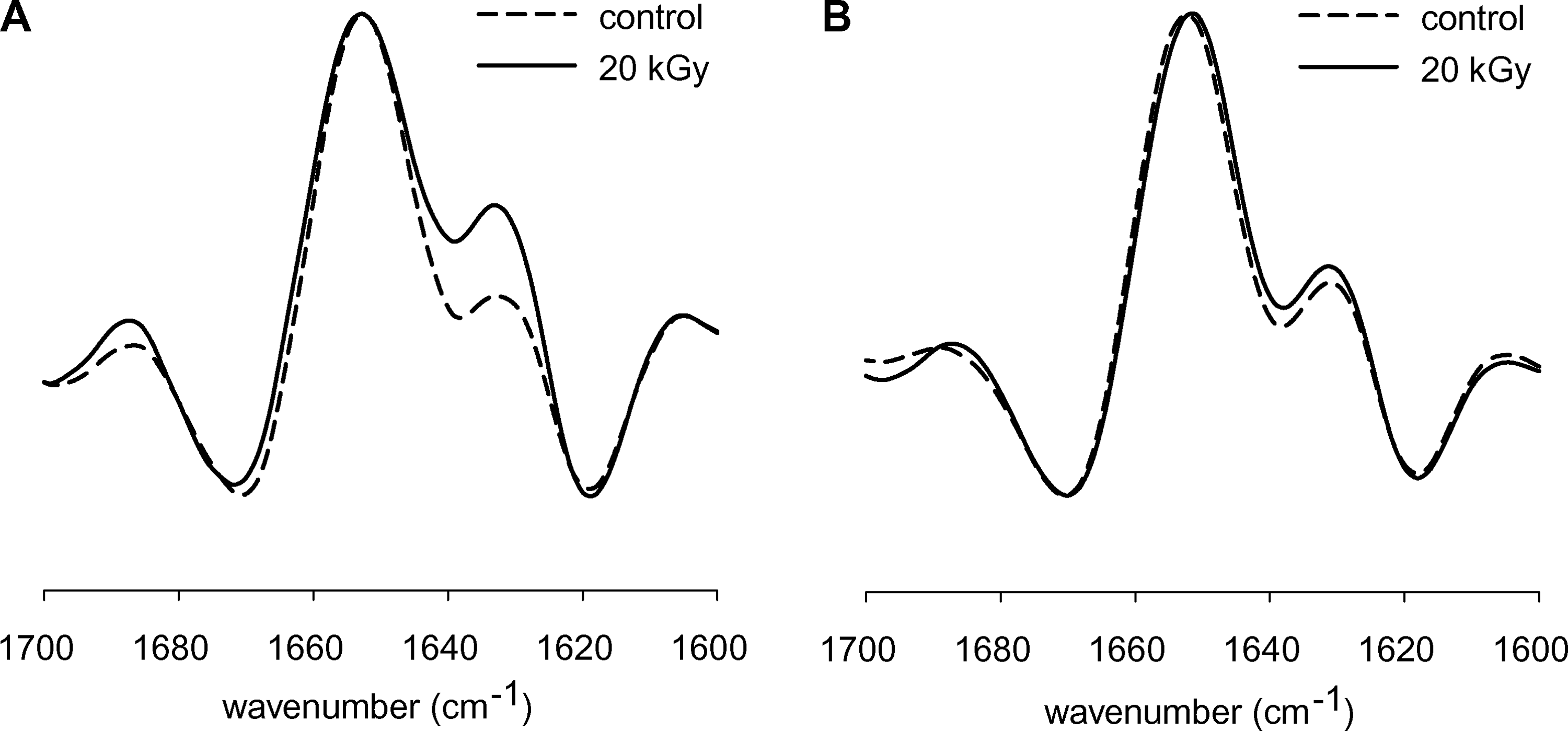
Second derivative infrared spectra of the amide-I band region of control and gamma-treated tissue matrix that were taken 1 (
Protein solvent accessibility
The differences in protein secondary structure after rehydration between gamma-treated and control tissue matrices were further quantified using H/D exchange experiments. In these experiments, the exchange of amide protons in D2O is measured as a function of time using methods described elsewhere.19,20 The hydrogen of the NH amide-II bond is exchanged for deuterium in D2O, and the resulting bond is termed amide-II' or ND. The exchange rate of individual NH groups depends on the local protein conformation. Unordered structures, for example, generally exhibit a more-rapid exchange than β-sheet structures. 19 The exchange kinetics can thus be monitored by recording the FTIR spectra as a function of time to D2O exposure (Fig. 4A). The ND band increases over time, as shown in Figure 4B. A plot of the ND band area as a function of time was constructed (Fig. 4C) and was used to determine the exchange rates (Fig. 4C). For control tissue matrix, the major protein fraction is highly solvent accessible and exhibits rapid exchange (τ1 = 30 min), whereas a smaller protein fraction exhibits slow exchange (τ2 = 212 min), indicative of a less-solvent-accessible protein fraction. Gamma-treated tissue matrix exhibits slower NH–ND exchange (τ = 103 min) than control tissue matrix (in this case, the data could be fitted using only one exponential term, with the coefficient of determination = 0.99), which indicates that gamma treatment decreases the protein solvent accessibility and suggests that it promotes condensation and collapse of the protein architecture in the tissue matrix.
