
Abstract
The commonly applied cryopreservation protocols routinely used in laboratories worldwide were developed for simple cell suspensions, and their application to complex systems, such as cell monolayers, tissues, or biosynthetic constructs, is not straightforward. In particular for monolayer cultures, cell detachment and membrane damage are often observed after cryopreservation. In this work, combined strategies for the cryopreservation of cells attached to Matrigel-coated well plate's surfaces were investigated based on cell entrapment in clinical-grade, ultra-high viscosity alginate using two cell lines, neuroblastoma N2a and colon adenocarcinoma Caco-2, with distinct structural and functional characteristics. As the cryopreservation medium, serum-free CryoStor™ solution was compared with serum-supplemented culture medium, both containing 10% DMSO. Using culture medium, entrapment beneath an alginate layer was needed to improve cell recovery by minimizing membrane damage and cell detachment after thawing; nevertheless, up to 50% cell death still occurred within 24 h after thawing. The use of CryoStor™ solution represented a considerable improvement of the cryopreservation process for both cell lines, allowing the maintenance of high postthaw membrane integrity as well as full recovery of metabolic activity and differentiation capacity within 24 h postthawing; in this case, entrapment beneath an alginate layer did not confer further protection to cryopreserved Caco-2 cells, but was crucial for maintenance of attachment and integrity of N2a neuronal networks.
Introduction
The commonly applied protocols for cryopreservation of cell suspensions consist of slow cooling and rapid warming rates in cell culture media containing high concentrations of DMSO and animal-derived products. It does not take into account the differences between complex cell systems and simple cell suspensions with respect to their responses to cooling, warming, and dehydration, which often results in poor postpreservation quality. 3 Thus, while many cells in suspension can be cryopreserved with almost complete recovery, it has been demonstrated that cells in monolayers are often more susceptible to cryoinjury.4–8 Cell–cell and cell–matrix junctions and the organization of the cytoskeleton have been shown to render cells more sensitive to cryoinjury when compared to isolated cells in suspension.5,9 Further, the close interactions between cells are potential sites for damage caused by the osmotic stresses and phase changes involved in the process of cryopreservation. 10 Finally, there appears to be an enhanced susceptibility to intracellular ice formation (IIF) in cells adherent to a substrate when compared to cells in suspension, even in the absence of cell–cell interactions,4,11,12 although the exact relation between IIF and cryoinjury in adherent cells is still unclear. 13
Successful cryopreservation of ready-to-use cell monolayers would support the implementation of routine cryopreservation practices during preparation of engineered cells and tissues for clinical applications 14 and benefit high-throughput cell-based assays by reducing batch-to-batch variation and eliminating the time-consuming, labor-intensive process of inoculation and expansion from a frozen vial of cells, thus reducing time between cell storage and use in experimental settings.
Different cryopreservation methodologies alternative to the commonly applied protocols are reported in the literature; some refer to the use of structured substrates, 15 cell entrapment on extracellular matrix (ECM)16,17 or gels, the use of other cryoprotectants rather than DMSO,7,16,17 or the use of solutions specially designed to reduce the ionic and hydraulic imbalances induced in cells during hypothermia.18–20
Cell entrapment in a gel or ECM has been proposed as a promising cryopreservation strategy.16,21–25 Among the biopolymers used in biotechnological and medical applications, clinical-grade, highly purified alginate of extremely high viscosity extracted by a novel process and uniformly cross-linked with Ba2+ has been described as an improvement for the entrapment of cells, tissues, or other biologically active material. 26
In this study, a novel cryopreservation strategy based on cell entrapment beneath a layer of clinical-grade, ultra-high viscosity permeable alginate hydrogel was investigated. Two cell lines, mouse neuroblastoma N2a and human colon adenocarcinoma Caco-2, were used as model systems due to their distinct structural and functional characteristics as well as cell–cell and cell–matrix interactions.
Caco-2 cells, after long-term culture (15–25 days), spontaneously differentiate to cells expressing morphological and functional characteristics of the mature intestinal enterocytes,27,28 which renders them valuable in vitro models and high-throughput screening tools in the pharmaceutical industry.29,30 Neuroblastoma cells are able to differentiate into randomly connected neuronal populations through treatment with retinoic acid, which has promoted their extensive use as model systems to study neuronal communications and screen novel compounds for neurotoxic properties.31–34 Successful cryopreservation of ready-to-use micro-plates of both cell lines would reduce heterogeneity of sub-populations, thus promoting the standardization of these models and data comparison from different laboratories, and support the implementation of faster drug screening assays.35,36
The use of standard cell culture media was compared with CryoStor™ solution as the vehicle solution for the cryoprotectant (DMSO). CryoStor™ is a protein- and serum-free, intracellular-like (low Na+, high K+) cryo-medium designed to meet the specific molecular requirements of individual cells and tissues, which has been reported to significantly enhance postthaw cell survival and recovery in a series of cellular systems3,37–42 and thus could further potentiate the advantages of alginate encapsulation.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), trypsin-EDTA (1 × solution), and alamarBlue were obtained from Invitrogen (Carlsbad, CA). RPMI-1640 medium, poly-L-lysin, poly-D-lysin, retinoic acid, DMSO, fluoresceine diacetate, and ethidium bromide were obtained from Sigma (Steinheim, Germany). Culture plasticware was obtained from Nunc (Roskilde, Denmark). Matrigel™ was obtained from BD Biosciences (San Jose, CA). CryoStor™-CS10 was obtained from BioLife Solutions (Bothell, WA). Other chemicals were of the purest grade available from regular commercial sources.
