
Abstract
An intelligent culture system accompanied by automated operations (liquid transfer and cell passage) was newly developed to perform serial cultures of human skeletal muscle myoblasts. To realize a desired performance, a laminin-coated surface was applied to myoblast expansion in a culture flask. It was found that the laminin coating enhanced the overall growth ability attributable not to shortening of the doubling time but to prevention of differentiation toward myotube formation, compared with that on a conventional plain surface. In addition, the effects of seeding density and confluence degree on the growth were investigated quantitatively in terms of cell attachment and division as well as proliferative cell population in the culture on the laminin-coated surface. With increasing in seeding density, the number of proliferative cells decreased at the end of culture accompanied by an increase in the confluence degree, which caused poor attachment of the passaged cells on the surface in the subsequent culture. The quantitative analyses of these cell behaviors helped us determine the appropriate seeding density and attainable confluence degree during one passage, which were 1.0 × 103 cells/cm2 and 0.5 as the initial and boundary conditions, respectively. An automated culture system that could manage two serial cultures by monitoring the confluence degree was constructed. The automated operation with the intelligent determination of the time for passage was successfully performed without serious loss of growth activity, compared with manual operation using conventional flasks. These results indicated that the monitoring of confluence degree is effective to perform the culture passage of myoblasts, being contributable to automating the cell expansion process.
Introduction
The formation of myoblast sheets demands a large number of the patient's cells, followed by in vitro cell expansion. The culture of myoblasts is still challenging, given their low growth ability owing to the differentiation accompanied by myotube formation. 8 Mononuclear cells, which can proliferate, actively fuse with surrounding cells, forming the myotubed multinuclear cells that do not proliferate. Therefore, the apparent growth ability in cultures depends on the balance between mononuclear and multinuclear cell populations. Cell expansion sufficient for myoblast transplantation has been studied by considering biological phenomena for the shortening of doubling time of mononuclear cells as well as hindering their myotube formation, resulting in desired culture performances accompanied by the surface modification of substrates9–11 as well as the administration of soluble growth factors in media.12–15
Laminin is well known to play an important role in attachment, growth, and differentiation of cultured cells. Paulsson 16 reviewed that its biological functions have been used to offer preferable conditions for cell expansion as well as differentiation of cultured cells. In addition, while many matrix components, such as fibronectin and collagen, are rather ubiquitous in distribution of the developing embryos, laminin is localized in the basement membranes between epithelial cells and others, suggesting that laminin may make a specific contribution to the guidance of cell migration and the induction of mesenchyme by epithelial layers during morphogenesis. Goodman et al. 17 reported that in vitro myoblast growth was promoted on a laminin-coated surface because of its action as a myoblast mitogen to induce higher locomotion, leading to suppressed differentiation into myotubed cells. These biological functions of laminin can be helpful for the improvement in cell expansion based on sophisticated process managements, including optimization and automation for culture operations.
In practical processing for cell expansion, the cell sources harvested from the patients are limited in quantity. Haider et al. 6 pointed out that cell processing with serial passages is required to obtain the sufficient number of 106 to 109 cells for transplantation. From an operational aspect, however, the manually performed culture of autologous cells is often associated with unstable outcomes owing to fluctuating in culture conditions. An automated culture system can be a possible choice to overcome these requirement and problem. In our previous study, a mechanically automated system was developed to perform serial cultures for the expansion of human mammary epithelial cells, 18 and the culture system was incorporated with the operations of medium change and passage, which were scheduled in advance with a prediction tool using a kinetic growth model.
To meet medical demands, the current study aimed at attaining the total number of myoblasts and cell expansion to seeding density of over 1.5 × 107 cells and 200 times, respectively, in a serial culture run. In addition, a larger-scale culture system was newly designed for the expansion of myoblasts. To achieve a higher population of proliferative cells, the boundary condition of the confluence degree at the end of the myoblast culture and the initial condition of seeding density were determined by considering the behaviors of the myoblasts. An intelligent culture system that can determine the time for passage autonomously was established to perform serial cultures of myoblasts.
Materials and Methods
Cells and culture conditions
Cryopreserved human skeletal muscle myoblasts (Lot no. 4F1618; Lonza Walkersville, Walkersville, MD) were thawed according to the supplier's instruction. The subculture of the myoblasts was conducted in a 75-cm2 T-flask (Nunclon Delta Flask; Nalgen Nunc International, Rochester, NY) using Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO) containing 10% fetal bovine serum (FBS; Invitrogen, Grand Island, NY) and antibiotics (100 U/cm3 penicillin G, 0.1 mg/cm3 streptomycin, and 0.25 mg/cm3 amphotericin B; each obtained from Gibco, Invitrogen, Grand Island, NY). The culture was maintained in an incubator controlled at 37°C under an atmosphere of 5% CO2. The medium was maintained at a depth of 2 mm in the T-flask, and the spent medium was replaced daily. After reaching 70% confluence on the culture surface, the cells were detached by enzymatic treatment with a 0.1% trypsin/0.02% EDTA solution (Sigma-Aldrich), and the trypsin was inactivated using a 0.1% trypsin inhibitor solution (Wako Pure Chemical Industries, Osaka, Japan). After the removal of trypsin and trypsin inhibitor by centrifugation, the collected cells were resuspended in fresh medium, followed by the inoculation of the cells for the subsequent subculture. Depending on necessity, the cultures were conducted using an eight-square-well plate (Nunclon Delta Multidish; Nalgen Nunc International). The seeding densities were in the range of X0 = 1.0 × 102 to 1.0 × 104 viable cells/cm2, as determined by the trypan blue exclusion test on a hemocytometer under an optical microscope. The cells that have been passed less than five times were used in the experiments.