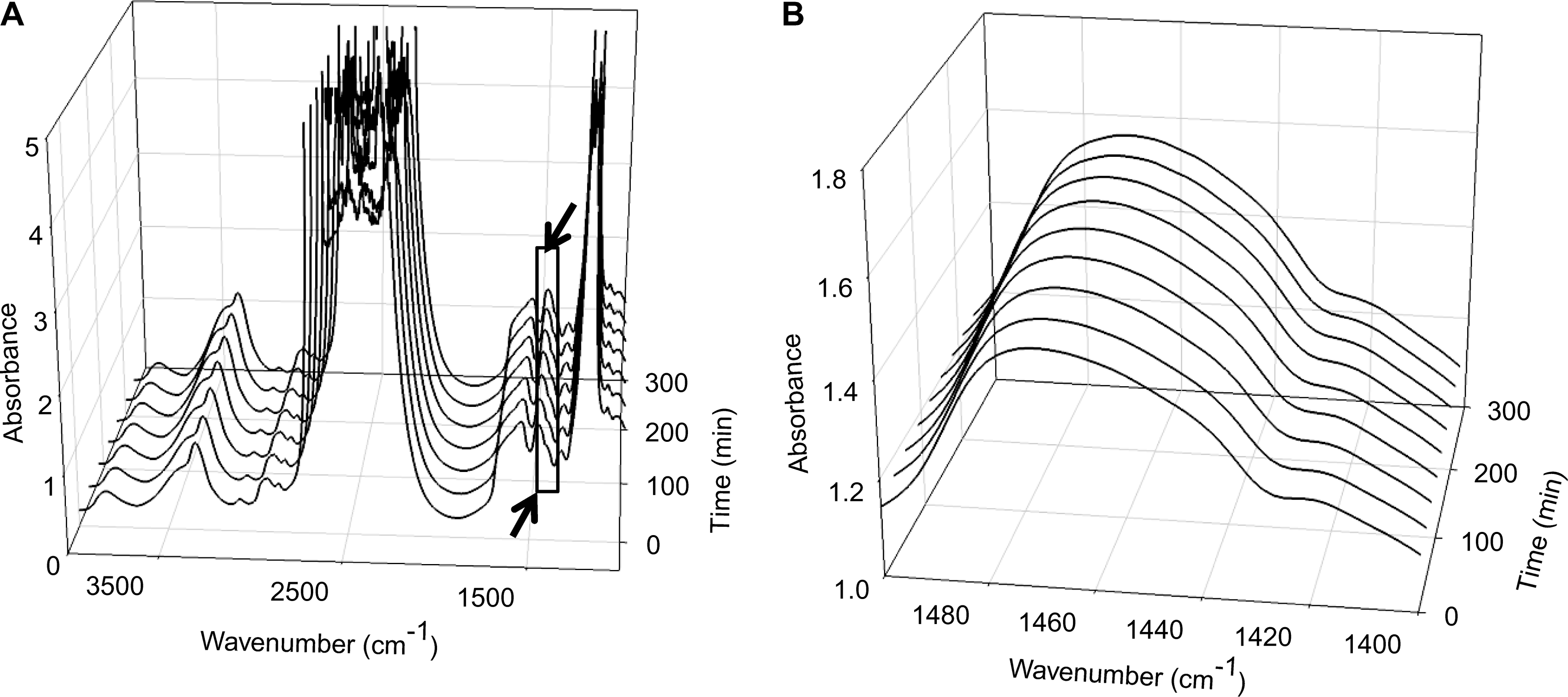
Solvent accessibility studies: (
Protein denaturation profile
The temperature-dependent FTIR spectra of control AlloDerm show changes in the amide-III band profile associated with changes in α-helical, triple helix/β-sheet, and random coil structures during thermal denaturation (Fig. 5); the intensity of the band at 1320 cm−1 decreases, the band at 1245 cm−1 shifts to a lower wavenumber, and a shoulder at approximately 1265 cm−1 becomes visible.7,9 The appearance of the band at 1265 cm−1 with heating is suggested to be due to an increase in the random coil structures upon heat denaturation. 9 Several structures tend to overlap in FTIR. There is no unambiguous assignment of triple helical structures in the amide-III region. It has been suggested that the triple helical structures overlap with β-sheet structures in the amide-III region, 21 which make the quantitative analysis of individual protein structural elements difficult. Nevertheless, FTIR provides accurate information about the configurational and structural changes in AlloDerm by detecting dynamic inter- and intra-molecular changes (Fig. 5) that capture denaturation onset and completion, as shown in Figure 6 and summarized in Table 1.