Alginates
Ultra-high viscosity, clinical-grade alginates (viscosity of a 0.1% [w/v] solution in distilled water was 20–30 mPa · s) were produced and purified from stipes of Lessonia nigrescens and Lessonia trabeculata as described elsewhere.26,43–45 A 1:1 mixture of purified alginates was used at 0.7% (w/v) in NaCl.
Cell culture
The Caco-2 cell line was obtained from the Deutsche Sammlung von Mikroorganismen und Zellkulturen (Braunschweig, Germany). Cells were maintained in RPMI-1640 medium supplemented with 10% (v/v) heat-inactivated FBS and sub-cultured every 3–5 days. Differentiation into enterocyte-like cells was achieved by long time culturing (up to 21 days) with medium exchange every 3–4 days.
The N2a cell line was obtained from the European Collection of Cell Cultures (Salisbury, UK). Cells were maintained in DMEM supplemented with 10% (v/v) heat-inactivated FBS and 4.5 g/L glucose and sub-cultured every 5 days. For induction of differentiation, the medium was replaced by DMEM + 2% FBS + 15 μM retinoic acid 24 h postplatting and changed every 2 days during 5 days.
Both cell lines were maintained at 37°C in a humidified atmosphere of 5% CO2 in air. All experiments were performed using Caco-2 and N2a cells between passages 10–30.
Cell culture beneath an alginate gel layer
Caco-2 and N2a cells were seeded onto sterilized glass coverslips (Marienfeld GmbH, Lauda-Königshofen, Germany) coated with Matrigel™ (0.28 mg/mL in culture medium), inside four-well plates, at a density of 1 × 104 and 5 × 103 cells · cm2, respectively, and cultured for 5 days in the respective growth medium or differentiated (as described above).
An alginate gel layer was added on top of the cells either on day 1 or on day 4 postinoculation to assess the best addition time for alginate. Cells were rinsed in phosphate-buffered saline (PBS), incubated with poly-L-lysin for 30 min (to allow adhesion of alginate to the well's surface), and washed with PBS before addition of 100 μL/well alginate gel (approximately 2.5 μm thickness, as estimated through electron microscopy). Alginate gelling was achieved through 25 min incubation with 20 mM BaCl2 solution followed by three times washing with PBS with 0.5 mM MgCl2.
Cells cultured in the same conditions without an alginate layer were used as control.
Cell cryopreservation
Undifferentiated Caco-2 and N2a cells were frozen after 5 days of culture inside the four-well plates with 300 μL/well cryopreservation solution, which consisted of either culture medium supplemented with 10% DMSO or CryoStor™-CS10. Differentiated N2a and Caco-2 cells were similarly cryopreserved (after 5 and 21 days of culture, respectively) in CryoStor™-CS10. Cells were allowed to equilibrate in the cryopreservation solution for 30 min at 4°C and cooled to −80°C, at a cooling rate of 1°C/min, using a rate-controlled freezing system (Planer Kryo 560–16; Planer, Middlesex, UK). The plates were stored at −80°C for 1–2 weeks. After storage, cells were thawed in a CO2 incubator (37°C, 5% CO2 in air), (approximately 15°C/min warming rate), and a one-step dilution (1:3) in growth medium was performed. Total growth medium replenishment was performed 30 min after, and cells were further cultured for postthaw studies or used for membrane integrity assays immediately after thawing. The alginate layer remained on top of the cells during further culture and recovery assays.
Cell recovery assays
Cell viability was assessed before freezing and after thawing daily for 3 days, both through a qualitative cell membrane integrity assay and the quantitative alamarBlue assay. Cell growth and recovery after thawing was also monitored during 3 days by visualization on an inverted microscope (Leica DM IRB; Leica Microsystems GmbH, Wetzlar, Germany).
Cell membrane integrity assay
The enzyme substrate fluorescein diacetate and the DNA-dye ethidium bromide were used as described in the literature. 46
AlamarBlue™ assay
The metabolic indicator alamarBlue was used following the manufacturer's recommendation. Briefly, cells were incubated with fresh medium containing 10% alamarBlue, and 4 h later fluorescence was measured in 96-well plates using a microplate fluorescence reader (Biotek Instruments, Winooski, VT).