A laminin-coated surface was prepared according to the supplier's instruction under a sterile condition just before starting the culture. A 50 μg/mL laminin solution (laminin-1; Sigma-Aldrich) in phosphate-buffered saline (PBS; Sigma-Aldrich) was introduced into the culture vessel and incubated for 1 h at 37°C, followed by washing with a large amount of PBS.
Determination of adherent cell number, efficiency of cell attachment, specific growth rate, growth achievement and potential, and confluence degree
It is well known that the fusion between myoblasts occurs to yield the myotubed cells showing a multinuclear form, leading to the difference between cellular and nuclear numbers during culture. To express overall culture profile or growth potential, the parameter estimation on the basis of cells or nuclei, respectively, was conducted.
With respect to growth profile, adherent cell number, efficiency of cell attachment, and specific growth rate were estimated by visual cell counting on captured images. To determine the adherent cell concentration (Xa), the images of culture surface were captured from more than four different positions in each vessel every 24 h using a digital camera (CS6931; Toshiba Teli Corporation, Tokyo, Japan) attached to an optical microscope (area of captured image: 0.023 cm2). The number of cells was counted manually on the images obtained at a given culture time (t). The efficiency of cell attachment (α) was defined as the ratio of adherent cell concentration at t = 24 h to seeding density, and apparent specific growth rate (μapp) based on the total adherent cell concentration was evaluated by the following equation.
where Xa1 and Xa2 are adherent cell concentration at t = t1 and t1 + Δt, respectively, and Δt is differential time (Δt = 24 h in this study).
To estimate growth potential, the number of proliferative nuclei was estimated as follows. The myoblasts were stained with 5-bromo-2-deoxyuridine (BrdU; Roche Applied Science, Penzberg, Germany) and 4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes, Eugene, OR). Before staining, the cells were incubated with 10 μM BrdU in DMEM for 12 h according to the supplier's instruction. The fixation of cells and denaturation of the DNA double strand were carried out by adding FixDenat (Roche Applied Science) followed by three washings with PBS. After masking the nonspecific binding sites for 30 min at ambient temperature with 10% FBS for 30 min, the cells were kept for 2 h at 37°C with a mouse monoclonal anti-BrdU antibody (1:250 dilution; Sigma-Aldrich). The cells were then immunolabeled with goat anti-mouse IgG (1:400 dilution, Alexa Fluor 568; Molecular Probes) accompanied by nuclear staining using DAPI (1:15,000 dilution). Images were captured from each sample using a fluorescence microscope, and the populations of BrdU-positive and DAPI-positive nuclei as proliferative and total nucleus numbers, respectively, were evaluated.
The growth achievement (Ec) was defined as the expansion index on a nucleus basis, which was determined from the ratio of nucleus number at a given culture time to seeding density. The growth potential (Rp) was evaluated from the ratio of proliferative (BrdU-positive) nucleus number to total (DAPI-positive) one.
The confluence degree (Cd) was defined as the ratio of the area occupied by the cells to the entire area of captured image on the culture surface. The Cd was determined using a custom-made tool programmed with commercially available software (LabVIEW, National Instruments, Austin, TX) as described elsewhere. 19
F-Actin staining
For the F-actin staining, the cells were fixed for 10 min at room temperature with 4% paraformaldehyde in PBS, followed by permeabilization with 0.1% Triton X-100 in PBS for 5 min. The cells were then treated with rhodamine-conjugated Alexa Fluor 568 (Molecular Probes) for F-actin staining at room temperature.
Gene expression analysis
To detect the mRNA expression of myogenin, which is a differentiation marker for myotube formation, total RNA was extracted from the cells using an RNeasy mini kit (Qiagen, Hilden, Germany), and the RNA sample was subjected to DNase-I (Qiagen) treatment according to the supplier's instruction. In brief, the reverse transcription from total RNA (10 μL of sample with a total of 0.13 μg RNA) was carried out using an ExScript™ RT reagent kit (Takara Bio, Shiga, Japan) initially for 15 min at 42°C and afterward for 5 min at 95°C. An aliquot of obtained cDNA sample (2 μL) was used to examine gene expression. Gene expressions were examined for the cDNA (0.026 μg) by means of quantitative real-time PCR with a Chromo4™ detector and furnished program (Bio-Rad Laboratories, Hercules, CA). Specific primers were designed using the Primer3 software (available at
Analysis
All the data were recorded as an average of triplicate determinations. The statistical analyses were performed using unpaired t-test. The performance using the automated culture system was conducted twice under each experimental condition, and the reproducibility of the data was confirmed.