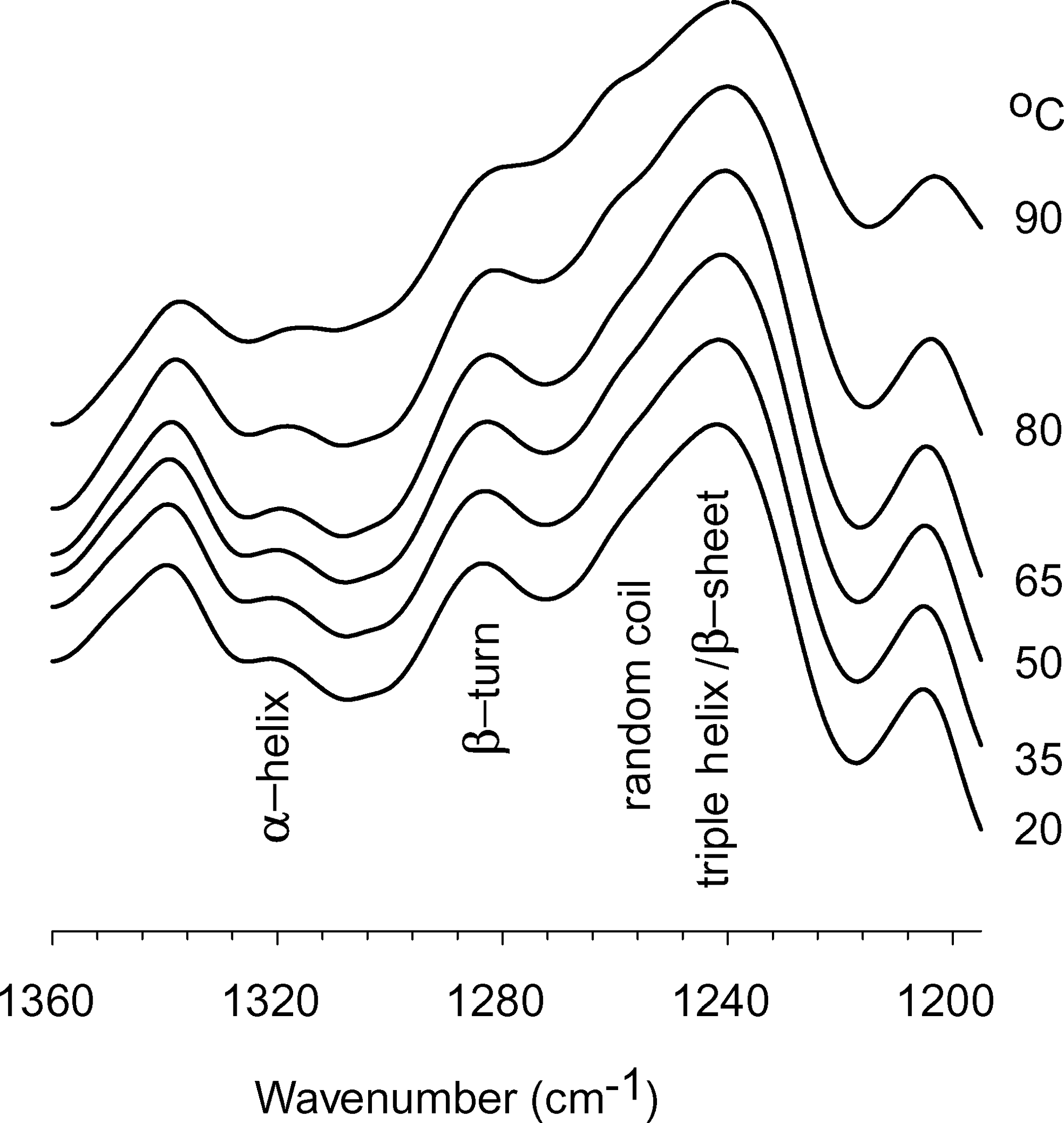
Temperature dependent Fourier transform infrared spectroscopy spectra show changes in amide-III band profile. Regions corresponding to α-helical, triple helix/β-sheet, and random coil structures are indicated.