Cell differentiation assessment
Caco-2 differentiation
The differentiation state of postconfluent Caco-2 monolayers before and after cryopreservation as well as the capacity of confluent (5 days culture) Caco-2 cells to spontaneously differentiate during long-term postthaw culture were evaluated by quantifying the activity of the brush-border enzyme alkaline phosphatase. This was performed by measuring the reaction of the enzyme with the substrate 4-methylumbelliferyl phosphate as described in the literature. 46
N2a differentiation
Neuronal differentiation was assessed before and after cryopreservation using an immunocytochemistry assay with neuronal cell markers. For cells covered with an alginate layer, this was dissolved before performing the assay by incubating with a 20 mM Na2SO4 solution for 20 min at 37°C in a humidified atmosphere of 5% CO2 in air. Cells adherent to glass coverslips were processed as described previously. 47
Primary antibodies and dilutions used were anti-tubulin beta III isoform (β-tubulin, 1:10; Chemicon, Temecula, CA) and anti-microtubule–associated protein-2 (MAP2a/b, 1:100; Sigma). The secondary antibodies and dilutions used were anti-mouse Alexa 488 (1:500) and anti-rabbit Alexa 594 (1:500) (Molecular Probes, Eugene, OR). Cells were visualized in a Leica DMRB fluorescence microscope (Leica Microsystems GmbH).
Scanning electron microscopy
To evaluate the structural integrity and differentiation state of Caco-2 monolayers, scanning electron microscopy (SEM) was performed before and after cryopreservation. Cells adherent to the glass coverslips were prepared as described elsewhere, 48 and SEM studies were performed on field emission scanning electron microscope–type FESEM XL30 (Phillips, Eindhoven, The Netherlands).
Data presentation/analysis
Each experimental condition represents n ≥ 6 experimental replicates (minimum of two experimental repeats with three sample replicates per experiment). Error bars denote the standard deviation of the mean. Comparison of mean values between groups was performed using analysis of variance (ANOVA), with p < 0.05 considered statistically significant. Representative photographs are depicted.
Results
Effect of alginate entrapment on cell growth, morphology, and differentiation
Cell entrapment beneath a layer of alginate gel added 1 or 4 days postinoculation resulted in different responses to growth and differentiation of both cell lines as shown in Figure 1.
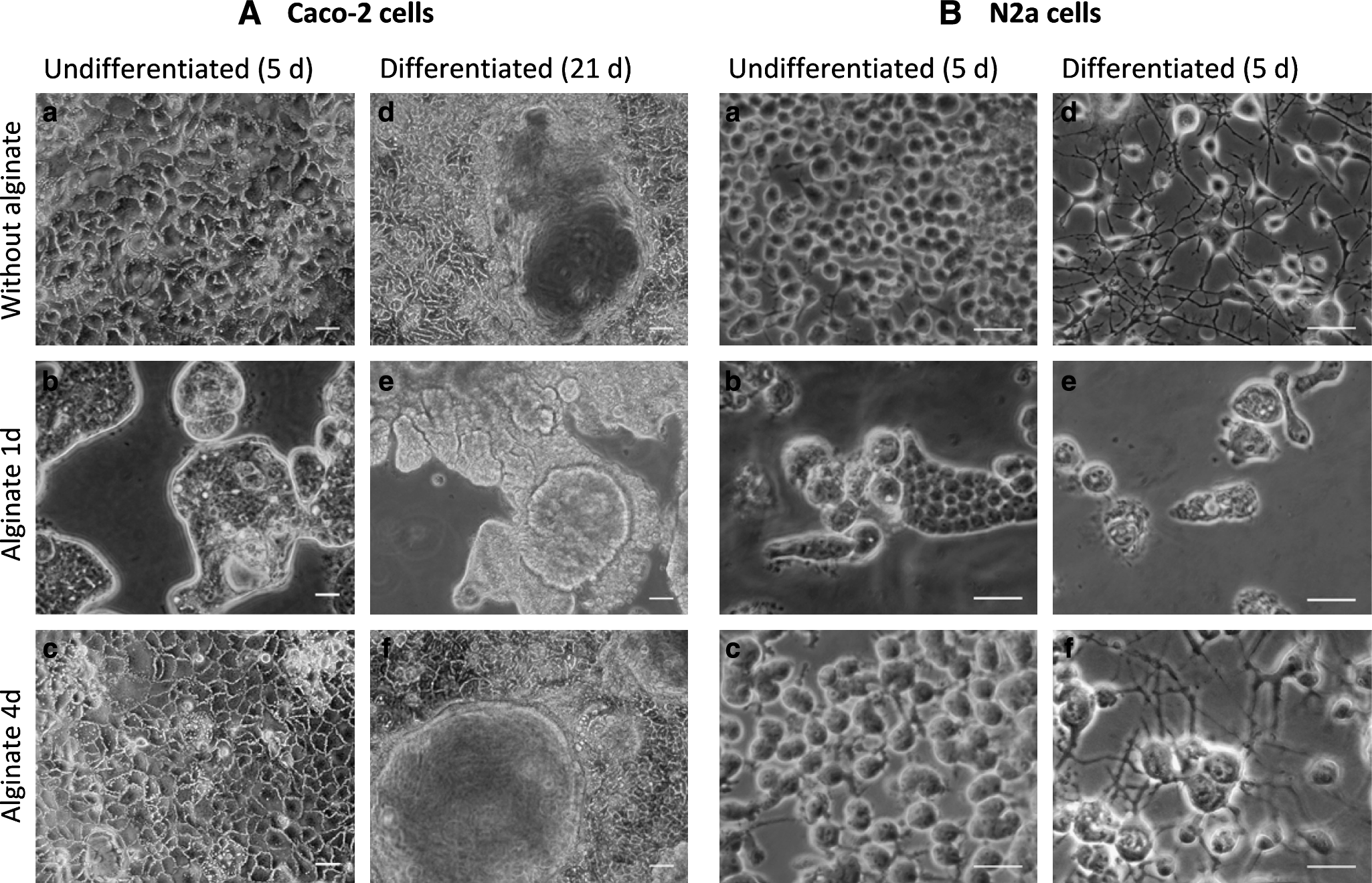
Effect of alginate entrapment on Caco-2 and N2a cell cultures on four-well plates. Phase-contrast photographs of Caco-2 (