Results
Culture of myoblasts on laminin-coated surface
The cultures of human skeletal muscle myoblasts were performed on plain and laminin-coated surfaces at a seeding density (X0) of 1.0 × 103 cells/cm2. As shown in Table 1, the myoblasts on the laminin-coated surface showed a higher growth ability, and the cell density reached Xa = 1.6 ×104 cells/cm2 at t = 144 h, which was 1.5-fold higher than that of the cells grown on the plain surface. In addition, the numbers of proliferative (BrdU-positive) nuclei decreased in both cultures on plain and laminin-coated surfaces in a time-dependent manner, and the growth potential estimated at t = 144 h was Rp = 0.58, which was 1.3-fold higher than that of the cells grown on the plain surface. To evaluate a differentiation state, the mRNA expression of myogenin (one of markers for myotube formation) was analyzed in the cultures of myoblasts on the plain and laminin-coated surfaces. The expression levels for myotube formation were found to increase with elapsed culture time and downregulated expression of myogenin on the laminin-coated surface occurred in comparison with the expression on the plain surface, which showed a reverse trend in the levels of Rp. These results suggested that the differentiation toward myotube formation took place at a higher frequency on the plain surface, compared with that on the laminin-coated surface.
Numbers within parentheses indicate standard deviations (n = 3).
The data on the laminin-coated surface were significantly higher than those on the plain surface (p < 0.01).
The data on the laminin-coated surface was significantly lower than those on the plain surface (p < 0.05).
To estimate the extent of growth enhancement on the laminin-coated surface, the apparent specific growth rate (μapp) based on the total cell concentration was determined. In addition, the proliferative and nonproliferative cells are expected to exist due to the differentiation toward myotube formation during the cultures. Then, the μapp value was considered to depend on the concentration of proliferative cells (Xap) and to be correlated to true specific growth rate of proliferative cells (μ*) as follows.
Hence, the value of Rp was assumed to be constant during differential time (Δt) in Eq. (1), thereby Xap = Xa × Rp.
As shown in Table 1, the μapp values were obtained at t = 48, 72, and 144 h. The estimated μ* values were kept at almost a constant level (μ* = 0.033/h on average) regardless of surface and culture time, meaning that the myoblast growth depended only on the ratio of proliferative cell number. It was thus considered that the apparent specific growth rate mainly relied on the number of proliferative cells on the laminin-coated surface, concluding that the observed enhancement of growth on the laminin-coated surface can be attributed from the suppression of myotube formation.
For the further confirmation of the myotube formation, the F-actin staining and nuclear staining were conducted. As shown in Figure 1, mononuclear cells were found to prevail at the early culture phase (t = 48 h) on both surfaces. Myotubes aligned with multinuclear cells on the plain surface at the late growth phase (t = 144 h), while less myotube formation was observed on the laminin-coated surface. These results indicate that laminin is one of appropriate substrates that allow human myoblasts to propagate with suppressing myotube formation. In addition, it can be stated that Rp is a possible indicator for evaluating an expanding potency of cultured myoblasts in such a way that the Rp shows the inverse trend against the frequency of myotube formation.

Fluorescent images of F-actin (red) and nuclei (blue) of myoblasts observed at t = 48 h (
Effect of confluence in a prior culture on subsequent growth after passage
To investigate the effect of confluence, serial cultures were performed. The first cultures of myoblasts on the laminin-coated surface were conducted for t = 168 h at various seeding densities, and the cells harvested from the first cultures were seeded to conduct the second cultures for estimating the subsequent growth properties in terms of cell attachment and division at t = 48 h. In the first culture, as shown in Figure 2, the growth potential at the end of culture (t = 168 h) decreased with increasing X0, being Rp = 0.13 at X0 = 7.5 × 103 cells/cm2, which was considered to be attributed to the differentiation toward myotube formation as well as contact inhibition with a decrease in the number of proliferative cells. In addition, with increasing X0, the Cd at the end of culture increased, while the growth achievement defined as the expansion index on a nucleus basis at t = 168 h was kept almost constant (Ec > 21) up to X0 = 1.4 × 103cells/cm2, giving Cd < 0.5. A further increment in X0 caused a decrease in Ec, being Ec = 6.7 in the culture at X0 = 7.5 ×103 cells/cm2, which was one-third of the Ec value at X0 =1.0 × 103 cells/cm2.