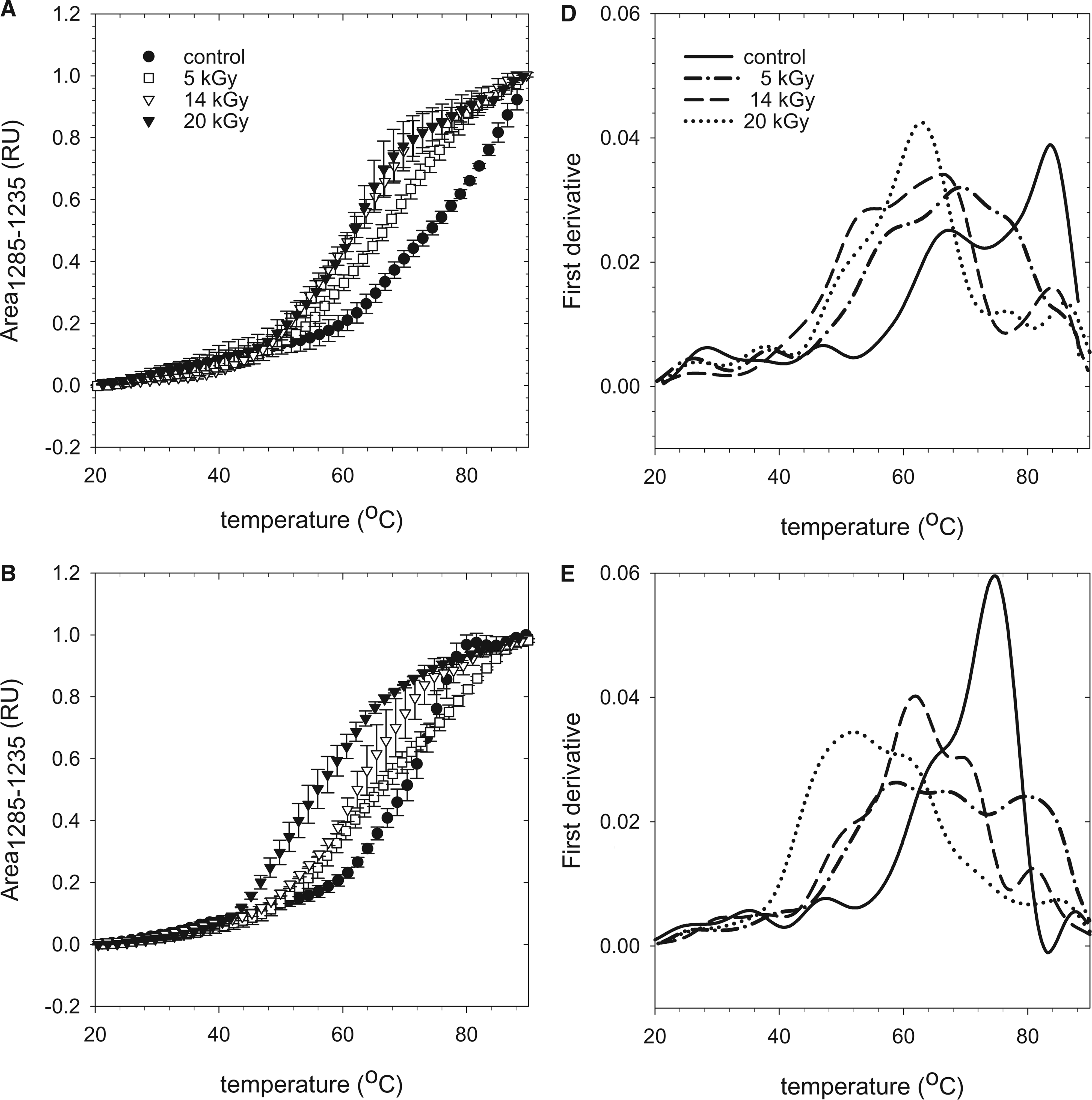
Denaturation profiles of control and gamma treated AlloDerm 1 h (
The data are the average of three different lots with corresponding standard deviations. Differential scanning calorimetry (DSC) data (on the right) are shown for comparison.