Control cultures (without alginate) of Caco-2 and N2a cells formed confluent monolayers after 4 days in culture (Fig. 1Aa, Ba). With alginate added 1 day postinoculation, both cell lines lost their characteristic shape and grew beneath the alginate layer as closely packed cell aggregates displaying round cell morphology (Fig. 1Ab, Bb). Further, Caco-2 cells proliferated at lower rates when compared to control cultures as confirmed through alamarBlue assays (Fig. 1C). Whenever the alginate was added 4 days postinoculation, Caco-2 cells' capacity to proliferate and differentiate during long-term culture was not affected as shown by metabolic activity profiles (Fig. 1C) and the development of the morphological features typical of enterocyte differentiation, such as domes and flower-like clusters on the cell surface (Fig. 1Af). The difference in fluorescence values observed between control and alginate-covered cells (Fig. 1C) might be due to a delayed diffusion of the metabolic products through the alginate matrix leading to the incomplete alamarBlue reaction. After alginate dissolution, no significant difference was found between control and alginate-entrapped cells, confirming that alginate entrapment 4 days postinoculation does not affect cell growth (data not shown).
Concerning N2a cells' neuronal differentiation, whereas alginate addition at day 1 resulted in significant reduction of neurite sprouting (Fig. 1Be) as well as inhibition of neuronal markers expression (data not shown), adding alginate 4 days postinoculation did not affect the morphology of the neuronal networks (Fig. 1Bf) or the expression of typical neuronal markers, as confirmed through immunocytochemistry (data not shown).
Effect of alginate entrapment and cryopreservation solution on postthaw recovery and monolayer integrity of undifferentiated cells
Cryopreservation efficacy was evaluated by cell recovery postthawing, assessed by the alamarBlue assay as previously described.7,49,50 Additionally, loss of membrane integrity and cell detachment from the surface after thawing were qualitatively evaluated using a dual fluorescent membrane integrity test (Fig. 3).
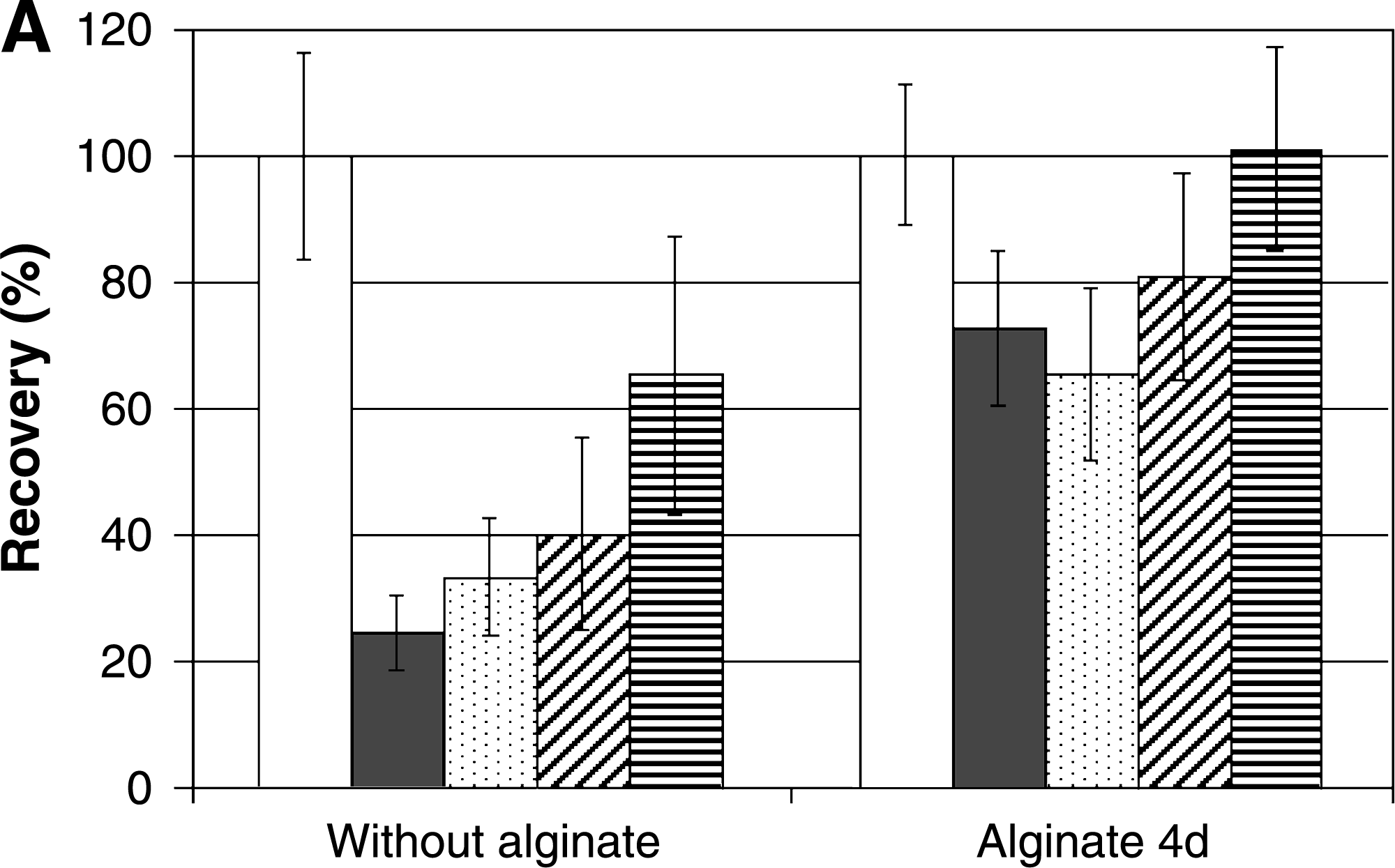
As shown in Figure 2, cryopreservation of both cell lines in culture medium without alginate led to a strong drop in metabolic activity immediately after thawing: 97 ± 1% and 53 ± 14% for N2a and Caco-2 cells, respectively. It is important to notice that such high loss of metabolic activity resulted from both damage of the cell membrane as well as high cell detachment from the well's surface shortly (within 30 min to 4 h) after thawing (Fig. 3), leading to cell loss when medium was exchanged during alamarBlue assays. Although the few cells that remained attached to the wells were able to recover and resume growth, only 17 ± 8% and 43 ± 18% recovery was found 3 days after thawing for N2a and Caco-2 cells, respectively.