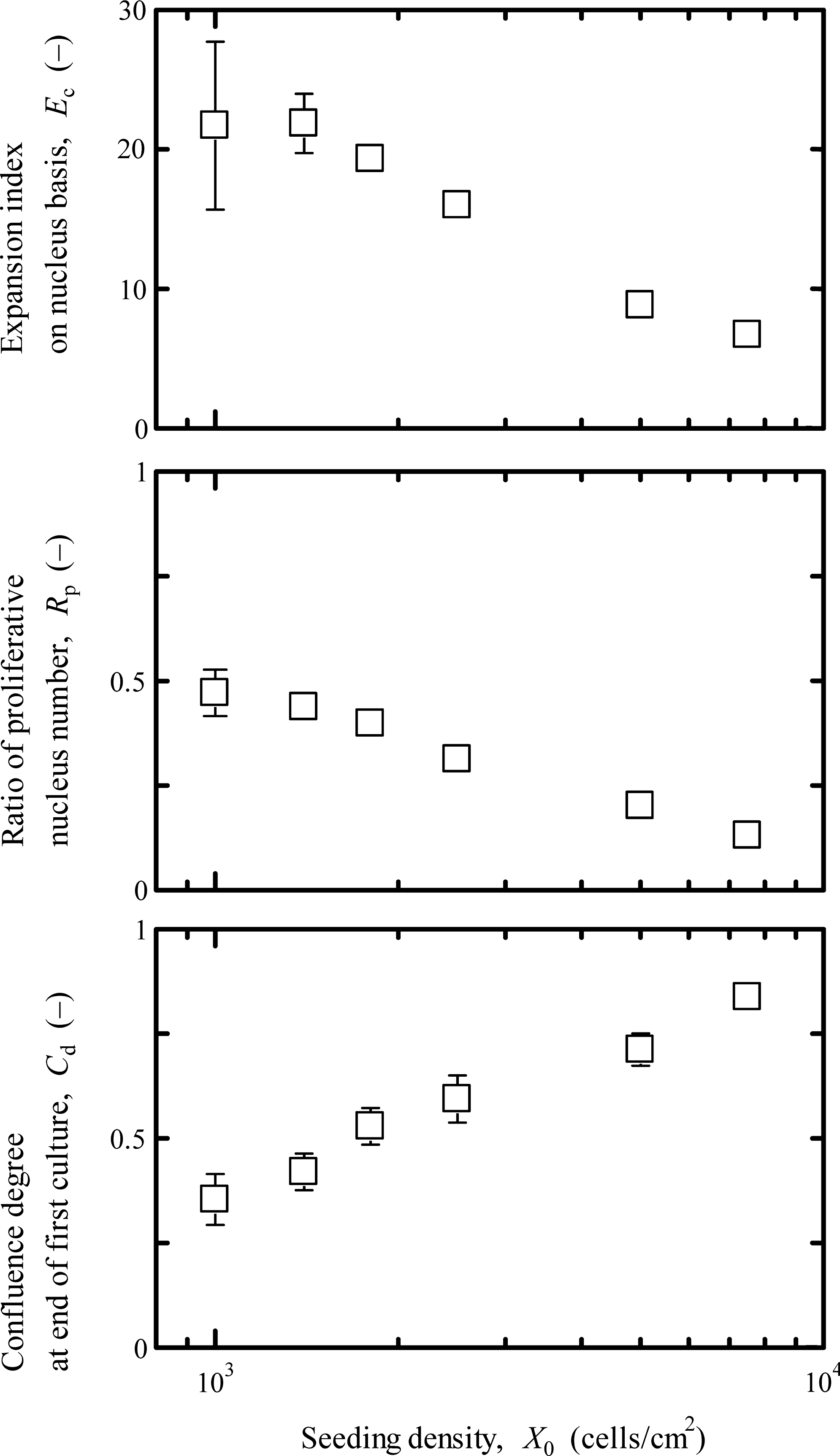
Growth achievement and potential of myoblasts on laminin-coated surface at the end of the first culture (t = 168 h) with various X0 values. The cultures were conducted on the laminin-coated surface. The error bars indicate standard deviations (n = 3).
To understand the effect of culture confluence on the subsequent culture profiles, the cell behaviors of attachment and division in the second cultures were evaluated using cells having different Cd values at the end of the first cultures. As seen in Figure 3, the efficiency of cell attachment in the second culture was constant at α = 0.84 up to Cd = 0.42, and with an increase in Cd over 0.5, the α decreased, achieving α = 0.54 at Cd = 0.84, which was approximately 36% less than that at Cd = 0.35. However, the growth potential at t = 48 h in the second culture was kept constant at Rp = 0.76 on average, which was comparable to that at the onset of the first culture. By arrangement of the data in Figures 2 and 3, with increasing Rp at the end of the first culture (t = 168 h), the Rp at the beginning of the second culture (at t = 48 h) increased. This suggested that the myotubed cells in the population at the end of the first culture failed the attachment after passage, and in the subsequent culture the proliferative cells become enriched, resulting in the higher Rp at t = 48 h. In the practical performance of serial cultures, the appropriate Cd at the end of a foregone culture was found to be less than 0.5 so as to realize the desired cell attachment in a subsequent culture.