Average of two different lots.
Statistical significance determined by student t-test at p-value = 0.05.
Data obtained from Gouk et al. 1
Figure 6 depicts the protein denaturation profiles for control, 5-, 14-, and 20-kGy gamma-treated AlloDerm samples measured at varying times after rehydration of the tissue matrix (Fig. 6A, 1 h; Fig. 6B, 1 day; Fig. 6C, 1 week). Three different lots were analyzed for each of the above samples. The onset of protein denaturation occurs in a narrow temperature regime, with relatively little variation between the different lots that were tested (Table 1). Protein denaturation of control tissue matrix was found to commence at 62.3°C. The first-derivative analysis reveals two major denaturation transitions, with maxima at 66°C and 82°C. These two denaturation transitions remain visible 1 day and 1 week after rehydration (Fig. 6D, E). The maxima in the denaturation transitions, however, decrease to 61°C and 76°C 1 week after rehydration, and the onset temperature decreased to 59.6°C (Fig. 6F, C).
Gamma irradiation decreases the onset temperature of protein denaturation (Fig. 6 and Table 1). The onset temperature of protein denaturation was found to commence at 53.4°C, 48.7°C, and 46.7°C for the 5-, 14-, and 20-kGy-treated samples, respectively, after 1 h of rehydration (p < 0.05). First-derivative analysis shows that gamma treatment increases the complexity (number of denaturation transitions) of the denaturation profile, which could reflect the sequential denaturation of gamma-irradiation cleaved protein groups. The effect of gamma irradiation on the heat denaturation profile is also dependent on the time that the sample has been kept in the rehydration solution (1 h, 1 day, or 1 week). The differences in the denaturation profile seem to diminish over time and are hardly visible 1 week after rehydration, which indicates low availability of native protein in gamma-irradiated tissue matrices over time. Measurable differences in the onset denaturation temperature, however, remained visible 1 week after rehydration (Table 1).
Discussion
Currently, gamma irradiation is a preferred method for sterilization of biomedical devices and engineered tissue matrices. However, native tissues are collagen-based materials that are susceptible to gamma irradiation. Post-gamma irradiation structural and biochemical alterations in collagen-based materials have been well documented,2,22 and they can, in turn, affect biochemical and biomechanical properties and tissue performance in vivo. Gamma irradiation has been shown to cause molecular fragmentation, inter- and intra-molecular cross-linking, secondary structure disorganization, and protein denaturation, even at low doses.2,3,23–25 Structural alterations of this nature can be precisely studied using FTIR. FTIR has proven to be a versatile analytical tool to study protein conformations.8,26–28 It is particularly useful for studying complex tissue matrices because of the availability of a wide range of sampling methods (transmission, emission or reflection) depending upon sample constraints.29–31 In the present study, we used the transmission mode for hydrated tissue matrix analysis and the reflection (ATR) mode for dry tissue matrix analysis. In the dried state, tissue matrix thickness restricted analysis using the transmission mode. Results from both sampling methods provided complementary information about protein conformation changes within tissue matrix.
Our results clearly demonstrate a dose-dependent effect of gamma irradiation on AlloDerm. Specifically, gamma irradiation induces changes in protein secondary structure, protein stability, and protein solvent accessibility. Although changes in protein structure are not apparent in the dried state (Fig. 1), it is possible that the effects are masked because molecular mobility is too low to allow protein structural changes. On the other hand, gamma irradiation changes the fingerprint region of the spectrum (between 1200 and 1000 cm−1). The bands in this region arise predominantly from the stabilizing excipient molecules. The changes in protein:excipient ratio that were observed here could be indicative of chemical reactions between proteins and sugars; however, details of these reactions are not well understood at this point and would require further studies.