Effect of cryopreservation medium and alginate entrapment on the recovery of Caco-2 (

Effect of cryopreservation medium and alginate entrapment on the membrane integrity of Caco-2 cells after cryopreservation. Cells were cryopreserved in culture medium or CryoStor™-CS10 solution in an undifferentiated state after 5 days of culture without alginate or beneath a layer of alginate added 4 days postinoculation, and assayed during 3 days after thawing with a dual fluorescence membrane integrity test: green cells with undamaged membranes and metabolic activity; red cells with damaged membranes. Scale bars = 50 μm.
Entrapment beneath a layer of alginate added 4 days postinoculation led to improved recovery of culture medium cryopreserved cells immediately after thawing (90 ± 33% and 63 ± 6% for Caco-2 and N2a cells, respectively). This was due to reduction of both membrane damage and cell detachment after thawing, as shown in Figure 3 for Caco-2 cells. Despite such improvement, a 50% lost of cell viability still occurred within 24 h postthawing, when cells with damaged membranes were found beneath the alginate layer (Fig. 3). Nevertheless, after 3 days in culture, both cell lines were able to recover approximately 90% of the initial, that is, before freezing, metabolic activity (Fig. 2).
As depicted in Figures 2 and 3, the use of CryoStor™-CS10 solution resulted in a considerable improvement of cell recovery immediately after thawing when compared to standard culture medium: 91 ± 15% and 68 ± 15% for Caco-2 and N2a cells, respectively. In this case, entrapment beneath an alginate layer added 4 days postinoculation did not confer further protection to undifferentiated Caco-2 and N2a cells as high metabolic activity was found for both nonentrapped and alginate-entrapped cells immediately after thawing. Further, full recovery of metabolic activity, initiation of proliferation, and maintenance of cell membrane and monolayer integrity were observed within 24 h postthawing (Figs. 2 and 3).
Concerning the moment of alginate entrapment, studies with Caco-2 cells revealed no significant difference on the immediate postthaw recovery of cells cryopreserved beneath a layer of alginate added either 1 or 4 days postinoculation: 88 ± 23% or 89 ± 24% and 90 ± 33% or 102 ± 31%, for cells cryopreserved in culture medium and CryoStor™-CS10, respectively. Thus, and because alginate entrapment 1 day postinoculation led to a considerable inhibition of cell growth when compared to control cultures, alginate entrapment at day 4 was chosen for the continuation of the experimental work.