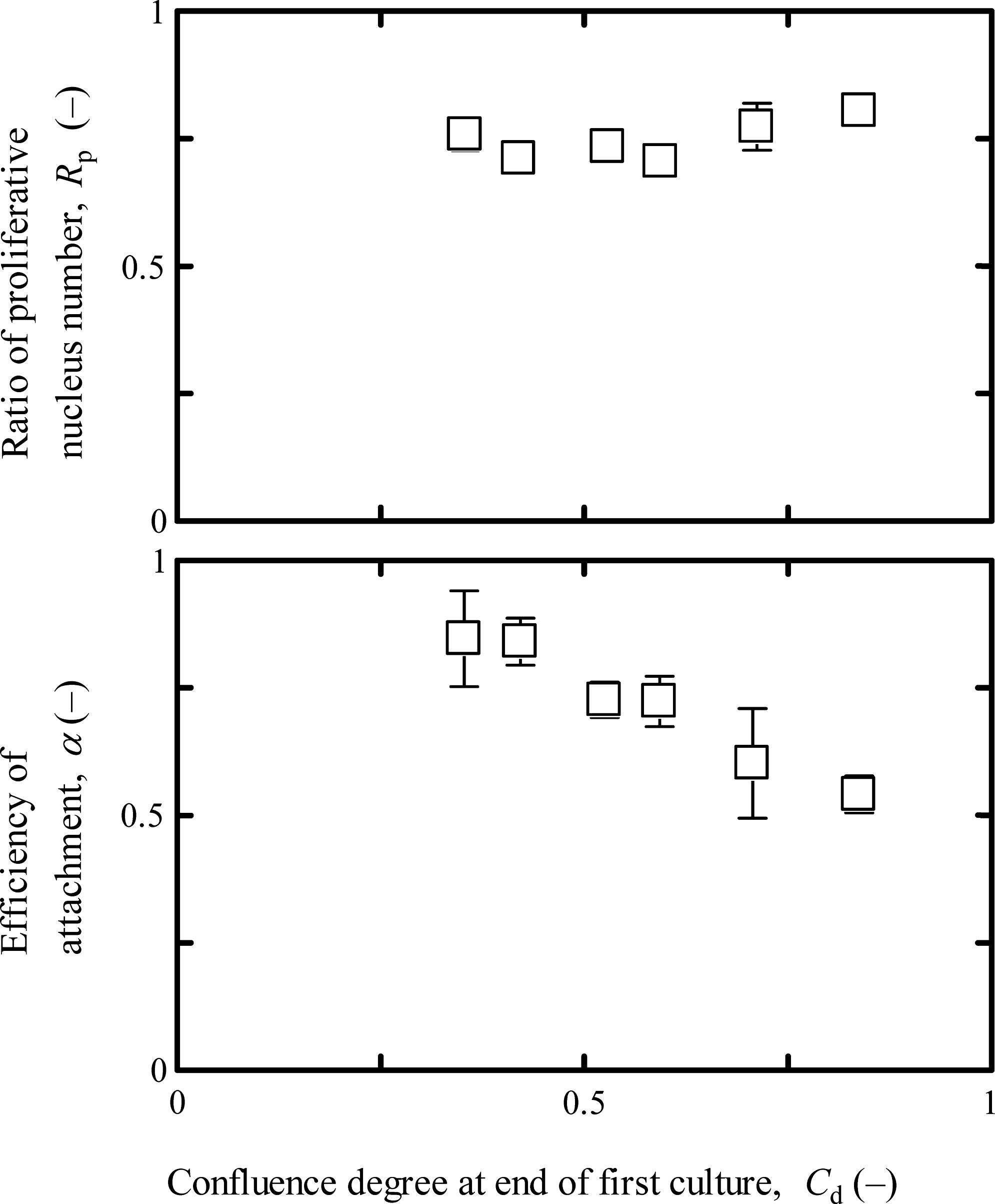
Effect of confluence degree at the end of the first culture on attachment and growth potential in the second culture. The cultures were conducted on the laminin-coated surface, and the α and Rp values were determined at t = 24 and 48 h, respectively, in the second culture. The error bars indicate standard deviations (n = 3).
Effects of seeding and passage on growth potential
To evaluate the growth potential, the cultures seeded at X0 = 1.0 × 102 to 1.0 × 104 cells/cm2 were conducted on the laminin-coated surface until the same confluence of Cd = 0.5 was achieved. As shown in Figure 4, with increasing X0 up to 1.0 × 103 cells/cm2, the Rp increased linearly and then reached a plateau. The Rp at X0 = 1.0 × 103 cells/cm2 was estimated to be 0.36, which was twofold that at X0 = 1.0 × 102 cells/cm2, meaning that the growth potential at the end of the culture was enhanced with increasing seeding density, and the appropriate seeding density for cell expansion was over X0 = 1.0 × 103 cells/cm2.

Effect of seeding density on growth potential at Cd = 0.5 in the first culture. The cultures were conducted on the laminin-coated surface. The error bars indicate standard deviations (n = 3).