Upon rehydration, random coil structures in gamma-treated tissue matrix increase at the expense of ordered β-sheets (Fig. 2B). When monitored over time in rehydration solution (1 h and 4 h), the differences between gamma-treated and control tissue matrices become smaller, indicating dissociation of the condensed tissue matrix (Fig. 3). We speculate that gamma irradiation degrades or breaks proteins into smaller fragments that result in condensation of the tissue matrix and that aggregated protein structures dissociate over time. These results are consistent with previous observations of a lower swelling ratio of gamma-irradiated tissue matrices, greater susceptibility to enzyme degradation, and destabilization of collagen. 1 H/D exchange studies further support our speculation. The major protein fraction of a control tissue matrix is much more solvent accessible than a gamma-sterilized tissue matrix, which shows a slow H/D exchange rate (Fig. 4C). The rapid exchange in the control tissue matrix points to native loose configuration of the tissue matrix, which is readily accessible to the solvent. In the control tissue matrix, a smaller protein fraction shows a slower exchange rate, which highlights a different proton population that resides in the inner hydrophobic cores of proteins involved in stable H-bonding. Unlike the two distinct exchange rate constants in the control tissue matrix, the gamma-sterilized tissue matrix shows only one. The gamma-treated tissue matrix displays a clearly lower exchange rate than the control tissue matrix. Slow dissociation of tissue matrix in solution due to rehydration allows weakly H-bonded and structurally unstable regions of the proteins to be accessible to solvent. Scanning electron microscopy studies on AlloDerm treated with various levels of gamma radiation 1 provides visual basis for this theory. These images show that gamma-irradiated tissue matrices exhibit a collagen network that is more condensed than the loose mesh network of the nonirradiated tissue matrix. 1
Differential scanning calorimetry (DSC) studies reveal a decrease in the onset temperature and denaturation enthalpy with increasing dosage of gamma irradiation. 1 Temperature scan FTIR studies are in good agreement with these DSC studies, as shown in Table 1. The minor differences in onset temperature (58°C with DSC, 60°C with FTIR) are probably related to differences in the experimental conditions such as the heating rate and doses of gamma irradiation. One notable difference between FTIR and DSC is the continued denaturation above 70°C, which is not observed in DSC. FTIR shows a clear denaturation transition at high temperatures that is not evident in the DSC traces. This denotes that FTIR detects structural alterations that are not measurable using DSC approaches. The presence of multiple denaturation events is evident after all rehydration times (Fig. 6), suggesting the heterogeneous nature of the tissue matrix. This, together with the lower thermal stability after gamma irradiation, suggests the presence of multiple protein fragments in the condensed tissue matrix. The onset denaturation temperature progressively decreases with increasing rehydration for gamma-treated tissue matrices, and the differences in denaturation profile at different doses of gamma irradiation seem to diminish over time. In the control tissue matrix, the onset denaturation temperature also decreases over time but not as severely as in gamma-treated tissue matrix. In addition, more-separate denaturation transitions are visible in the gamma-treated tissue matrix. One possible explanation for this could be that gamma irradiation degrades proteins into smaller fragments, which aggregate upon heating because of loss of hydrogen bonds. At supra-physiological temperatures, random aggregations of these already denatured protein molecules coagulate, resulting in multiple transitions upon heating (Fig. 6).
In summary, we have demonstrated that FTIR provides a fast and non-invasive way to detect post-processing protein structural changes in complex biological tissue matrices. Results show that gamma irradiation significantly alters protein structure and stability and protein–protein interactions in AlloDerm. Solvent accessibility and heat denaturation studies indicate fragmentation of the tissue matrix, which is prominent at high doses. These results are in agreement with data from calorimetric, histological, and scanning electron microscopy studies, demonstrating the matrix collapse reported previously. Additional studies are required to correlate protein structural changes to in vivo behavior of the tissue matrix in terms of wound healing and tissue remodeling through host cells.
Footnotes
Acknowledgments
This project was financially supported in part by LifeCell Corporation. Parts of this work were carried out in the Minnesota Characterization Facility, which receives partial support from the National Science Foundation through the National Nanotechnology Infrastructure Network program.