Effect of cryopreservation upon proliferation and differentiation capacity of Caco-2 cells
The capacity of Caco-2 cells to proliferate and differentiate when long-term (18 days) cultured after thawing is shown in Figures 4 and 5. One can observe that nonentrapped cells cryopreserved in culture medium were able to initiate growth only 3 days after thawing, which resulted in a significantly lower number of metabolically active cells in culture 18 days after thawing as compared to CryoStor™-CS10 cryopreserved and control cells (Fig. 4Aa). When entrapped beneath a layer of alginate, added either 1 or 4 days postinoculation, despite an initial delay in growth initiation, cells cryopreserved in culture medium were able to proliferate as both control cells and CryoStor™-CS10 cryopreserved cells; no significant difference was found in the number of metabolically active cells in culture after 18 days between the three samples (Fig. 4Ab, Ac).

Effect of cryopreservation medium and alginate entrapment on (

SEM photographs of Caco-2 monolayers after long-term culture (18 days) after cryopreservation in culture medium (
Concerning cell differentiation, as shown in Figure 4B, the percentage of alkaline phosphatase expressed after 18 days of culture postthawing by CryoStor™-CS10 cryopreserved cells was comparable to the expression of this enzyme by control cells. No difference was found between cells cryopreserved with or without alginate entrapment. On the other hand, for cells cryopreserved in culture medium, alginate entrapment led to a considerable increase in the percentage of alkaline phosphatase expressed.
For cells cryopreserved in CryoStor™-CS10, either with or without an alginate gel layer, SEM observations of Caco-2 cells cultured for 18 days postthawing confirmed the high confluence, development of multiple cell layers, and formation of a thick carpet of microvilli (Fig. 5e, f and 5c, d). On the other hand, for cells cryopreserved in culture medium, full monolayer confluence could still not be observed and cell connections were still being formed after 18 days in culture (Fig. 5a, b).
Effect of alginate entrapment and cryopreservation solution on postthaw recovery and monolayer integrity of differentiated cells
As confirmed by the results presented above, freezing cells in CryoStor™-CS10 solution without alginate or beneath a layer of alginate added 4 days postinoculation resulted in higher cell recoveries postthawing. These conditions were thus chosen for the cryopreservation of both cell lines in a fully differentiated state.
Caco-2 cells were frozen after 23 days of culture, which corresponded to fully differentiated cultures, forming multiple layers and covered by highly dense microvilli. As seen in Figure 6A, when cells were frozen with or without alginate entrapment, although high recovery was found immediately postthawing (88 ± 10% and 96 ± 5%, respectively), a decrease in cell metabolic activity was observed until 3 days postthawing, resulting in 55 ± 7% and 70 ± 10% metabolic activity, respectively; full recovery of cell metabolic activity could only be obtained 10 days after thawing.

Effect of alginate entrapment on the recovery (
SEM observations revealed different responses to freezing in different regions of the monolayer (Fig. 6): both regions of undamaged cell surface, covered with typical thick carpet pattern of intact microvilli (Fig. 6Ba, Bb, Bc), as well as areas with damaged cells at the monolayer's surface (Fig. 6Bd, Be) were found within the same well. It was further observed that, although surface morphology with intact microvilli and close junctions were maintained in some areas, the typical tissue-like structure that grew above the average height of the monolayer suffered considerable shrinkage (Fig. 6Bf, Bg), throughout the whole monolayer independently of the position of the cells within the well.
As shown in Figure 7A for CryoStor™-CS10 cryopreserved N2a cells as fully differentiated neuronal networks, with or without alginate entrapment, a drop in cell metabolic activity was found immediately after thawing (72 ± 12% and 24 ± 6%, respectively). Although neuronal networks cryopreserved without alginate remained attached to the well's surface immediately after thawing (Fig. 7Ba), it was found that 24 h later neurite connections would break and cells would detach from the well's surface (Fig. 7Bb) and eventually be lost during alamarBlue assays. For alginate entrapped cells, however, the neuronal network integrity was maintained (Fig. 7Bd) and 100% cell metabolic activity was recovered within 3 days postthawing. Further, after 1 day in culture postthawing, cells were expressing the typical neuronal differentiation markers beneath the alginate gel layer, as shown in Figure 7C.
Effect of alginate entrapment on the recovery (
Additionally, alginate could efficiently be dissolved by Na2SO4 without causing any damage to cell morphology or function, as shown through SEM (Figs. 5e, f, and 6B) for Caco-2 and immunocytochemistry for N2a cells (Fig. 7C).