To ensure the synergy of seeding and passage, the cultures for t = 288 h were carried out under various operational conditions as shown in Figure 5. In the performance at X0 = 1.0 × 103 cells/cm2 with passage operation (run 1), the growth achievement reached Ec = 248 at t = 288 h. Moreover, the growth achievement in the performance at X0 =1.0 × 102 cells/cm2 without passage operation (run 2) was slightly higher, but not significantly different from that in run 1, whereas the growth potential at t = 288 h in run 1 was estimated to be Rp = 0.49, which was threefold that in run 2. In the performance at X0 = 1.0 × 103 cells/cm2 without passage operation (run 3), the confluence reached a higher level of Cd = 0.72 at t = 288 h, resulting in lower growth achievement and potential, compared with those in run 1. From these results, it was concluded that the passage is inevitable to enable a successful culture performance acquiring preferable growth achievement accompanied by high population of proliferative cells.

Growth achievement and potential of myoblasts in various culture performances. The cultures were conducted on the laminin-coated surface, and the Cd, Ec, and Rp values were determined at the end of each culture (total t = 288 h). Passage operation was carried out at Cd = 0.41 in run 1 at the end of the first culture. #The data in run 1 was significantly higher than those in runs 2 and 3 (p < 0.0005). ##The data in run 3 was significantly lower than those in runs 1 and 2 (p < 0.005). Numbers within parentheses indicate standard deviations (n = 3).
Performance for expansion of myoblasts in automated culture system
To establish an automated system, the culture process was designed according to the conditions shown in Table 2. As medical demands, in this study, the final number of myoblasts and the growth achievement on a nucleus basis were set to be over nf = 1.5 × 107 cells per each run and over Ec = 200, respectively. As culture constraints, the seeding density and passage were set to be X0 = 1.0 × 103 cells/cm2 and Cd = 0.5 as initial and boundary conditions, respectively, which were determined from the foregoing mentioned results. Under these conditions, then, we plan to proceed the myoblast cultures in a novel culture system fabricated using a 75-cm2 T-flask (Nalgen Nunc International) and a 632-cm2 container (Cell Factory; Nalgen Nunc International). Details of the culture system are described in the appendix. As shown in Figure 6, two culture vessels were connected to each other using a sterile silicon tube for the transport of the cell suspension. The entire operation, including observation, medium change, and passage, was carried out automatically using a computer installed with custom-made program. The cell growth was monitored by the capturing and processing of images. The observation was conducted every 24 h to determine the Cd from more than four images at different positions of the bottom surface. Here, the Xa values were also obtained by the manual counting of cells on the images. Medium change was carried out daily by replacing the spent medium with fresh one while keeping at 2-mm depth in the culture vessel. The passage was designed to eliminate centrifugation, similarly to that used in the previous work. 18 The detachment of cells in the first culture was conducted by the trypsinization after rinsing the culture surface with PBS, and the trypsin inhibitor solution was added to the cell suspension. The resulting cell suspension was transferred into the container using a tube pump. After transporting the cells into the container with the medium, the passage without centrifugation was accomplished to initiate the second culture. At t = 24 h after reseeding in the container, cell attachment was completed, and the medium with trypsin and its inhibitor was replaced with a fresh one. As an alternative control, the manual performance employed in the routine subcultures, accompanied with centrifugation in the passage operation, was also conducted in 75-cm2 T-flasks according to the subculture protocol described in Materials and Methods section.

Schematic drawing and images of the automated culture system. (
Figure 7 shows the performance of serial cultures using the automated system. The first culture was initiated at X0 = 1.0 × 103 cells/cm2. With elapsed time, the Xa increased, and the cells grew exponentially after t = 24 h. When the confluence reached Cd = 0.5 (t = 186 h), passage was carried out to initiate the second culture. At 24 h after reseeding in the second culture (t = 210 h), the exponential growth started. The culture ceased at Cd = 0.5 (t = 330 h), producing the final cell number of nf = 1.8 × 107 cells in the system. The culture achievement using the automated system was totally equivalent to those obtained from both the manual performances with and without centrifugation in the passage operation. Table 3 summarizes the culture performance in the automated system. The final values of growth achievement and potential on a nucleus basis in the automated system were Ec = 201 and Rp = 0.32, respectively, which meets the predetermined requirement to a successful extent. Moreover, the cell attachments in the first and second cultures using the automated system reached α = 0.91 and 0.88, respectively, giving the equivalent values obtained from the cultures conducted manually.

Time profiles of serial cultures of myoblasts in automated manner (square) and manual manner with centrifugation in passage operation (triangle). The cultures were conducted on the laminin-coated surface and proceeded according to the conditions listed in Table 2.
Numbers within parentheses indicate standard deviations (n = 3).