Discussion
Previous data from our lab has shown that routine cryopreservation of Caco-2 and N2a cell lines in suspension using a slow-freezing cryopreservation protocol in cell culture medium (containing 10% DMSO) yields over 90% recovery postthawing. 46 However, when cryopreserved as confluent monolayers on well plates, high loss of metabolic activity and membrane integrity after thawing were observed. These results are in accordance with previous reports of loss of viability and excessive cell detachment for different cell lines cryopreserved as monolayers.6,7,51 The presence of cell–cell and cell–surface interactions has been shown to render cells more susceptible to freezing injury.4,5,52 Such interactions are likely sites for monolayer damage by the osmotic stresses and phase changes involved in the process of cryopreservation10,53 and have been associated with enhanced susceptibility to intracellular ice formation.4,11,54 Further, the extended morphology of attached cells may create conditions for cryopreservation-induced damage to the cell ultra-structure (cytoskeleton, focal adhesions, or gap junctions) due to mechanical forces, such as extracellular ice and differential thermal contraction between cells and substrates.55,56
In the present study, cell entrapment beneath a layer of ultra-high viscous alginate gel was investigated as a strategy to improve the cryopreservation of both cell lines as undifferentiated or fully differentiated monolayers. To develop biosynthetic conduits for cell culture and cryopreservation, the selected hydrogels must be tested for their compatibility with cells. Thus, the morphology of each cell line was closely monitored during proliferation and differentiation assays before freezing. The results presented herein demonstrate that the addition of an alginate layer over cells cultured for 4 days on Matrigel-coated surfaces does not influence cell growth or differentiation, which confirms the possibility of using this ultra-high viscosity alginate as a biocompatible matrix for cell entrapment. 57 Nevertheless, for both cell lines investigated, alginate addition 1 day postinoculation inhibited metabolic activity and affected cell morphology and differentiation. This difference may be due to the fact that at day 1 postinoculation most of the cells in culture were loosely attached, as indicated by their round morphology, while after four cells have already developed strong cell–cell and cell–surface contacts. Thus, we hypothesize that alginate addition 1 day postinoculation inhibits cell attachment and spreading on the surface, which in turn limits cell migration and division under the alginate. This might also explain the inhibition of neurite extension of N2a cells entrapped in alginate, which may not be able to establish the proper connections with the ECM required for axon growth. Impaired cell growth in alginate has also been reported for other cell lines, such as olfactory ensheathing cells and Schwann cells, 58 whereas murine insulinoma 59 and fibroblasts 60 have been shown to rapidly proliferate in alginate beads. This contradictory data might be due to differences in the type and characteristics, namely, different viscosities, of the alginate used, which will influence the diffusion rates of oxygen, nutrients, and differentiation factors throughout the matrix as well as the mechanical constrains imposed to the cells within the gel.59,61,62
The results presented in this paper show that cryopreservation strategies based on ultra-high viscous alginate gel entrapment led to improved recovery of metabolic activity and function of Caco-2 and N2a cell lines adherent to Matrigel-coated culture well plates. Our results demonstrate that alginate entrapment improves the recovery of cells cryopreserved in cell culture medium by reducing membrane damage and cell detachment immediately after thawing. Further, it was found that the proliferation and differentiation capacities during long-term culture of Caco-2 cells cryopreserved beneath an alginate layer added 4 days postinoculation were not affected, which is a critical issue for cells intended to be used on therapeutic or pharmacological purposes immediately after thawing. Despite Matrigel has previously been shown to promote improved adherent stem cells recovery after cryopreservation, 16 in the present study Matrigel coating by itself had no significant impact on the survival of both cell lines, as similar recovery values were found for cells cryopreserved on uncoated and Matrigel-coated surfaces (results not shown).
Cell entrapment by micro-encapsulation within alginate beads has previously been reported as a promising strategy to improve postthaw viability and function of cell types that were shown to poorly survive the cryopreservation process, such as hepatocytes63–65 and pancreatic islets,66–69 although the underlying mechanisms are still unclear.
To our knowledge, this is the first report on the cryopreservation of adherent cells beneath a layer of alginate gel. Our results suggest that monolayer entrapment beneath a layer of alginate improves cell recovery by providing cell immobilization, thus avoiding detachment from the surface and breakage of cell–cell interactions. Further, as a considerable increase in the number of attached cells with undamaged membranes was found immediately after thawing, it seems plausible that the presence of the gel not only avoids physical cell rupture but also mitigates cryopreservation-induced cell death. Such effect may be related with the influence of the polymeric matrix of the gel on extracellular ice formation,70,71 which is known to be involved in cryo-induced injury due to modification of the mechanical chemical and environment of the cell2,72–74 as well as direct involvement in initiation of ice formation inside cells.73,75,76
Despite the improved cell recovery immediately after thawing for culture medium cryopreserved cells beneath alginate, up to 50% cell death still occurred within 24 h postthawing. In agreement with reports on the literature describing postthaw survival overestimates, such decrease in cell viability might be related to sequential apoptotic and necrotic processes not evident immediately subsequent to thawing.49,77 In this sense, the use of CryoStor™ solution resulted in a considerable improvement of the cryopreservation process for both cell lines in an undifferentiated state, either with or without alginate gel entrapment: high postthaw membrane integrity and metabolic activity as well as the structural integrity of the confluent monolayers were maintained, not only immediately after thawing but also for after 3 days in culture.