Discussion
For the autologous transplantation of patient's cells, cell expansion is one of the most important steps. Serial cultures accompanied by passage are inevitable because contact inhibition prevents cell division, resulting in slow proliferation at high confluence. In our previous study, 20 the behavioral property of cell division was characterized by the direct observation of individual murine dermal fibroblasts in culture, and the doubling time for division was found to depend on the number of surrounding cells, indicating the importance of confluence at passage as a process parameter in serial cultures for cell expansion. Human skeletal muscle myoblasts examined in the current study also showed contact inhibition at high confluence (data not shown). Therefore, the confluence was evaluated quantitatively by defining the confluence degree, which can be used as an operational parameter for determining the start time for passage.
There have been developments to improve in vitro proliferation of myoblasts, and the major efforts are to encourage the cell division by using the laminin-coated surface. Öcalan et al. 11 reported that cell division on the laminin-coated surface was promoted successfully in the culture of murine myoblasts, resulting from the enhancement of migration. In this study, the laminin-coated surface was applied to the culture of human myoblasts, and it was demonstrated that the growth potential was enhanced, compared with that on the plain surface. In addition, the quantitative analysis of proliferative cells revealed that the growth enhancement was caused by not only the promotion of cell division but also the increase in population of proliferative cells in culture, resulting from the inhibition of myotube formation. Myotube formation is well known as a process toward terminal differentiation, which cooperates with growth arrest, making cell expansion difficult. 9 Therefore, its prevention can alternatively improve the growth of myoblasts. Myotube formation occurs when cells make contact during cell division as well as when they encounter each other. Therefore, the separation of parent and daughter cells was suggested to inhibit the myotube formation. 10 The laminin-coated surface promoted cell migration, and the average migration rate was estimated to be 0.64 μm/min, which was 1.8 times higher than that on the plain surface (data not shown). Goodman et al. 17 also recognized the promoted migration of myoblasts on the laminin-coated surface. The promotion of migration was suggested to assist in the separation after cell division, resulting in the inhibition of myotube formation that affects the population of proliferative cells (Table 1). In addition, higher confluence enhances the frequency of cells encountering each other, considering that the prolongation of the culture period was inadequate to maintain a higher population of proliferative cells.
In the serial cultures, the population of proliferative cells was maintained at a relatively high level (Rp = 0.76 on average) after passage (Fig. 3) despite of the different Rp levels at the end of foregone culture, which was suggested to arise from the lower attachment of myotubed cells after seeding (Fig. 3). In addition, the prolongation of culture without passage (run 2 in Fig. 5) caused a decrease in the Rp although the Ec increased with an increase in nonproliferative cells, compared with those in the culture with passage (run 1 in Fig. 5), clarifying that the operational improvement by passage in serial cultures led to the increased population of proliferative cells at the onset of subsequent culture.
Together with these phenomena described above, the determination of the operational parameters in serial cultures such as seeding density and confluence degree at passage could be critical to the successful expansion of myoblasts under initial and boundary conditions, respectively. In this study, the culture strategies for myoblast cell expansion were established to include initial and boundary conditions of X0 = 1.0 × 103 cells/cm2 and Cd = 0.5, respectively (Table 2), and successful results were achieved in such a way to satisfy the required performance (Fig. 7).
For manufacturing consideration, the culture process for cell expansion aims to obtain the desired quantity and quality of cells as quickly and stably as possible, and the process has been designed by this evaluating of cell productivity with regard to the growth achievement and potential. 21 In addition, the use of autologous cells often faces an operational difficulty owing to the fluctuation of culture conditions in the manual performance of in vitro cultures, leading to the varied quality of final cell products.22,23 In this context, the development of an automated culture system can be a major challenge to enhance culture performance efficacy. In the present study, the growth achievement and potential were proposed in terms of indicators of expansion index and population ratio of proliferative cells, respectively, and the culture process was designed to achieve higher levels of these indicators in serial cultures of myoblasts (Table 3). In addition, cell maintenance and proliferation were always performed manually using conventional culture vessels. The manual performance in the vessels, however, requires individual handlings in cell seeding, medium change and passage, which limits the scale-up of cultivation. 24 Pörtner et al. 25 reviewed the advantages of the automated culture system, which have been expected to reduce the risk of biological contamination and the fluctuation of culture states, contributing to reproducible culture performance owing to the high operational stability of cell processing. In our previous study, 18 we pointed out that the operational complexity in manual handling raised the contamination risk, suggesting that the operational retrenchment in culture procedures under an aseptic conditions is important. Then, the simplified procedure for passage without centrifugation was proposed to contribute to the operational sequencing of serial cultures in a closed system.