CryoStor™ has been developed from the platform hypothermic storage solution Hypothermosol, which is part of a number of preservation solutions currently available in the field of regenerative medicine.78,79 These solutions have been carefully formulated to maintain the ionic and hydraulic balances of cells at low temperatures, thus facilitating the preservation of cell homeostasis and control of the ionic environment that cannot be achieved using traditional preservation media formulations consisting of a basal culture media with serum protein and Me2SO supplementation.18,20,80–82 The vehicle solutions Euro-Collins and Unisol-cryoprotectant have shown to allow the cryopreservation of two vascular cell lines as monolayers, although only 50% recovery of metabolic function immediately after thawing was obtained and full recovery could only be observed 3 days postthawing. 19 Besides its intracellular-type (low Na + , high K+) composition, CryoStor™ is supplemented with several additional components to reduce the generation of free radicals or energy deprivation at nonphysiological temperature and inhibit apoptosis. 49 The improved cell survival and function of different cell types, such as hepatocytes, cord blood, or stem cells, when cryopreserved in suspension in CryoStor™ solution, has been related to a direct reduction in the level of both apoptosis and necrosis by inhibition of cellular stress (ionic, osmotic, and/or biochemical) during cryopreservation.40,42,49 We can hypothesize that a decrease in postthaw necrotic and/or apoptotic cell death pathways is related with the improved postthaw recovery found for the two cell lines used in the present study. However, this is the first report on the use of on the use CryoStor™ for the preservation of cell monolayers, and further studies must be performed to understand the cryoprotective mechanisms.
The cryopreservation of differentiated monolayers proved to be more difficult than for undifferentiated cells, due to the development of cell-specific features such as multiple cell layers for Caco-2 or neuronal networks with formation of extensive neuritic processes for N2a cells. Cryopreservation of fully differentiated Caco-2 monolayers resulted in reduced recovery 1 day postthawing when comparing with cells cryopreserved as undifferentiated monolayers. This might be due to additional factors influencing the cryopreservation outcome when freezing such multiple cell layer structures, which are related to the three-dimensional arrangement and limitations in heat and mass transfer. 83 Full recovery of cell metabolic activity could only be obtained after 10 days in culture, which may not be acceptable for the desired applications of the monolayers on clinical and experimental settings. Thus, to achieve the desired cell recoveries, adjusting other cryopreservation parameters, such as freezing and thawing rates,2,53,84 cryoprotectant concentration, or incubation time with the cryoprotectant, 2 may be necessary.
For the cryopreservation of differentiated N2a cells, even though cell damage was reduced when CryoStor™ was used, entrapment beneath a layer of alginate was crucial for maintenance of attachment and integrity of the neuronal network. Ma et al. have reported on 60% cell recovery immediately postthawing for adherent neuronal networks cryopreserved beneath a thin film of collagen gel using culture medium supplemented with high concentrations of bovine serum and DMSO and trehalose as the cryoprotectants. 85 However, no further studies were performed to assess cell recovery within the following h postthawing, which is of major importance for the evaluation of the full extent of postthaw cell death due to the time course of apoptotic and necrotic mechanisms has previously reported.42,49,77 Our results show that combining the use of serum-free CryoStor™ solution and alginate entrapment improves the cryopreservation of neuronal networks by maintaining proper neuronal connections and functionality and allowing 100% recovery of cell metabolic activity within 3 days postthawing.
Conclusions
The combination of different strategies based on cell entrapment in ultra-high viscous alginate and serum-free CryoStor™ solution leads to optimized protocols for the cryopreservation of adherent cells in a fully differentiated state. Such protocols enabled the retention of cell membrane and whole monolayer integrity, metabolic activity, and cell-specific function/differentiated state, a major requirement for clinical and pharmacological applications of such cell lines. Successful cryopreservation of large numbers of viable and functional adherent cells on micro-plates eliminates the time-consuming process of inoculation and expansion from a frozen vial of cells, thus reducing time between cell storage and use in experimental or clinical settings. Further, such protocols as developed here facilitate the creation of cellular clone banks, reducing batch-to-batch variability and normalizing passage age, which are obstacles for achieving reliable results in pharmacological testing. Finally, these protocols should support the implementation of routine cryopreservation practices during preparation of engineered cells and tissues for clinical applications and enable the immediate availability of cells grown on biocompatible matrices for transplantation to treat several injuries (e.g., spinal cord, cartilage, skin, cornea, etc).
Footnotes
Acknowledgments
The research described herein was supported by the European project Cell Programming by Nanoscaled Devices (NMP4-CT-2004-500039) and by the Fundação para a Ciência e Tecnologia (“Cryothera” 010.6/A006/2005). Additional granting was received by the Bundesministerium für Bildung und Forschung (BMBF, Grant no. 03N8707) given to H. Zimmermann. We thank Daniela Ferreira and Norbert Puetz (Center of Electron Microscopy of University of Saarland) for technical assistance. R Malpique acknowledges Fundação para a Ciência e Tecnologia for finantial support (Grant no. SFRH/BD/22647/2005). The authors thank C. Brito for helpful discussions.
Disclosure Statement
No competing financial interests exist.