In our previous work, 19 the scale-up of the culture system was conducted up to 3000 cm2 of surface area by stacking growth chambers and the laborious tasks accompanying machinery movement were realized smoothly by introducing pneumatics-aided equipment, which will be contributable to a larger culture performance in our current automated system. A robotic arm system is an alternative machinery movement for handling numerous culture flasks during culture operations. Thomas et al.23,26 reported the culture performance of mesenchymal stem cell expansion in the commercially available system of CompacT SelecT (The Automation Partnership, Hertfordshire, United Kingdom), resulting in the process improvements not only by automation of cultures operated in parallel but also by scalability for the production in an ad hoc lab-based system. Moreover, an essential technique to perform the large-scale culture is the monitoring of culture states in a noninvasive manner. In the reported culture system, a digital camera was placed externally at the bottom of the growth chamber to monitor the culture states by measuring the confluence degree. 23 This monitoring technique could be applied to the culture system for the production of skin substitutes. 27 Observation and medium analysis, which can be accomplished without any loss of cells, are suitable for the practical performance of culture owing to the low risk of contamination, and the information acquired by monitoring can be applied to the determination of the time for medium change or passage. The intelligent culture system can be used for the autonomous performance of culture and facilitates the streamlining and automation of numerous labor-intensive procedures. The current culture system was installed with an observation tool to accomplish passage operation, which can be determined autonomously from confluence degree (Fig. 5). In addition, the cell number was also estimated by monitoring the amount of lactate produced in medium after determining the linear regression of concentrations between cells and lactate (data not shown). The intelligent culture system can be applied to cultures of other adherent cells and will lead to the qualitative stability of products in the practical manufacturing of cells available for transplantation. In other words, the culture system will contribute to the process control as well as the quality control.
In conclusion, the intelligent culture system for the expansion of human skeletal muscle myoblasts was developed to operate the culture passage autonomously. The seeding density and confluence degree were determined to be X0 = 1.0 × 103 cells/cm2 and Cd = 0.5 as initial and boundary conditions, respectively, while considering the inhibition of myotube formation. The performance of serial cultures was conducted successfully by monitoring the time-dependent variation in confluence degree, and the expansion index on a nucleus basis and populational ratio of proliferative cells at the end of culture reached Ec = 201 and Rp = 0.32, respectively, which were equivalent to those obtained in the manual culture performances. The intelligent culture system accompanied with autonomous passage operation was considered to enhance stabilities not only of culture performance but also of cell populational quality.
Footnotes
Acknowledgments
We are grateful to Drs. S. Miyagawa and Y. Imanishi in Osaka University for their invaluable advices. We thank Nova Biomedical Co. for technical supports with respect to monitoring culture profiles using the BioProfile medium analyzer. This study was conducted as a part of the program “Center for integrated cell and tissue regulation” for Centers of Excellent (21st COE), and was supported in part by the New Energy and Industrial Technology Development Organization (NEDO) of Japan and a Grant-in-Aid for Scientific Research on Priority Areas in the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.
Disclosure Statement
No competing financial interests exist.
Appendix
Figure 6 shows the schematic illustration of a culture system, which consists of three parts: a culture module, liquid handling, and control units. The culture module works as an incubator where the growth chambers can be mounted on the stage.
During the cultures, the chambers were maintained at 37°C, and the gas was blown using sterile air with 5% CO2 through a mass flow controller. Medium evaporation could be neglected while keeping the medium depth at 2 mm throughout the culture periods by introducing humidified air. In the liquid handling unit, the liquid transfer during medium replacement and culture passage was achieved using electrical pinch valves and tubing pumps. While the chamber stage was tilted, the spent medium was pumped out of the chamber into a waste medium reservoir. Fresh medium and solutions of trypsin and trypsin inhibitor were preserved at 4°C, and warmed to 37°C before being introduced into the chamber. For cell observation, an 8-bit CCD camera (CSB4000CL-10; Toshiba Teli Corporation) with an objective lens and a lamp was attached to an electrically driven stage placed below the chamber. The observation area was 0.61 mm2 of the culture surface per image with 496 × 500 pixels. The camera can be moved horizontally at an arbitrary location and vertically to adjust the image focus at a preset position. The control unit employed a personal computer with boards for image capturing and RS-232C, and every operation was performed automatically by programming on LabVIEW software (National Instruments).
The culture system can employ a simplified passage procedure without centrifugation, in contrast to the manual handling using conventional flasks. The growth chambers of the T-flask and the container have two culture surfaces, which are 75 and 632 cm2 for the first and second cultures, respectively. When the confluence in the first culture reaches Cd = 0.5, the passage operation starts. On trypsinization of the cells, the medium in the growth chamber is discarded, and the culture surface is washed twice by rinsing with PBS buffer, followed by adding the trypsin solution. After cell detachment from the bottom surface within 5 min, the trypsin inhibitor solution is added to stop the enzymatic digestion. The cell suspension prepared by the further addition of an adequate volume of fresh medium is transferred into the container by tilting the growth chamber. The passage operation is performed within 20 min, and the second culture can be started on the larger surface. In the second culture, the cells can attach, but cell division does not begin owing to the involvement of trypsin and trypsin inhibitor in the medium. 28 Therefore, after cell attachment for 24 h, the medium containing trypsin and trypsin inhibitor is replaced with a fresh one to initiate cell division. 18
During the automated culture, the adherent cell concentration at a given culture time is evaluated by counting manually the cell number on the images captured at several positions. Medium change can be performed by a series of procedures, by tilting the growth chamber, removing the spent medium, filling with fresh one, and returning the chamber to an original position.